
短轮伐毛白杨人工林耗水规律及作物系数曲线构建*
2023-11-05 12:59李玲雅刘金强赵小宁邹松言付海曼席本野
林业科学 2023年10期
李玲雅 邸 楠 刘金强 赵小宁 邹松言 付海曼 席本野
(1. 林木资源高效生产全国重点实验室 北京 100083;2. 干旱半干旱地区森林培育和生态系统研究国家林业和草原局重点实验室北京 100083;3. 北京林业大学省部共建森林培育与保护教育部重点实验室 北京 100083;4. 内蒙古大学生态与环境学院呼和浩特 010021;5. 重庆市林业投资开发有限责任公司 重庆 401147;6. 国家林业和草原局产业发展规划院 北京 100010)
杨树(Populusspp.)因其生长快、适应性强而成为我国发展速生丰产用材林的主要树种。第九次全国森林资源清查数据显示,我国杨树人工林面积已达757 万hm2,占全国人工林总面积的13.25%,是世界上杨树人工林面积最大的国家。人工林产量与质量的提高很大程度上取决于高效集约栽培技术的应用(Xiet al., 2021)。水分是影响杨树生长的最重要因素(董文怡等, 2011; Xiet al., 2016, 2021),可通过水分管理的手段来提高杨树人工林的生产力(Gochiset al, 2000;李广德, 2010; 任忠秀等, 2011; 孙兆地等, 2012; 席本野等, 2012),但人工林水分管理策略的合理制定和高效实施,需依赖对林木耗水规律的准确把握和对林分耗水量的准确估算。
林分耗水量系指林木蒸腾、土壤蒸发消耗的水分以及植株体内所含水分的总和。由于植株体内所含水分占比非常小且影响因素较复杂而常忽略不计,因此,林分的耗水量即林分蒸散量(林木蒸腾+土壤蒸发)。目前,国际上较为通用的植物耗水量计算方法是通过参考作物潜在蒸散量(reference crop potential evapotranspiration,ET0)与作物系数Kc(crop coefficient)和基础作物系数Kcb(basal crop coefficient)计算的作物系数法(Allenet al., 1998)。该方法包括单作物系数法及双作物系数法,其中,单作物系数法以作物系数Kc表示植物耗水量与参考作物耗水量之间的差异,Kc综合反映了植物的物种特异性和土壤蒸发的平均效应。双作物系数法可以区分植物蒸腾和土壤蒸发对植物耗水量的影响,将Kc分为基础作物系数Kcb和土壤蒸发系数Ke两部分,其中,Kcb代表植物蒸腾部分,是表层土壤干燥且根区土壤含水量不构成水分胁迫时植物蒸散量与ET0的比值。在植物完全覆盖地面时,2种作物系数法计算结果差别不大,而地面被植物部分覆盖的情况下,双作物系数法计算得到的植物耗水量比单作物系数法更接近实测值(樊引琴等, 2002)。因此,精确获取作物系数,探明植物耗水变化规律,是实现区域水资源高效利用和制定节水灌溉制度的前提。
研究表明,杨树比大田作物及其他阔叶树种具有更高的耗水需求(王彦辉等, 2006;Petzoldet al., 2011),因此,杨树林地的高效水分管理更加需要获取精准的林分耗水规律和耗水量信息,否则难以在林分最需水的时期给予最适量的水分供给。目前,已有不少杨树耗水特性研究集中于苗木上(方晓娟等, 2010; 董文怡,2011),且近十几年来,关于杨树人工林的耗水研究也大量开展。例如,李广德(2010)和莫康乐等(2014)分别研究了4~5 年生和13 年生杨树人工林的蒸腾特征,但未关注林地土壤蒸发和蒸散的变化规律;Gochis 等(2000)观测了1~3 年生杂交杨人工林的蒸散变化,并构建了用于灌溉管理的Kc曲线,但却未揭示林分蒸腾与土壤蒸发的变化特征;Di 等(2019)系统分析了6~7 年生杨树人工林的林木蒸腾和土壤蒸发变化,并构建土壤蒸发和Kcb预测模型,但未对林分蒸散量进一步分析。由此可见,目前关于杨树人工林耗水规律的研究大多关注林分耗水的单一组分,鲜有综合分析耗水多组分变化特征;同时,现有研究多为在较短的时间尺度上,鲜有对整个轮伐期内的杨树人工林耗水规律开展深入分析,更缺乏将林分耗水信息转化为作物系数信息并将其应用于林分的水分管理。
综上,本研究针对我国北方重要乡土树种、同时也是华北地区木材储备林建设的重要树种—毛白杨(Populus tomentosa),以其短轮伐人工林为研究对象,系统研究其轮伐期尺度上的耗水规律。本研究的具体目标为:1) 分析毛白杨人工林蒸腾、土壤蒸发及蒸散量的动态变化规律;2) 探明毛白杨的季节生长动态及其作物系数和基础作物系数的变化特征,并依此构建毛白杨的(基础)作物系数曲线;3) 揭示林地Kc、Kcb与叶面积指数及地下水位间的关系,构建Kc和Kcb预测模型。本研究结果将有助于准确估算短轮伐毛白杨人工林的耗水量,从而为其林地高效水分管理的实现提供关键理论依据与数据支撑。
1 研究区概况
研究区位于山东省高唐县国有旧城林场(36°48′11′′—36°48′12′′N,116°4′57′′—116°4′58′′E),海拔30 m,年日照时数 2 651.9 h,年均气温 12. 0~14. 1 ℃,全年无霜期 204 天。试验地中,0~140 cm 土层为砂壤土,140~300 cm 土层为粉壤土,平均土壤密度为1.43 g·cm-3,田间最大持水量为 0.35 cm3·cm-3,饱和含水率为 0.44 cm3·cm-3。
三倍体毛白杨无性系‘B301’((P. tomentosa×P.bolleana)×P. tomentosa),简称毛白杨,于2015 年春季栽植,林分株行距2 m×3 m。2016—2018 年及2020 年每株每年穴施纯氮82 g,2019 年每株穴施纯氮113 g,采用除草剂定期去除杂草。林分内共设置 5 个水分处理,具体信息详见 Liu 等 (2022)和 Jiao 等(2021)。本研究滴灌系统采用一行两带的方式布设(滴管位于树木两侧,距树30 cm 处,滴头间距50 cm,流量1.6 L·h-1),于2016 年4 月启动使用,滴灌充分,在无土壤水分亏缺的条件下研究林分的潜在耗水规律根据以及毛白杨生长与土壤水分有效性间定量关系(Xiet al.,2016)。2016—2017 年当滴头下方 20 cm 处土壤水势达到 -20 kPa 时进行滴灌,使土壤湿润体内水分达到田间最大持水量。随着林分生长,为满足林木增大的水分需求,2018—2020 年当土壤水势达到 -18 kPa 时进行滴灌。根据试验期间获取的生长数据可知,林分6 年生时(2020 年)的蓄积连年生长量与年均生长量变化曲线交叉,因此,本研究中毛白杨林分轮伐期为6 年。
2 研究方法
2.1 林分蒸腾
利用热扩散探针(北京时域通科技有限公司,北京,中国)于树干北侧1.3~1.5 m 高度处插入测定液流速率(Js, cm·s-1)。探针安装完毕后,使用橡皮泥对探针基部与树干连接处进行黏固,然后采用泡沫和锡箔纸分别进行探针的固定和包裹,具体安装过程参照Yang 等(2019)。2016 年使用10 mm 长度探针测定液流速率,2017—2020 年使用30 mm 长度探针。2016—2017 年生长数据显示,林木胸径变异很小,选择平均标准木进行插针测定;2018—2020 年,林木分化导致林木胸径差异增大,根据胸径分布规律,将林木划分为不同径阶,并在每一径阶选择1 株平均标准木进行插针测定。由于探针数量有限,2016—2020 年测定样树株数分别为5、5、5、5 和6 株。根据赵小宁等(2023)对毛白杨不同液流速率计算公式的对比研究结论,本研究采用Granier 原始公式(1)(Granier, 1987)对热扩散探针测定的温差数据进行计算以得到液流速率。
式中:温差系数K=(ΔTM-ΔT)/ΔT;ΔTM为液流速率为0 cm·s-1时的温差 (℃);ΔT为某一时刻的瞬时温差(℃)。林分蒸腾量 (Tr, mm·d-1)由单木蒸腾量推导而来,2016—2017 年Tr 计算见公式(2),2018—2020 年Tr 计算见公式(3)。
式中:Js-ave为测定的平均标准木的液流速率平均值(cm·s-1);Js-i为第i级径阶平均标准木的液流速率(cm·s-1);As-total为测定林地所有林木的边材面积总和(cm2);As-i为第i级径阶所有林木的边材面积总和(cm2);AG为测定林地的面积 (m2)。
林木边材面积利用前期建立的异速方程(Zhaoet al., 2023),以液流测定位置处的树干直径为自变量计算得到。
2.2 土壤蒸发
受冠层发育的影响,林地内距树不同距离的土壤会接受到不同程度的降雨量和太阳辐射,从而导致林地不同位置的土壤蒸发存在差异。为将土壤蒸发的空间变异考虑在内,2016 年于林内选取2 株标准木作为样树,在距离每株样树树干30、80 和130 cm 处各安装1 个内径7.5 cm、高度20 cm 的微型蒸渗仪(Boastet al., 1982)进行土壤蒸发测定。2016 年的土壤蒸发数据表明,距树80 和130 cm 的土壤蒸发无显著差异,此外,前期研究发现灌溉下的最大土壤水平湿润距离约为50 cm(李豆豆等,2018)。因此,2017—2020 年只在距树30 和80 cm 处测定土壤蒸发。试验期间,每3~4 天灌溉或降雨后对蒸渗仪进行换土,换土后每天使用精度为0.1 g 的电子天平对微型蒸渗仪进行称重,然后采用质量减差法(Xiet al., 2017)计算林内每日的土壤蒸发量(Es,mm·d-1)。
2.3 林分蒸散量及参考作物潜在蒸散量
林分蒸散量(ET,mm·d-1)即林木蒸腾和土壤蒸发的总和,采用公式(4)进行计算。
2016 年,使用位于林地中心的自动气象站(Delta-T Devices Ltd., Cambridge, UK)对气象数据进行监测。2017 年起,采用林内和距离林地900 m 的自动气象站(北京时域通科技有限公司,北京,中国)对林内和林外的气象数据进行同时监测。测定的气象指标包括太阳总辐射(W·m-2)、空气温度(℃)、空气相对湿度(%)、风速(m·s-1)、风向和降雨(mm)。由于2016 年(2年生)林木高度较低,且2016 年和2017 年同时期林内和林外气象站采集的气象数据没有明显差异。因此,根据Allenet al. (1998)的计算方法,利用林内气象站监测的气象数据计算2 年生林分的ET0(mm·d-1),利用林外气象站监测的气象数据进行计算3~6 年生林分的ET0。
式中:Rn为净辐射量 (MJ·m-2d-1);G为土壤热通量(MJ·m-2d-1);T为平均气温 (℃);u2为2 m 高处的平均风速 (m·s-1);es为饱和水汽压 (kPa);ea为实际水汽压(kPa);Δ 为饱和水汽压与温度曲线的斜率;γ 为干湿表常数。
2.4 作物系数及基础作物系数
采用FAO-56 (Allenet al., 1998)中双作物系数法对作物系数Kc和基础作物系数Kcb进行计算。
式中,Ks为水分胁迫系数,反映土壤含水率不足时对植物蒸腾的影响,本研究中毛白杨在生长过程中水分供应充足,故Ks=1。Ke为土壤表面蒸发的蒸发系数,Ke= Es/ ET0。
由上推导可得,Kc为林分实际蒸散量与ET0的比值,Kcb为林分蒸腾量与ET0的比值。
2.5 叶面积指数、地下水位及茎干直径
采用Li-Cor 2 200 冠层分析仪(PCA, Li-Cor Inc.,Lincoln, Nebraska, USA)测定林分叶面积指数(LAI,m2·m-2)( Liuet al.,2022)。基于实测的LAI 值,采用线性内插法估算林分生长季内每日的LAI。
利用位于林地中心的自动观测井和水位计(G3061, 北京时域通科技有限公司)测定林分地下水位(GWL,m)。此外,在林内选择3 株标准木,于树干约 1m 高度处安装直径微变化传感器(北京时域通科技有限公司)对树干直径的生长动态进行测定,测定频率为 每10 min 一次,精度为1 µm。上述采集的数据均存储于自动采集器(i-logger,北京时域通科技有限公司)。
2.6 数据分析
本研究中的所有数据分析均在半月时间尺度上进行,为此,从4 月1 日开始,将每15 天的数据做平均(10 月13 日—10 月31 日期间的数据为19 天数据的平均值)后用于分析。此外,因降雨天未能获取土壤蒸发实测数据,且降雨会导致蒸腾大幅度下降,为得到环境稳态期间的Kc和Kcb,故剔除其降雨天数据。
热扩散液流探针采集的温差数据采用Baseliner软件进行处理和计算(Oishiet al., 2016)。使用SPSS(v. 19.0, SPSS Inc., Chicago IL, USA)对不同时期的平均茎干直径日增长率(Trunk diameter daily growth rate,DGR; µm·d-1)数据进行配对样本t检验,以确定其与0 µm·d-1是否在P= 0.05 水平上存在显著差异,如差异显著,则认定该时期为林木主要生长阶段(Deslaurieret al.,2003)。使用Origin 10.0 (OriginLab Corporation, MA, USA)对不同变量间的关系进行线性和非线性回归分析以及绘图。
3 结果与分析
3.1 林分生长动态
由图1 可知,毛白杨每年的主要生长阶段有所变化,但4 月中旬—7 月末为所有林龄林分的主要生长阶段。此外,5—7 月为毛白杨生长速度最快的时期,2~6 年生林分在该时期的生长量占年总生长量的69%~88%。

图1 2~6 年生毛白杨人工林茎干直径日增长率(DGR)的动态变化Fig. 1 Dynamic changes of trunk diameter daily growth rate (DGR) in 2-6 years old Populus tomentosa plantation
根据轮伐期内林分的平均 DGR 和 LAI 季节动态,可将毛白杨生长季划分为 4 个阶段(图2)。第I 阶段为生长前期,时间为 4 月初— 4 月中旬,该时期内林木刚启动展叶,生长速率较小。第 II 阶段为发展期,时间约为 4 月中旬— 6 月中旬,该阶段林木冠层发育迅速,LAI 快速升高,林木生长速度较快,并出现全年的第1 个生长高峰(5 月初—5 月中旬)。第 III 阶段为生长中期,时间跨度约在 6 月中旬— 8 月中旬,该时期内LAI 达到全年最高水平且变化相对较小,并出现第2 次生长高峰(6 月中旬—6 月末),但生长速度从 7月中开始快速降低。第 IV 阶段为生长后期,时间约为8 月中旬— 10 月末,这个时期林木基本不再生长,同时开始大量落叶,林分 LAI 快速减小。
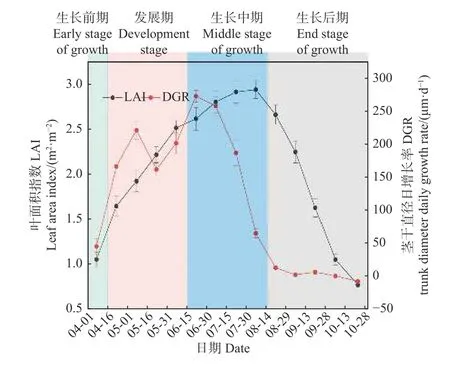
图2 2~6 年生毛白杨人工林平均茎干直径日增长率(DGR)和叶面积指数(LAI) 的季节动态Fig. 2 Seasonal dynamics of the average trunk diameter daily growth rate (DGR) and leaf area index (LAI) in 2-6 years old Populus tomentosa plantations
3.2 林分蒸腾、蒸发和蒸散动态
不同林龄林分生长季内的蒸发、蒸腾及蒸散量变化见图3。2~6 年生林分生长季内的累积Es分别为250、200、198、179 和255 mm,在林木栽植后的前5年,Es随林龄增大而呈非线性逐渐下降,但在栽植后的第 6 年即轮伐期末,Es又出现明显升高(图4)。年际尺度上,Es对ET 的贡献比例随林龄增大而非线性逐渐下降(P= 0.038),并在 5~6 年生林分中趋于相对稳定(图4)。在年内尺度上,Es对ET 的贡献存在季节变化,且变化模式因林龄而异(图3)。2 年生林分中,Es在整个生长季的ET 中几乎均占主导地位;随着林木发育,林分在 3 年生时达到郁闭,因而除了生长季和/或生长季末,其他时期内Es在ET 中的占比均较小,且林龄越大该趋势越明显。此外,2 年生 和 6 年生林分中,Es的季节变化与 ET0呈显著的正相关关系(P< 0.05),但在 3~5 年生林分中相关性不明显(P>0.05)。
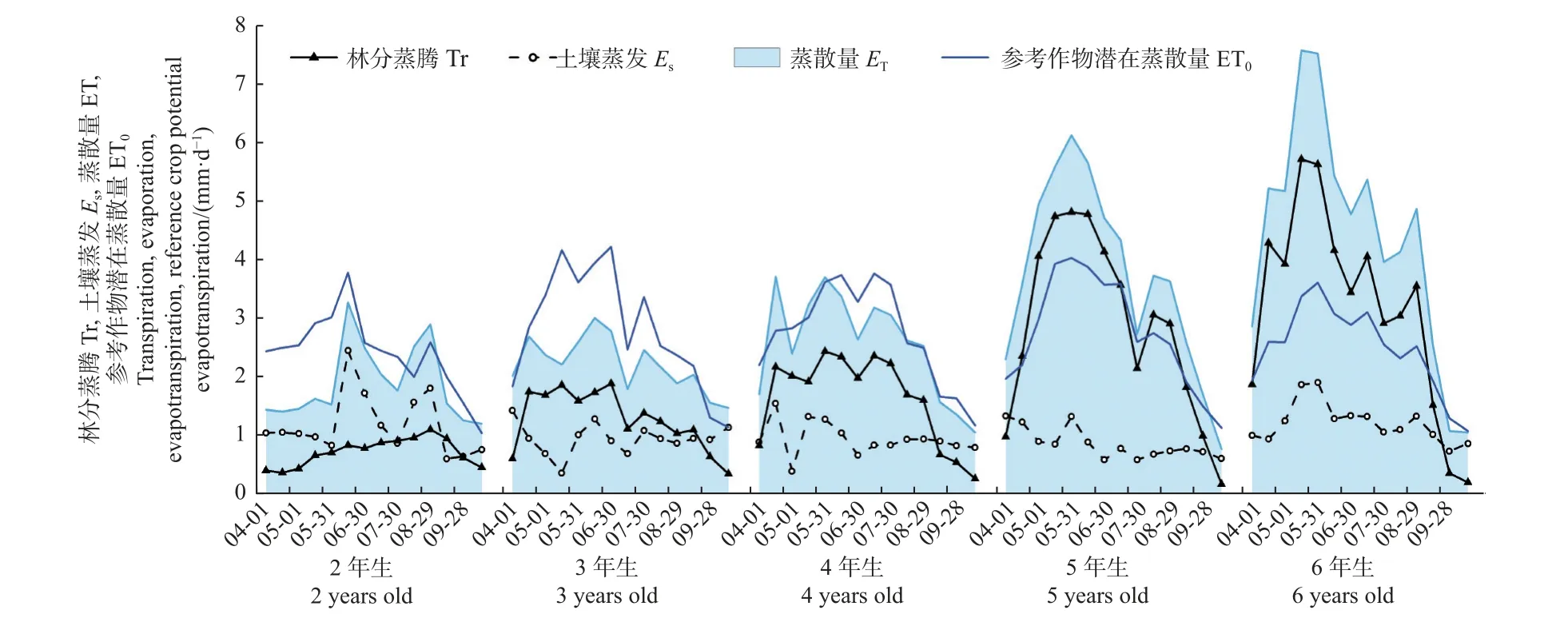
图3 2~6 年生毛白杨林分土壤蒸发(Es)、林木蒸腾(Tr)、林地蒸散量(ET)及参考作物潜在蒸散量(ET0)变化Fig. 3 Variations of soil evaporation (Es), tree transpiration (Tr), stand evapotranspiration (ET), and reference crop potential evapotranspiration(ET0) in 2-6 years old P. tomentosa plantations
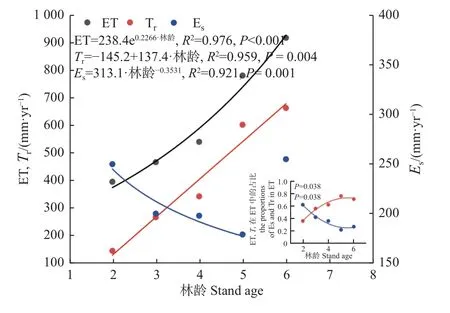
图4 毛白杨林分林地蒸散量(ET)、土壤蒸发(Es)及其在ET 中的占比、林木蒸腾(Tr)及其在 ET 中的占比随林龄的变化Fig. 4 Variation of stand evapotranspiration (ET), soil evaporation(Es) and its proportion in ET, and tree transpiration (Tr) and its proportion in ET with stand age
2~6 年生林分的累积Tr 分别为145、267、343、603 和664 mm,随林龄增加而线性增大(P= 0.004),但其在林分ET 中的占比则随林龄变化而非线性增大并在 5~6 年生林分中保持相对稳定(P= 0.038),2~6年生林分中的该占比分别为37%、57%、63%、77%和72%(图4)。Tr 在生长季内的变化规律随林龄而异(图3)。其中,在2 年生林分内,Tr呈缓慢上升趋势,直至9 月中下旬大量落叶后逐渐下降。3 年生和4 年生林分进入生长季后,其Tr 在约半个月内即快速升高至全年较高水平,之后在一定范围内稳定波动,并分别从7 月上旬和 8 月上旬开始逐渐下降。5~6 年生林分的Tr在进入生长季后同样迅速上升,但上升期明显延长,直至在 5 月下旬达到全年最高峰后保持稳定1~1.5 个月,然后逐渐波动下降。总体而言,Tr 的季节变化在 2 年生林分中与ET0的季节变化无关(P>0.05),但在 3~6 年生林分中则显著正相关(P< 0.05)。
年际尺度上,2~4 年生林分的ET 从低于ET0到逐渐与其相当,随后5~6 年生林分ET 明显高于ET0,且高出程度逐年增大(图3);2~6 年生林分的累积ET 分别为396、467、541、782 和919 mm,在整个轮伐期内随林龄增大而呈指数增加(P< 0.001)(图4)。年内尺度上,林分 ET 的动态变化与ET0基本一致,二者间存在显著的正相关关系(P< 0.05),且协同程度逐年升高(回归方程的R2逐年增大)。整个轮伐期内,ET 均在6 月达到最大;在毛白杨生长最快的5—7 月(图1),林分 ET 占生长季总 ET 的比例在2~6 年生林分中分别为47%、48%、52%、61%和59%;在历年主要生长阶段(图1)的累积ET 占生长季总ET 的58%~76%。
3.3 林分(基础)作物系数动态
不同林龄林分的作物系数季节动态差异明显(图5a):2 年生林分中,Kc在生长季内整体呈阶梯状逐渐升高,但在3 年生林分中,Kc则呈“下降—稳定—逐渐升高”变化;4~6 年生林分的Kc季节动态大体一致,呈双峰变化,且在生长后期快速下降。依据划分的4 个生长阶段(图2),构建 2~6 年生林分的作物系数曲线(图6),利用该曲线可获取不同林龄林分每半月尺度的作物系数。此外,不同林龄林分各生长阶段的平均作物系数见表1,发展期和生长中期的平均Kc均随林龄增加而逐渐增大(P< 0.05),但生长初期和末期则未出现该现象(P> 0.05)。
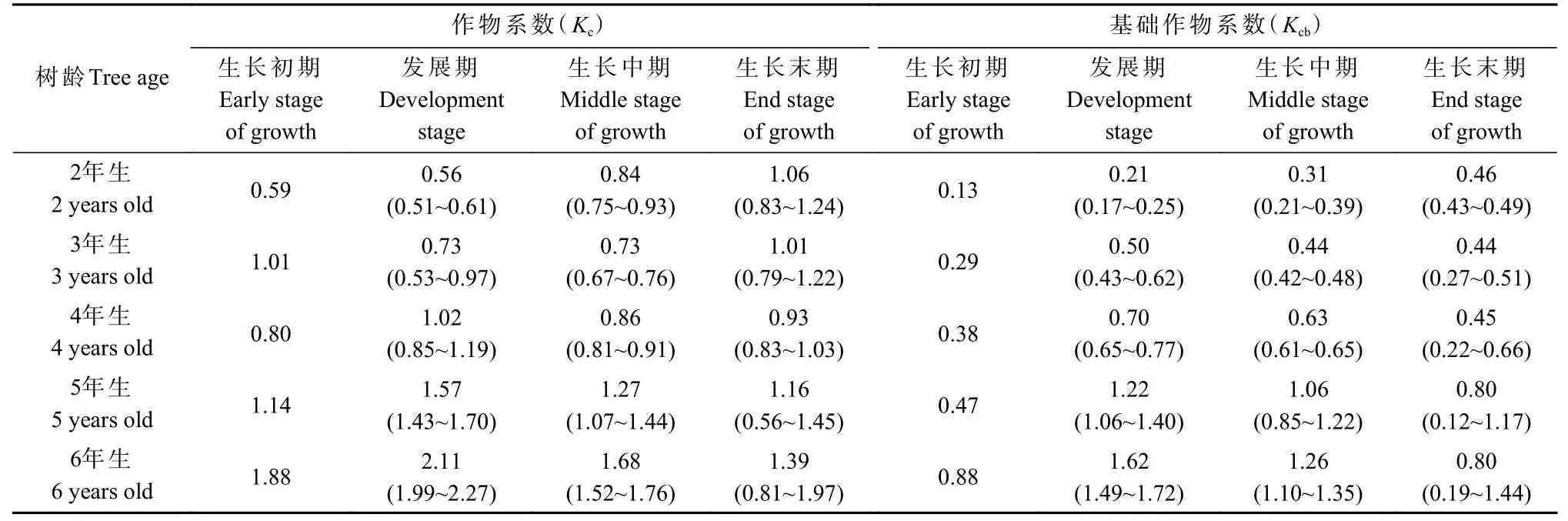
表1 2~6 年生毛白杨林分不同生长阶段的平均作物系数(Kc)及基础作物系数(Kcb)①Tab. 1 Average crop coefficient (Kc) and basal crop coefficient (Kcb) at different growth stages in 2-6 years old Populus tomentosa plantations

图5 2~6 年生毛白杨人工林作物系数(Kc)和基础作物系数(Kcb)的季节动态Fig. 5 Seasonal dynamics of crop coefficient (Kc) and basal crop coefficient (Kcb) in 2-6 years old P. tomentosa plantations

图6 2~6 年生毛白杨林分作物系数(Kc)曲线Fig. 6 Crop coefficient (Kc) curves in 2-6 years old P. tomentosa planations
毛白杨基础作物系数的季节动态在不同林龄林分中也存在明显差异(图5b)。2 年生林分中,Kcb逐渐升高,直至进入生长后期以后保持相对稳定。3~ 4 年生林分的Kcb季节动态呈单峰变化:进入生长季后,Kcb快速升高,于4 月中旬— 4 月末即达到全年最大值,在 5 月逐渐降低后保持相对稳定,并在9 月中之后再次下降。5 年生和6 年生林分的Kcb整体呈双峰变化,且Kcb在达到第1 个峰值后开始快速下降,直至第2个峰值出现。基于Kcb变化规律,构建 2~6 年生林分的基础作物系数曲线(图7),由此可获取各林龄林分每半月尺度的Kcb信息。通过表1 可知,除生长末期外,其他3 个生长阶段的平均Kcb均随林龄增加而逐渐增大(P< 0.05)。
3.4 林分(基础)作物系数与叶面积指数和地下水位的关系
林分Kc和Kcb与LAI 间的关系会随林分发育而变化(图8)。林分郁闭前(2 年生),Kc与 LAI 间无关系(P> 0.05),而郁闭后(3~6 年生),LAI 的变化能对Kc产生显著影响,但影响方式并不固定:3 年生林分的Kc随 LAI 增大而非线性降低(P< 0.05),4 年生林分中Kc则随 LAI 增大呈先升高后降低的变化趋势(P< 0.05),5~6 年生林分,Kc随 LAI 增大而增大,但当LAI 达到某临界值后,Kc趋于稳定(P< 0.001)。总体而言,3~4 年生和5~6 年生林分Kc随LAI 的变化规律分别可用二次多项式函数和 S 型函数描述,其决定系数R2分别为 0.42~0.44 和 0.71~0.77(图8)。
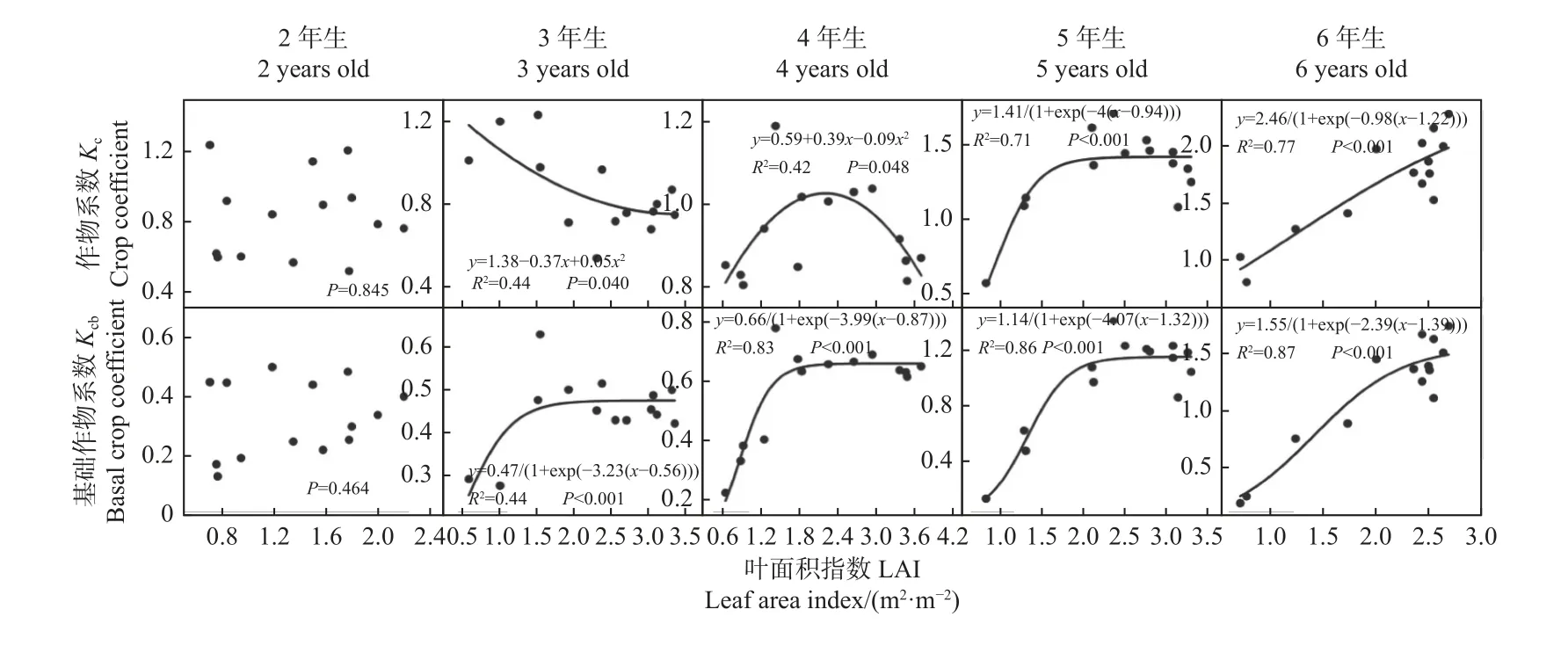
图8 2~6 年生毛白杨林分叶面积指数(LAI)与(基础)作物系数(Kcb, Kc)的关系Fig. 8 The relationships between (basal) crop coefficient (Kcb, Kcb) and leaf area index (LAI) in 2-6 years old P. tomentosa plantations
林分郁闭前,Kcb的变化同样不受 LAI 的影响(P> 0.05),但林分郁闭后则受其明显控制(P< 0.001)(图8)。在 3~6 年生林分中,Kcb均随 LAI 增大呈现先升高再趋于稳定的变化趋势,且Kcb开始趋于稳定的LAI 临界值以及趋于稳定的Kcb峰值有逐年增大趋势;Kcb与 LAI 间的共变规律均可很好地用 S 型函数来模拟,模型决定系数R2达到 0.83~0.87。
线性回归分析结果表明,整个轮伐期内,各林龄林分的Kc和Kcb均与地下水位之间在半月时间尺度上无显著相关关系(P> 0.05)。
4 讨论
4.1 毛白杨人工林生长发育特征
在准确了解林木生长动态基础上进行生长发育阶段划分,是构建林木(基础)作物系数曲线的前提,同时也能为人工林水分管理关键期的确定提供重要信息(Allenet al., 1998)。传统林木生长动态研究多是借助人力测定树干直径,时间分辨率低,同时,研究时间也大多较短(如1~2 个生长季),因此,对林木季节生长动态的了解可能并不全面和准确(王文全等,2001; Xiet al., 2014; Coyleet al., 2016)。本研究借助茎干直径微变化传感器,获取了长时间尺度(5 年)的高时间分辨率(天尺度)直径生长数据,并对整个轮伐期的生长动态进行分析(图1)。研究表明,每年 5—7 月是毛白杨生长最快的时期,占年总生长量的 69%~88%。该发现与 Xi 等 (2014)、王文全等(2001)和王烨等(2023)借助传统树木直径监测方法,针对1~2 个生长季观测得到的毛白杨速生期相一致。由此表明,每年5—7 月可作为毛白杨水分管理的关键时期。此外,本研究还发现,毛白杨每年的主要生长阶段长度并不一致,主要体现在4 月和 8 月的上半月是否有明显生长(图1)。本研究目前还无法解释该现象,但可推测,如果能通过调控措施在每年均维持这2 个时段的明显生长量,则能提高轮伐期内主要生长阶段的累积长度,从而进一步提升林分生产力。
目前,林木生长发育阶段有多种划分方法,且划分标准也较多(根据树干生长节律、叶片发育动态、枝条生长等)(裴保华, 1963)。例如,Li 等(2020)利用Gompertz 模型,根据树干直径日增长量,将2~3 年毛白杨生长阶段划为 4 个时期:早春出芽期(约 4 月初—4 月中旬)、春夏快速生长期(约 4 月中旬—6 月上旬)、夏秋缓慢生长期(约 6 月中旬— 10 月上旬)和冬季准备期(10 月中旬—10 月末)。裴保华等(1987)通过定期测定胸径,将 8~10 年生毛白杨的直径生长阶段划分为 5 个时期:早春缓慢生长期(4 月初—4 月底或 5月初)、春季速生期(4 月下旬或 5 月初— 6 月初或中旬)、春夏缓慢生长期(6 月初或中旬—7 月上旬)、夏季速生期(7 月上旬或中旬—8 月上旬)和夏秋缓慢生长期(8 月上旬— 10 月上旬)。与以上研究不同,本研究结合整个轮伐期内每年的叶面积指数和林木生长动态,根据 FAO-56(Allenet al., 1998)中的植被生长阶段划分方法,将毛白杨生长发育阶段划分为 4 个时期:生长初期、发展期、生长中期和生长末期(图2)。该划分方法不仅考虑了不同时期林分的冠层发育特征,而且也与前人的划分在一定程度上保持一致。例如,本研究中的生长初期与 Li 等 (2020)中的早春出芽期以及裴保华和等(1987)中的早春缓慢生长期基本对应;发展期与裴保华和等(1987)中的春季速生期大体一致;生长中期则包含了裴保华等(1987)中的春夏缓慢生长期和夏季速生期。因此,本研究划分的生长阶段同时兼顾了轮伐期内毛白杨林分的平均冠层发育和树干直径生长特征,不仅可用于(基础)作物系数曲线制定,而且也可很好地用于水分管理等集约培育措施的制定和优化。
4.2 毛白杨人工林长期耗水规律
杨树人工林耗水规律已有大量工作开展,但受限于研究方法,大多研究只获取了林分耗水单一组分的变化规律。例如,Gochis 等(2000)和Hou 等 (2010)分别利用土壤水量平衡法和波文比法仅研究了杨树人工林的蒸散季节动态特征;Navarro 等 (2018)和 Du 等(2021)利用树干液流法仅分析了杨树蒸腾的动态变化规律。有些研究综合利用微型蒸渗仪法与树干液流法获取了土壤蒸发、林木蒸腾和林分蒸散等杨树人工林的耗水多组分信息,但均局限在1~2 个生长季的短时间内(刘晨峰等, 2009; Xiet al., 2017; Cañete-Salinaset al., 2021; Songet al., 2021)。生产实践中,以木材生产为目的的人工林均以轮伐期为经营周期开展经营活动,而轮伐期长度因树种和目标收获材种而异,少则5~8 年(如杨树和桉树纸浆林),多则 15 年以上(如杨树大径材林、油松(Pinustabuliformis)用材林),因而仅获取短期内的林分耗水或其个别组分信息对于用材林科学经营的作用十分有限。为此,本研究利用微型蒸渗仪法和树干液流法,针对毛白杨纸浆林,率先在一个完整的轮伐期尺度上详细研究了林分耗水多组分的变化规律,可为短轮伐毛白杨人工林的全生命周期经营提供重要理论依据。
由于多关注耗水个别组分和研究周期较短,目前对杨树人工林耗水规律的了解多局限于特定林龄的生长季尺度,而对其耗水特征(总量、季节动态等)的年际变化或随林龄发育的变化还知之甚少。与其他研究结果相同(刘晨峰等, 2009; Houet al., 2010; Xiet al., 2017),本研究数据显示杨树林分ET 的季节动态与 ET0相一致,并同时进一步发现这种协同性并不受林分发育的影响(图3)。但是,毛白杨林分 Tr、Es和ET0的变化协同性则会随林分发育而改变:对于 Tr,林分郁闭前,两者的变化间无关系(P> 0.05),但郁闭后,则显著线性相关(P< 0.05),且相关程度逐年增大;对于Es,两者的显著协变关系(P< 0.05)仅出现在林分郁闭前和轮伐期末(6 年生林分)。由此表明,大气蒸发潜力能对林分耗水的各组分动态产生显著影响,但影响模式受林分发育控制:林分郁闭前,主要影响ET 和Es的季节动态,但郁闭后则主要影响 ET 和 Tr,并在轮伐期末能对三者均产生显著影响。
林分耗水总量随林龄增大会发生明显改变。众多研究表明,林龄越大,耗水总量越高(Xiet al., 2017;Gochiset al, 2000; Songet al., 2021),但也有研究发现林分耗水量随林龄增大并无明显升高趋势,仅呈波动变化(韩辉等, 2020; Ouyanget al., 2022),这可能是因为研究对象所处的林分发育阶段(如幼林或成熟林)或表征林分耗水的指标(如蒸腾或蒸散)不同所造成。与以上研究不同,由于本研究对耗水多组分进行了长时间观测,所以获取了耗水多组分随林龄的变化规律信息。 毛白杨林分的ET 和 Tr 随林龄增大而逐渐升高,但二者分别呈非线性和线性升高(图4)。与之不同,林分的年总Es随林龄增大而非线性下降,但其在轮伐期末的 6 年生林分中又出现明显升高,且达到接近 2 年生林分的水平,这可能是 6 年生林分中出现较强的自然整枝以及当年遭受风灾导致叶面积指数明显下降,冠层透光率突然升高所致。与Tr 和Es总量的逐年变化规律不同,Tr 和Es对林分ET 的贡献率随林龄增大分别呈非线性升高和降低趋势,二者在 5~6年生林分中趋于稳定,但 ET 贡献率是否会随林龄的继续增大而发生改变,还需更加长期的观测数据予以证明。综上,本研究揭示的林分耗水及其组分在整个轮伐期内的年际变化规律,可为人工林集约培育措施的优化及其耗水预测模型的构建提供关键信息。
4.3 毛白杨人工林(基础)作物系数曲线
在经营人工林时,由于林地面积一般较大,所以水分管理等经营措施的调整和实施的时间周期通常较长。鉴于此,本研究以每半个月为1 个周期来计算(基础)作物系数的平均值,并依此构建(基础)作物系数曲线及其预测模型。这种处理方式也符合FAO-56(Allenet al., 1998)所提到的“计算耗水量的时间步长应与获取(基础)作物系数值的时间步长相一致”原则。
毛白杨林分(基础)作物系数的季节动态变化规律随林分发育而变异较大(图5),该现象产生的重要原因之一是Kc和Kcb与 LAI 间的协变关系在不同林龄林分中存在较大差异(图8)。另外,与 FAO-56 的描述规律相一致,许多研究观测到的Kc和Kcb季节动态为:生长初期较低,在发展期逐渐升高,然后在生长中期达到高峰并保持稳定,最后于生长末期逐渐降低(Allenet al., 1998; Gochiset al., 2000; Gonget al., 2007;Dinget al., 2015; Zhaoet al., 2018)。与这些发现不同,本研究在任何林龄林分中均未观察到类似的季节变化规律,且主要体现在Kc和Kcb在郁闭林分的发展期即达到全年最高水平,但在生长中期却出现降低或维持在发展期的水平(图5)。这种结果差异可能是由于“(基础)作物系数与 LAI 间定量关系不同”和“冠层导度季节动态不同”的综合作用,如果Kc、Kcb与LAI 间存在线性关系,则Kc和Kcb在生长季内会逐渐缓慢升高,并在生长中期达到最高峰(Gonget al.,2007);但如果Kc、Kcb与 LAI 间存在S 型的非线性关系,则Kc和Kcb的最高峰会提前出现在林木发展期,如本研究中的毛白杨(图5, 图8)。此外,毛白杨林分在生长中期为防止木质部栓塞会关闭部分气孔,从而导致冠层导度下降和蒸腾下降(图3)(赵文芹等,2021),因此在郁闭林分的生长中期才会出现Kc和Kcb的明显降低现象(图5, 表1)。
本研究结果中,Kc和Kcb季节动态并不符合FAO-56 中所描述的常见规律,因此,无法采用 FAO-56 建议的方法来构建林分的(基础)作物系数曲线。本研究在综合数据变化规律和划分的生长阶段,并考虑使用方便性的基础上,采用手工拟合法构建了短轮伐毛白杨人工林的(基础)作物系数曲线图(图6, 图7)。在生产实践中,利用该图时应按照以下几点:首先,需要查阅某“半月”时期中心位置对应的曲线数值得到该时期的Kc和Kcb;其次,利用当地多年的气象数据算出该时期的多年平均ET0,或利用当年的气象数据算出该时期的实际 ET0;第三,将多年平均 ET0与Kc和Kcb相乘,可大致预测某一林龄林分在该时期的可能蒸散量和蒸腾量,或将当年实际ET0与Kc和Kcb相乘,得到当年该时期林分的实际蒸散量和蒸腾量;最后,利用上步获得的耗水量信息确定对应时期的灌溉量。此外,为了更加便于不同类型的生产人员进行毛白杨人工林的科学水分管理,本研究也绘制了不同林龄不同生长发育阶段的毛白杨Kc和Kcb平均值列表(表1),同时针对 3~6 年生林分构建了基于 LAI 的Kc和Kcb预测模型(图8)。在生产应用时,可根据实际情况进行方法选择,但需注意的是,本研究构建的(基础)作物系数曲线、列表和预测模型,仅适用于相似气候条件下以典型纸浆林栽植模式(密度 2 m×3 m)营建的短轮伐毛白杨人工林。
5 结论
结合树干液流法和微型蒸渗仪法,针对短轮伐毛白杨人工林,在其整个轮伐期尺度上系统研究了林分的长期耗水规律。林分蒸散、蒸腾和蒸发的季节动态特征存在年际变化,大气蒸发潜力的波动是控制耗水季节动态的重要因子,但控制方式因耗水组分和林龄而异。林分蒸散量和蒸腾量随林龄增大均逐渐升高,但升高方式分别呈非线性和线性;林分蒸发量随林龄增大先逐渐下降再明显升高,但其在蒸散中的占比则一直呈非线性降低。毛白杨生长阶段可划分为 4 个时期,且每年 5—7 月为林分水分管理的关键时期。林分(基础)作物系数的季节变化不受地下水位影响,而受叶面积指数控制,但该控制作用仅出现在林分郁闭后。此外,构建的毛白杨人工林(基础)作物系数曲线、列表和预测模型,可用于林分耗水量的估算,进而精准确定灌溉量。本研究发现将有助于短轮伐毛白杨人工林水分管理策略的制定和优化,并为深化认识人工林的长期耗水规律提供重要理论支撑。
猜你喜欢
防护林科技(2020年10期)2020-12-22
新农民(2020年21期)2020-12-08
防护林科技(2020年6期)2020-08-12
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
东坡赤壁诗词(2018年1期)2018-03-31
河南林业科技(2017年1期)2017-06-10
广东农业科学(2016年7期)2016-10-08
防护林科技(2016年9期)2016-09-27
防护林科技(2016年5期)2016-09-05
