
香芹酚对肉兔生长性能、养分表观消化率及肠道形态、短链脂肪酸含量和菌群相关指标的影响
2023-10-29 04:59武殿阁江皓天樊佳奇周思源刘树栋陈宝江
畜牧兽医学报 2023年10期
武殿阁,夏 苗,颜 安,江皓天,樊佳奇,周思源,韦 旭,刘树栋,陈宝江
(河北农业大学动物科技学院,保定 071000)
近年来,随着我国实施饲料全面禁抗政策,家兔养殖面临着巨大的挑战,由多种因素引起的兔腹泻和肠道炎症等疾病,严重制约着兔养殖行业发展[1]。现阶段,寻找新型畜禽饲料添加剂替代抗生素成为研究热点。在众多新型饲料添加剂中,越来越多的研究以天然植物产品为基础展开,有研究表明,天然植物产品是一类绿色安全的产品,并具有促进动物的生长发育、肠道健康和肠道菌群稳定的功能[2]。香芹酚是一种液体酚类物质,它主要存在于牛至、百里香、胡椒草和佛手柑等植物中[3]。香芹酚不仅可以通过氢键与细胞膜相互作用,增加细胞膜和线粒体的渗透性,从而起到抑菌抗菌的作用[4],且可以通过延长机体维生素C的活性从而提高机体免疫力[5],同时,香芹酚还可以通过抑制促炎因子(IL-1β、IL-6、IL-8和TNF-α)表达以及相关炎症信号通路(TLRS/NF-κB)的激活来缓解机体炎症反应[6]。目前,以香芹酚为添加剂在家兔生产上的研究应用鲜有报道。因此,本试验拟通过在肉兔饲粮中添加不同剂量的香芹酚,研究其对肉兔生长性能、养分表观消化率、肠道形态结构、短链脂肪酸、肠道菌群结构和肠道菌群代谢通路的影响,探索其在肉兔饲粮中的应用效果,旨在为肉兔无抗饲粮的研发和绿色养殖提供新途径和理论依据。
1 材料与方法
1.1 试验饲粮
香芹酚由北京菲迪饲料科技有限任公司提供(包被,粉末状,有效含量≥25%),基础饲粮参照De Blas和Wiseman[7]推荐的肉兔饲粮营养水平进行配制,基础饲粮的成分和营养水平见表1。
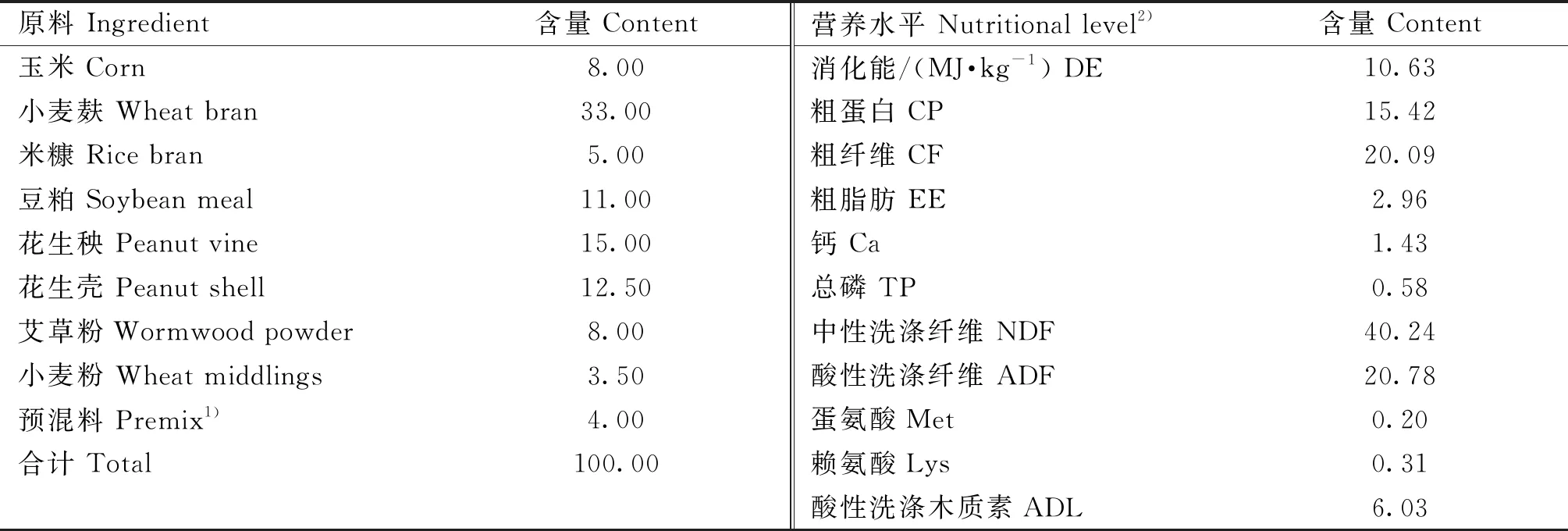
表1 基础饲粮组成及营养水平(风干基础)
1.2 试验设计与饲养管理
选取35日龄平均体重为1.25 kg的健康肉兔160只,随机分成4组,每组40个重复,每个重复1只。对照组(CON组)饲喂基础饲粮,试验组(T1、T2和T3组)饲喂在基础饲粮中添加100、200和300 g·t-1香芹酚(香芹酚有效含量分别为25、50和75 g·t-1)的试验饲粮。预试期5 d,正试期28 d,试验期间自由采食和饮水。
1.3 样品采集与处理
试验结束后,每组随机选取5只肉兔进行安乐死(戊巴比妥腿肌注射200 mg·kg-1体重),截取回肠肠段和盲肠肠段各一份,用磷酸缓冲液(PBS)冲洗干净后置于福尔马林溶液中,用于肠道结构形态的检测。用离心管采集盲肠食糜两份,于-80 ℃低温冰柜保存,一份用于盲肠挥发性脂肪酸的检测,另一份用于盲肠菌群结构的测定。
1.4 指标测定与方法
1.4.1 生长性能的测定 试验开始和结束时分别对各组试验肉兔进行空腹单只称重(禁食不禁水12 h),并于试验期记录各组试验肉兔饲喂量和剩余料量,计算平均日采食量(ADFI)、平均日增重(ADG)和料重比(F/G),同时记录各组试验肉兔的腹泻频率和死亡率。
平均日增重(ADG)=(试验兔末体重-试验兔初体重)/试验天数。
平均日采食量(ADFI)=试验兔总采食量/试验天数。
料重比(F/G)= 平均日采食量/平均日增重。
腹泻频率(%)= [(腹泻试验兔日只数/试验兔总只数)×试验天数]×100。
死亡率(%)=(死亡试验兔只数/试验兔总只数)×100。
1.4.2 养分表观消化率测定 正试期第21天,每组选取6只体重相近的健康试验肉兔进行消化代谢试验。消化代谢试验采用全收粪法连续收集7 d新鲜兔粪,并置于-20 ℃冰箱保存。消化代谢试验期间记录每只肉兔每天的实际采食量。参照《饲料分析及饲料质量检测技术》[8]的方法对饲粮和粪样中粗蛋白质(CP)、粗脂肪(EE)、粗纤维(CF)、中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)、钙(Ca)、总磷(TP)、总能(GE)和干物质(DM)的含量进行测定,计算养分表观消化率。
养分表观消化率(%)=((料中该养分DM×采食量)-(粪中该养分DM×粪重))/(料中该养分DM×采食量)×100。
1.4.3 肠道形态测定 将回肠和盲肠组织于福尔马林中固定48 h后,通过石蜡包埋切片,HE染色后用中性胶包封。在光学显微镜(Eclipse E100型,尼康日本)和成像系统(Nikon DS-U3型,尼康日本)下观察组织形态并扫描,用Case viewer软件测量回肠绒毛高度(VH)、绒毛宽度(VW)、隐窝深度(CD)、肠黏膜厚度(MT)、盲肠肠腺深度(DIG)和肠黏膜厚度(MT),并计算绒毛高度/隐窝深度比值(V/C)。
1.4.4 盲肠短链脂肪酸测定 将0.5 g盲肠食糜用蒸馏水稀释3倍,5 400×g离心10 min(4 ℃),取上清液1 mL,加入含内标2-乙基丁酸(2EB)的25%偏磷酸铵溶液0.2 mL,混匀,冰浸30 min,10 000×g离心10 min,去除样品中的蛋白沉淀。取上清液,采用岛津气相质谱仪(GC-2014型,岛津日本)对短链脂肪酸进行测定,具体检测步骤参照张勤[9]的方法进行。
1.4.5 盲肠微生物菌群结构测定 采用OMEGA Soil DNA试剂盒提取盲肠食糜中的微生物总DNA。以提取的总DNA为模板,采用正向引物338F(5′-ACTCCTACGGGAGGCAGCA-3′)和反向引物806R(5′-GGACTACHVGGGTWTCTAAT-3′)对微生物16S rRNA基因V3-V4区进行PCR扩增。送至上海派森诺生物科技股份有限公司,通过Illumina NovaSeq平台进行微生物16S rRNA高通量测序分析。并将16S rRNA基因序列在KEGG(https:∥www.kegg.jp/)数据库进行代谢通路预测。
1.5 数据统计与分析
试验数据采用SPSS 22.0软件进行单因素方差分析(one-way ANOVA),均值差异采用Duncan′s多重范围检验进行比较。数据结果以“平均值±标准误”表示,以P<0.05表示差异显著。
2 结 果
2.1 香芹酚对肉兔生长性能的影响
香芹酚对肉兔生长性能的影响如表2所示,T1组肉兔的末体重(FBW)显著高于CON组(P<0.05),T1和T2组的ADG和ADFI均显著高于CON组(P<0.05),T1组肉兔腹泻频率和死亡率均为0,显著低于CON组(P<0.05),T2组肉兔腹泻频率显著低于CON组(P<0.05)。
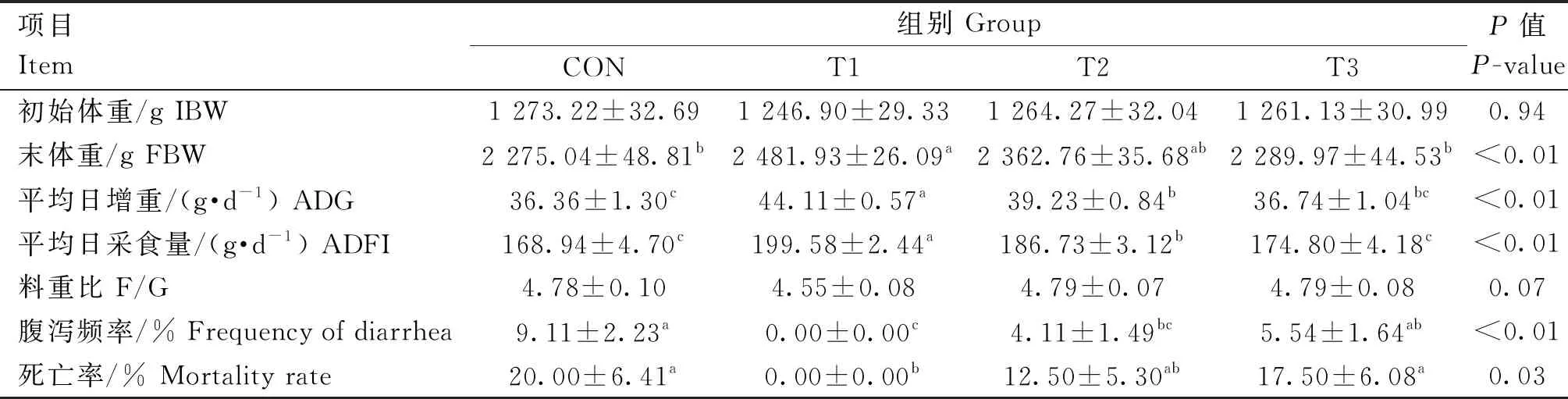
表2 香芹酚对肉兔生长性能的影响
2.2 香芹酚对肉兔养分表观消化率的影响
香芹酚对肉兔养分表观消化率的影响如表3所示,T1、T2和T3组的CP表观消化率均显著高于CON组(P<0.05),CF、NDF、ADF、TP和GE表观消化率均高于CON组,但未达到显著水平(P>0.05)。
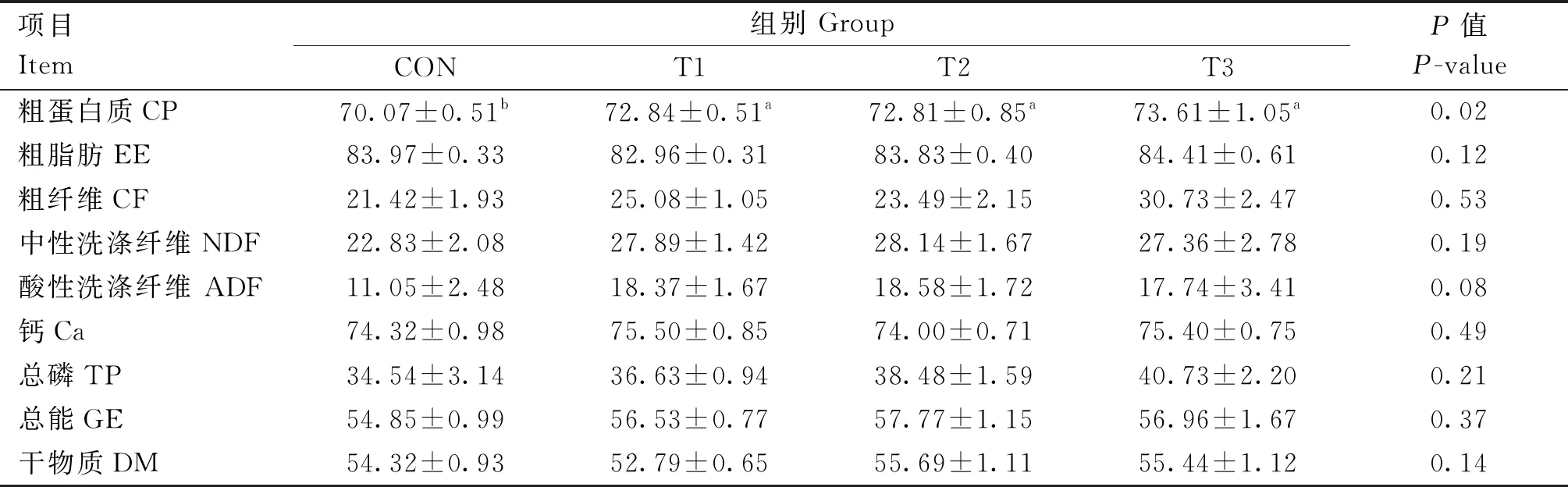
表3 香芹酚对肉兔养分表观消化率的影响
2.3 香芹酚对肉兔肠道形态组织结构的影响
香芹酚对肉兔肠道形态组织结构的影响如表4所示,T1、T2和T3组的回肠绒毛高度和肠黏膜厚度均显著高于CON组(P<0.05),其中T1组最高,分别达到了(665.00±23.27和822.50±25.62)μm,T1和T2组回肠的V/C显著高于CON组(P<0.05)。由图1可知,与CON组相比,T1、T2和T3组的回肠肠道绒毛排列更紧密。

表4 香芹酚对肉兔肠道形态的影响
2.4 香芹酚对肉兔盲肠短链脂肪酸含量的影响
香芹酚对肉兔盲肠短链脂肪酸含量的影响如表5所示,T1、T2和T3组肉兔盲肠乙酸含量高于CON组,但未达到显著水平(P>0.05),仅T1组丙酸含量高于CON组(P>0.05),同时,T1组丁酸的含量显著高于CON组(P<0.05)。
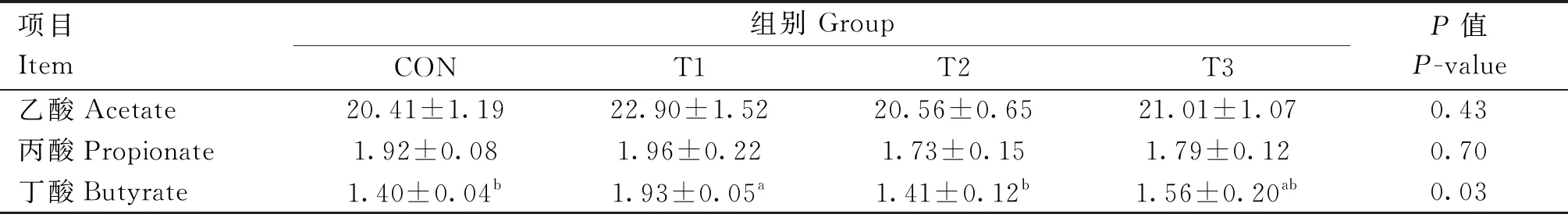
表5 香芹酚对肉兔盲肠短链脂肪酸含量的影响
2.5 香芹酚对盲肠菌群结构的影响
韦恩图(Venn)显示,CON、T1、T2和T3组中分别存在9 540、13 841、13 664和12 855个特定操作分类单元(operational taxonomic units,OTUs),四组共有OTU为1 244个(图1)。通过Chao1指数、Faith-pd指数、Shannon指数和Simpson指数评估单个样本内的微生物多样性,但未观察到组间的显著变化(表6,P>0.05)。对OTUs进行PCA分析,以评估样品和组之间的相似性和差异性(图2)。结果显示,CON组盲肠菌群与T1、T2和T3组有明显分离,但T1、T2和T3组之间没有明显分离。由图3可知,肉兔盲肠微生物在门水平上主要以厚壁菌门和拟杆菌门2种菌门组成,各组2种菌门相对丰度总和分别为84.16%、90.76%、87.92%和87.53%。对微生物菌群结构比例进一步分析结果见表7,与CON组相比,肉兔饲料中添加香芹酚可以显著提高肉兔盲肠中厚壁菌门的相对丰度(P<0.05);同时,肉兔饲料中添加香芹酚也会显著降低肉兔盲肠中拟杆菌门的相对丰度(P<0.05)。肉兔盲肠微生物属水平以微生物丰度前20作为各组的优势菌属进行分析(图4)。并由表8可知,与CON组相比,T1、T2和T3组的颤螺旋菌属的相对丰度显著升高(P<0.05)。
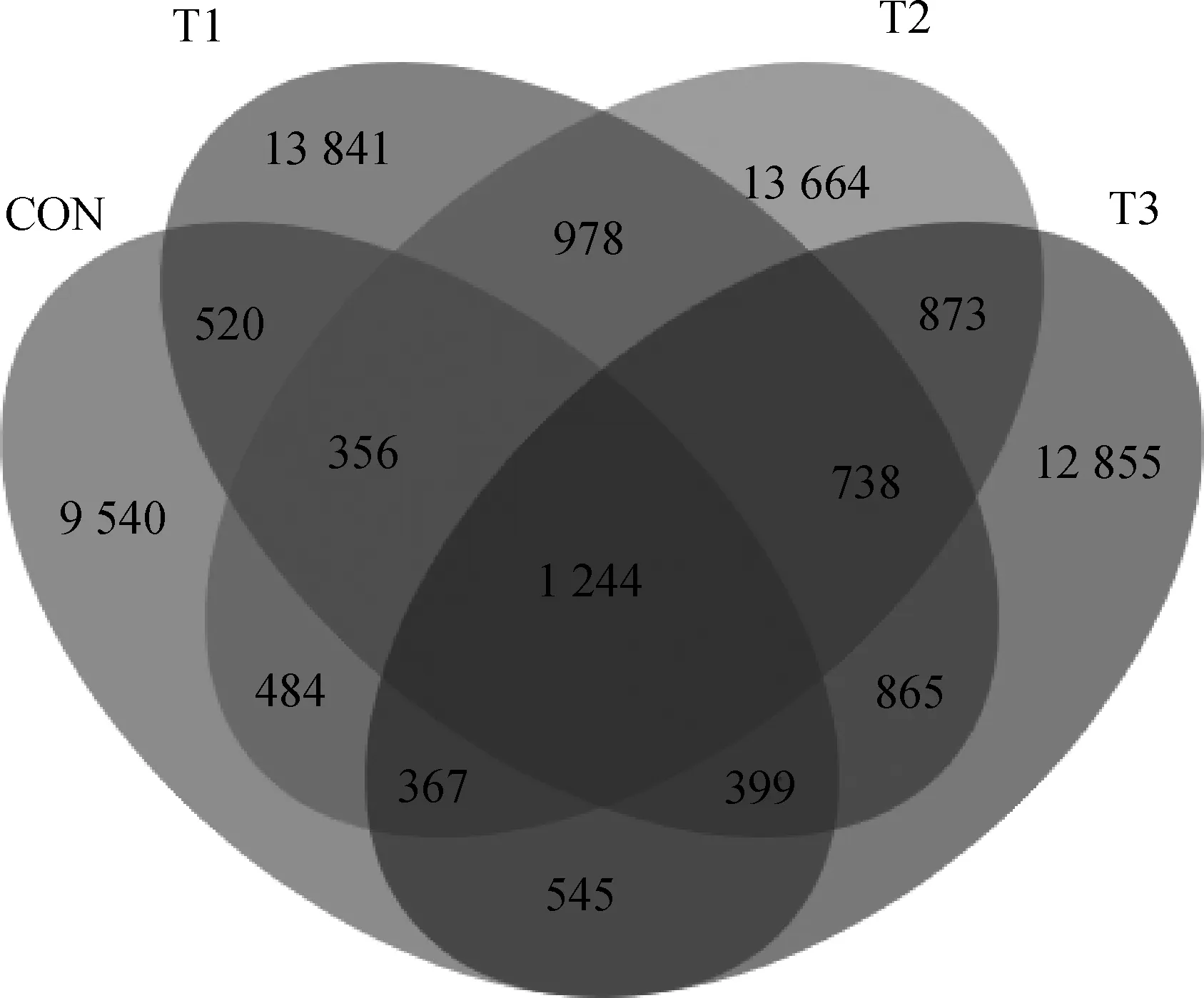
图2 特征值分布Venn图Fig.2 Eigenvalue distribution Venn diagram
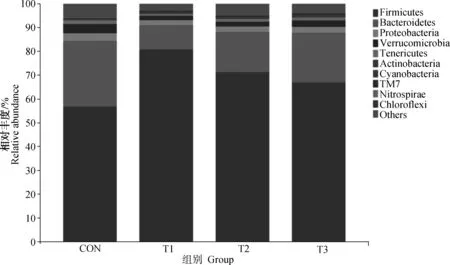
Firmicutes:厚壁菌门;Bacteroidetes:拟杆菌门;Proteobacteria:变形菌门;Verrucomicrobia:疣微菌门;Tenericutes:软壁菌门;Actinobacteria:放线菌门;Cyanobacteria:蓝细菌;TM7:TM7菌门;Nitrospirae:消化螺旋菌门;Chloroflexi:绿弯菌门;Other:其他图4 门水平上盲肠微生物菌群相对丰度Fig.4 Relative abundance of cecal microflora at phylum level

表6 香芹酚对肉兔盲肠菌群Alpha多样性指数的影响

表7 香芹酚对肉兔盲肠菌群门水平的影响

表8 香芹酚对肉兔盲肠菌群属水平的影响
2.6 添加100 g·t-1香芹酚对盲肠菌群代谢通路的影响
由图5可知,100 g/t香芹酚与对照相比可以显著上调的代谢通路为类固醇激素合成、糖胺聚糖的降解、次生胆汁酸合成、初级胆汁酸合成、泛素酮等萜类醌的生物合成、N-聚糖合成和酮体的合成与降解(P<0.05),鞘脂类代谢和脂多糖合成显著下调(P<0.05)。
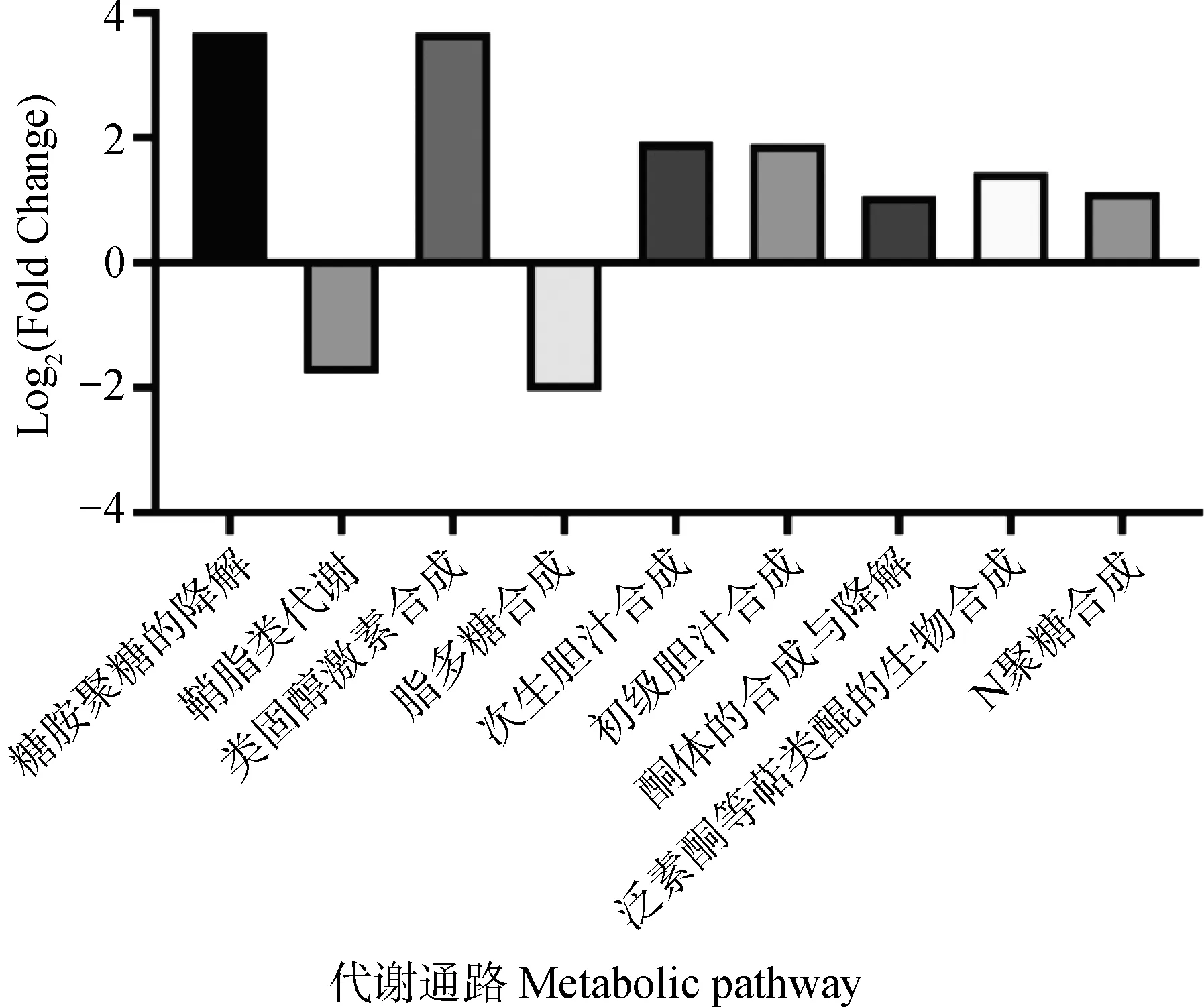
图6 香芹酚含量100 g·t-1对肉兔盲肠菌群代谢通路的影响Fig.6 Effects of 100 g·t-1 carvacrol on cecal microflora at metabolic pathway of rabbits
3 讨 论
3.1 香芹酚对肉兔生长性能的影响
动物在早期断奶阶段,容易受到日粮变化、环境、生理和心理因素的影响,出现生长缓慢、腹泻、肠屏障功能受损等断奶应激问题,对动物的健康和生长产生负面影响[10-12]。日增重和饲料转化率是衡量动物生产性能的主要指标[13]。Hashemipour等[14]研究发现,在肉鸡饲粮中添加200 mg·kg-1香芹酚能够提高ADG。本试验发现,添加香芹酚可以提高肉兔的ADG和ADFI。Brenes和Roura[15]研究发现,香芹酚能够增强食物的风味,刺激动物的味觉系统,进而提高采食量。这表明,香芹酚可能通过增加肉兔采食量来提高生长速度。在本研究中,添加香芹酚可以显著降低肉兔的腹泻频率和死亡率。
Gatica等[16]研究发现,香芹酚能够有效抑制促炎因子IL-1β、IL-6、TNF-α的过度分泌,从而抑制肠道炎症的发生,并且TNF-α分泌量降低还可以促进肠道紧密连接蛋白的表达[17],以此来维持肠道物理屏障的稳定性。这表明,香芹酚可能通过维持肠道健康以降低肉兔的腹泻频率和死亡率。此外,525 μmol·L-1的香芹酚会诱导猪肠上皮细胞凋亡,进而影响动物肠道对营养物质的吸收利用[18],并且当大鼠口服810 mg·kg-1BW的香芹酚时,会导致大鼠死亡[19]。这表明,香芹酚的添加量过高可能会导致机体功能出现紊乱进而影响到动物的生长性能。本研究结果表明,饲粮中添加100 g·t-1的香芹酚可以显著提高肉兔的生长性能。
3.2 香芹酚对肉兔养分表观消化率的影响
养分表观消化率可以判断动物对养分的消化能力,该指标对动物的生产性能至关重要[20]。本试验发现,在肉兔饲料中添加香芹酚可以显著提高粗蛋白质的表观消化率。刘立山等[21]研究发现,在饲料中添加2 g·t-1的香芹酚,连续饲喂15 d后,可以显著提高绵羊对粗蛋白质的消化率,与本试验结果一致。肉兔在生长过程中,由于消化器官发育速度的影响,其所分泌的蛋白酶含量影响着对饲料中蛋白质的消化吸收[22]。当未被消化的蛋白质进入肠道后,经过微生物发酵可产生高浓度的氨、胺、苯酚和对甲酚等代谢物,进而对肠道黏膜起到损伤的作用[23]。刘福鑫等[24]研究发现,4.56%香芹酚包被粉制成150 g·t-1饲料可以显著提高肉兔肠道中胰蛋白酶和脂肪酶的活性,进而提高肉兔对粗脂肪的表观消化率。这表明香芹酚可能通过增加肠道中胰蛋白酶的活性来提高肉兔对粗蛋白质的消化率,并降低由未被消化的蛋白质对肠黏膜造成的损伤,但在本试验中并未发现香芹酚可以显著提高肉兔对粗脂肪的表观消化率,这有可能是肉兔日粮中香芹酚有效含量不同,对消化酶活性调节能力不同所致。此外,周瑞等[25]研究发现,香芹酚通过提高肠道中黄色瘤胃球菌、白色瘤胃球菌和产琥珀酸丝状杆菌的相对丰度的方式,促进绵羊对纤维素的吸收利用。在本试验中,香芹酚对肉兔的纤维素消化率并无显著促进作用,这可能是由于绵羊属于反刍动物而肉兔属于单胃动物,在香芹酚作用于肠道微生物时无法提高瘤胃球菌的相对丰度所致。本研究结果表明,饲粮中添加香芹酚可以显著提高肉兔粗蛋白质的表观消化率。
3.3 香芹酚对肉兔肠道形态组织结构的影响
肠道是动物体的消化器官,对食物的消化和营养物质的吸收至关重要[26]。肠道功能的增强可以使肠道菌群稳定和蠕动加快,从而促进肠道消化吸收能力,以提高动物的生长性能[27]。肠道绒毛高度、绒毛宽度和隐窝深度是判断肠道功能的重要指标[28]。肠绒毛高度和宽度的增加有利于肠黏膜与食糜充分接触,这有助于对营养物质的吸收,而隐窝深度降低表明肠道上皮细胞成熟度变高,代表吸收能力增强[29]。王红玉和黄丽萍[30]研究发现,日粮中添加香芹酚可显著提高肠绒毛高度、绒毛高度与隐窝深度比值和肠道肌层厚度。在本研究中,饲粮中添加香芹酚可显著提高回肠的绒毛高度和肠黏膜厚度,并且当添加量为100和200 g·t-1时,可显著提高绒毛高度与隐窝深度比值。香芹酚可通过平衡肠道菌群结构并促进上皮细胞的有丝分裂,增加上皮细胞数量,从而提高绒毛高度和宽度,进而改善营养吸收[31]。
3.4 香芹酚对肉兔盲肠短链脂肪酸含量的影响
短链脂肪酸(short-chain fatty acid,SCFA)是肠道菌群的代谢产物[32]。乙酸转化为乙酸盐,可以改善肠道上皮对肠道的保护功能[33],丙酸可以抑制肠道炎症的发生[34],丁酸转化为丁酸盐,不仅可以刺激动物肠道中黏蛋白的分泌,还可以直接为肠黏膜供能,以保证肠道结构的完整性和增强肠道的免疫功能[35]。周瑞[36]研究发现,牛至油(主要成分为香芹酚)可以显著提高羔羊盲肠乙酸含量,但在本研究中,肉兔饲粮添加香芹酚后,盲肠中丁酸含量显著提高。这表明,单纯的香芹酚添加剂与含有香芹酚的复合添加剂相比,对调控微生物代谢物可能具有一定的选择性。
3.5 香芹酚对肉兔盲肠菌群结构的影响
动物肠道的微生物与宿主相互作用紧密,对加快宿主的营养物质吸收速度和提高免疫功能等方面具有重要意义[37]。肠道菌群结构变化容易受到环境变化、日粮配比以及动物本身等影响[38]。在哺乳动物中,厚壁菌门和拟杆菌门作为肠道的优势菌群,约占肠道总细菌数的90%[39]。在本试验中,肉兔盲肠内主要的菌门为厚壁菌门和拟杆菌门,其中添加香芹酚可以显著提高厚壁菌门的相对丰度并降低拟杆菌门的相对丰度。Ley等[40]研究发现,随着体重指数的升高,厚壁菌门和拟杆菌门的比例也会随之增加,这可能是厚壁菌门与拟杆菌门相比,厚壁菌门吸收食物中热量更充分,且其使热量转为脂肪积累于皮下的效率更高[41]。这表明,饲粮中添加香芹酚增加了肉兔盲肠中厚壁菌门和拟杆菌门的比例可能是促进肉兔体重升高的原因。在本研究中,香芹酚可以显著提高盲肠中颤螺菌属的相对丰度。Xu等[42]研究发现,颤螺菌属的丰度与肠道炎症呈负相关,并且颤螺菌属可以产生丁酸[43],丁酸通过促进肠道紧密连接蛋白的表达,进而显著改善肠道上皮功能[44]。这表明,饲粮中添加香芹酚可以通过改善肉兔盲肠菌群结构,以使肉兔肠道处于健康的状态,进而提高肉兔的生长性能。
3.6 香芹酚对肉兔盲肠菌群代谢通路的影响
为进一步探讨香芹酚通过调整肠道菌群对肉兔生长性能的影响,采用菌群功能性预测进行分析,对具有显著差异的代谢通路进行筛选。通过对CON组和T1组进行分析,共鉴定出177个差异代谢通路,其中显著差异的代谢通路为9个,2个上调与生长发育相关,5个上调和2个下调与机体健康相关。糖胺聚糖参与众多的生物学功能,主要表现为,糖胺聚糖降解后与其结合蛋白之间的相互作用,决定了结合的特异性和选择性[45]。如:糖胺聚糖与生长因子结合,可以促进细胞的有丝分裂和细胞迁移[46]、糖胺聚糖与成形素结合可以促进组织的分化和发育[47]、糖胺聚糖与脂质蛋白结合可以促进脂质代谢[48]。鞘脂是一种脂质小分子,参与多种重要的生命活动[49]。鞘脂在代谢过程中,最核心的分子为神经氨酰[50]。神经氨酰作为脂毒分子之一,可引起大量细胞应激反应,导致细胞凋亡[51],并且神经氨酰也被认为是心血管疾病发生的标志物[52]。类固醇激素在不同的器官均介导着重要的发育和生理功能,包括生殖系统、骨骼生长、大脑的发育和脂肪的分化代谢[53]。当类固醇激素缺乏或异常时,会导致机体的生育系统受损、骨骼生长速度减缓以及癌症的发生[54]。脂多糖通常由革兰阴性菌裂解时被释放[55],当脂多糖进入机体后,会导致肠道菌群失衡、肠黏膜屏障受损、诱导炎症基因的表达,进而造成机体损伤[56]。胆汁酸通过肠道菌群来发挥作用[57],一般的过程为将肝产生的初级胆汁酸转化为次级胆汁酸,次级胆汁酸可以通过减少参与炎症的关键细胞因子和趋化因子的表达来减轻急性和慢性小鼠肠道炎症模型中的炎症[58]。酮体代谢是生理稳态的中心节点[59],在过去研究中,酮体可以缓解由氧化应激造成的神经元和心肌细胞中细胞损伤、死亡的情况[60]。泛醌在机体内的主要形式为辅酶Q10[61],当机体中缺乏辅酶Q10,会降低机体清除自由基的能力,使抗氧化能力受损[62]。N-聚糖可以在细胞表面与半乳糖凝集素相互作用,产生大分子晶格,从而影响细胞分化和凋亡状态[63]。N-聚糖的缺乏会影响前B细胞、未成熟B细胞和T2过渡外周B细胞的存活,进而影响机体的免疫能力[64]。本研究中,在肉兔日粮中添加100 g·t-1的香芹酚可以提高盲肠菌群代谢通路中糖胺聚糖的降解、类固醇激素生物合成、初级胆汁酸合成、次生胆汁酸合成、酮体的合成与降解、 泛素酮等萜类醌的生物合成和N-聚糖的合成,降低盲肠菌群代谢通路中鞘脂类代谢和脂多糖合成。这表明,100 g·t-1香芹酚可以通过调整肉兔盲肠菌群的代谢通路,影响肉兔的生长性能。
4 结 论
在肉兔饲粮中添加100 g·t-1的香芹酚可以显著提高肉兔的生长性能和粗蛋白质的养分表观消化率,促进肠道健康,改善盲肠菌群结构并对盲肠菌群的代谢通路起到正向调节的作用。
猜你喜欢
现代畜牧科技(2021年5期)2021-07-20
中国比较医学杂志(2020年4期)2020-05-26
水生生物学报(2019年4期)2019-07-20
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
中国农业信息(2016年8期)2016-12-01
湖南畜牧兽医(2016年1期)2016-06-05
花卉(2016年3期)2016-04-16
动物营养学报(2015年9期)2016-01-07
食品工业科技(2014年23期)2014-03-11
