
腐殖酸对盐胁迫下土壤理化性质、微环境及苦瓜生长的影响
2023-10-24 04:44陈星星刘新社王盛荣
江苏农业科学 2023年17期
陈星星 刘新社 王盛荣

摘要:为了探究腐殖酸对盐胁迫条件下苦瓜生长的缓解效应和根际土壤理化性质、土壤微环境的作用,以绿星2号苦瓜为试料,研究不同浓度腐殖酸对盐胁迫环境下苦瓜幼苗形态建成、根际土壤化学性质、微生物群落结构和土壤酶活性的影响。结果表明,盐胁迫能够抑制苦瓜幼苗形态建成,降低土壤养分含量,提高土壤pH值,抑制土壤微生物數量的增加和土壤酶活性的提高。腐殖酸能够缓解盐胁迫对苦瓜幼苗株高、茎粗、最大叶面积、根长、地上部鲜质量、地下部鲜质量的抑制作用,缓解效果随腐殖酸浓度的增加呈现先升高再降低的单峰变化规律,且以NHA3处理(100 mmol/L NaCl+160 mg/L腐殖酸)下的值最大,分别较CK2(100 mmol/L NaCl)增加33.98%、15.10%、34.61%、25.18%、22.16%、36.00%,与CK2差异均显著(P<0.05)。腐殖酸能够缓解盐胁迫对根际土壤速效氮、速效磷和速效钾含量的抑制作用,且随腐殖酸浓度的增加呈现先增加再降低的变化趋势,但是盐胁迫下的速效氮含量以NHA2处理(100 mmol/L NaCl+80 mg/L腐殖酸)最大,速效磷、速效钾含量均以NHA3处理最大。腐殖酸能够提高盐胁迫条件下的有机质含量,且有机质含量随腐殖酸浓度的增加而升高;腐殖酸能够降低盐胁迫条件下的土壤pH值,且土壤pH值随着腐殖酸浓度的增加呈下降趋势。腐殖酸能够缓解盐胁迫对土壤微生物数量的抑制作用,且微生物数量随腐殖酸处理时间的延后而增加。在盐胁迫环境下,细菌、真菌数量以NHA3处理最大,放线菌数量以NHA4处理最大,但与NHA3处理间差异不显著。腐殖酸能够提高盐胁迫环境下脲酶、蔗糖酶和磷酸酶的活性,且上述酶活性随腐殖酸处理时间的延后而提高。随腐殖酸浓度的增加,蔗糖酶活性呈现先升后降的变化趋势,磷酸酶活性呈现逐渐升高的变化规律。细菌、真菌、放线菌数量与速效氮、速效磷、速效钾、有机质含量之间存在正相关关系,与土壤pH值之间存在负相关关系。综合比较得出,NHA3处理下的腐殖酸浓度能够有效缓解盐胁迫对苦瓜幼苗生长的抑制作用,改善土壤肥力状况,增加根际土壤微生物数量,提高土壤酶活性,进而增强苦瓜的抗盐胁迫能力。
关键词:腐殖酸;盐胁迫;形态建成;理化性质;微生物;土壤酶
中图分类号:S154.4;S642.506 文献标志码:A 文章编号:1002-1302(2023)17-0138-07
近年来,随着农业供给侧结构性改革的推进,我国农业进入快速发展阶段,设施农业的地位也变得愈加重要,设施栽培面积也呈现逐年扩大的变化趋势。但设施栽培多为半封闭状态,常年得不到雨水淋洗,大量无机离子在土壤水分蒸发和作物水分蒸腾的双重作用下向上迁移,表现出显著的表聚特征,这容易导致土壤次生盐渍化现象的发生[1]。而生产者为了追求高产出,盲目加大施肥用量,过量的养分不能被植物吸收,又以无机盐的形式在土壤中积累,进一步加剧了土壤次生盐渍化进程。土壤发生次生盐渍化后质量严重退化,土壤板结、不易耕种,植株生长势减弱、病害加重、产量降低、品质变差。陈昆等研究得出,盐胁迫能够显著降低西瓜幼苗株高、茎粗、干鲜质量,抑制抗氧化保护酶活性和根系对矿质元素的吸收利用[2-3]。操庆等研究发现,盐胁迫能够降低设施土壤微生物量碳和氮含量,同时抑制土壤酶活性,降低设施土壤肥力,最终影响设施作物生长[4]。由此可见,土壤盐渍化可对土壤可持续生产能力构成威胁,已成为制约设施农业可持续发展的重要障碍。因此,如何缓解设施土壤盐胁迫危害,是当前设施农业发展面临的重要难题。
腐殖酸是一类成分复杂的天然有机物质,具有吸附性、胶体性、弱酸性、络合性和氧化还原性等特点[5],对改良土壤结构、改善土壤理化性质、提高肥料利用率、促进植物生长等方面有重要作用[6]。高云晓等研究得出,适量的腐殖酸能够增加刺槐植株叶片含水量,缓解盐胁迫对其造成的生理干旱,促进幼苗生长[7]。金萍等研究发现,腐殖酸可通过提高多酚氧化酶活力、降低脱落酸和丙二醛含量等途径,提高大豆抗盐碱胁迫的能力[8]。由此可见,腐殖酸在刺激植物生长、提高植株抗盐胁迫能力等方面有重要的研究和利用价值。然而,当前有关腐殖酸对盐胁迫环境下苦瓜生长及根际土壤微环境影响的研究尚未见报道。为此,本研究以苦瓜为试验材料,借助盆栽的方式模拟盐胁迫环境,研究不同浓度腐殖酸对盐胁迫环境下苦瓜幼苗形态建成、根际土壤化学性质、微生物群落结构和土壤酶活性的影响,探究腐殖酸提高苦瓜耐盐性的最佳浓度和改善土壤肥力状况的机制,揭示腐殖酸修复盐胁迫土壤的微生态机制,以期为设施苦瓜的耐盐优质高产可持续栽培提供参考依据。
1 材料与方法
1.1 供试材料
供试苦瓜品种为绿星2号;供试腐殖酸水溶肥由内蒙古际美生物科技有限公司生产。
1.2 试验设计
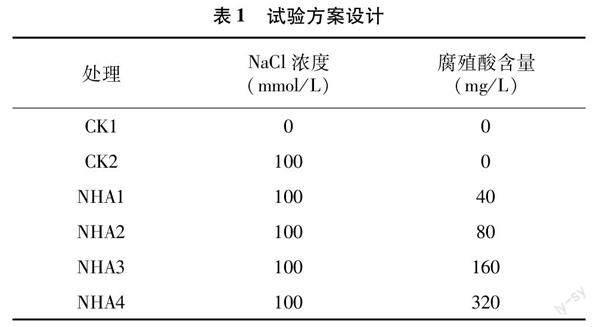
试验于2021年12月至2022年5月在河南省农业职业学院实验站人工气候室内进行。试验设置2个对照、4个处理,其中对照1(CK1)浇灌清水,对照2(CK2)浇灌100 mmol/L NaCl溶液,4个处理组按表1方案浇灌含NaCl和不同浓度腐殖酸的水溶液,分别记为NHA1、NHA2、NHA3、NHA4处理。试验用盆栽方式进行,花盆高20 cm,直径18 cm,每盆装大田土至15 cm高度。当苦瓜幼苗长至1叶1心期时,选取大小基本一致的苦瓜幼苗栽植于花盆中,每盆栽培1株。每个处理设置3次重复,每10盆为1次重复,共计180盆。栽植完毕后,按试验设计浇灌相应的水或溶液进行胁迫试验,浇灌时以水或溶液浸透80%以上的土壤且不从花盆底部渗出为准。
1.3 指标测定与方法
胁迫20 d时测定苦瓜幼苗的形态指标和生物量,胁迫15、30 d时测定土壤理化性质、土壤微生物数量和土壤酶活性。株高、根长采用直尺测量,茎粗采用游标卡尺测量,最大叶面积采用裁剪质量法[9]测定,地上部干鲜质量采用万分之一天平进行称量。土壤速效氮含量、速效磷含量、速效钾含量、有机质含量、pH值分别采用碱解扩散法、钼锑抗比色法、火焰光度计法、重铬酸钾外加热法、水土比法(1 ∶5)[10]进行测定。根际土壤微生物数量采用稀释平板涂抹法[11]进行测定,其中细菌、真菌、放线菌分别采用牛肉膏蛋白胨培养基、马丁氏培养基、改良高氏一号培养基进行培养。脲酶、蔗糖酶、磷酸酶活性分别采用次氯酸钠比色法、对硝基苯磷酸盐法、磷酸苯二钠比色法[12]测定。
1.4 数据处理
用Excel 2007软件处理数据,用SPSS 23.0软件进行统计分析,用Duncan's新复极差法进行多重比较。
2 结果与分析
2.1 腐殖酸对盐胁迫下苦瓜幼苗形态建成的影响
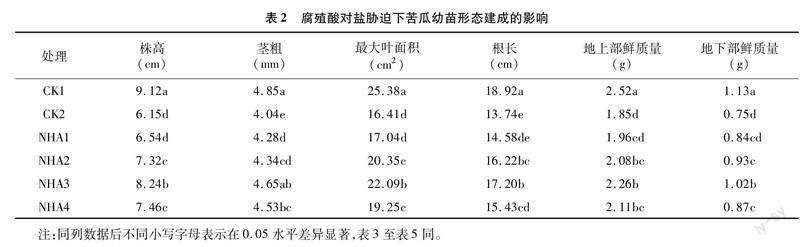
由表2可知,与CK1处理相比,CK2处理能够显著降低苦瓜幼苗株高、茎粗、最大叶面积、根长、地上部鲜质量、地下部鲜质量,与CK1处理相比分别降低了32.57%、16.70%、35.34%、27.38%、26.59%、33.63%,表明盐胁迫能够抑制苦瓜幼苗的生长,不利于苦瓜幼苗的形态建成和生物量积累。在NHA1、NHA2、NHA3、NHA4处理下,苦瓜幼苗株高、茎粗、最大叶面积、根长、地上部鲜质量、地下部鲜质量均高于单纯的盐胁迫处理(CK2处理),且以NHA3处理对应的值最大,与CK2处理相比分别增加了33.98%、15.10%、34.61%、25.18%、22.16%、36.00%,与CK2处理间差异均显著(P<0.05),说明腐殖酸的施用能够在一定程度上缓解盐胁迫对苦瓜幼苗生长的抑制作用。另外,在4个处理中,随着腐殖酸浓度的增加,苦瓜幼苗上述指标呈现先升高再降低的变化趋势,表明腐殖酸对苦瓜幼苗的生长存在一定的閾值,在阈值范围内表现为促进,反之表现为抑制。
2.2 腐殖酸对盐胁迫下苦瓜幼苗土壤化学性质的影响
由表3可知,与盐胁迫对照(CK2处理)相比,腐殖酸能够增加盐胁迫土壤的速效氮、速效磷、速效钾和有机质含量,且上述指标对腐殖酸用量的差异存在不同响应。随着腐殖酸用量的增加,苦瓜幼苗根际土壤速效氮含量呈现先升高再降低的单峰变化规律,在NHA2处理下最大,较CK2处理增加了23.67%,与CK2处理间差异显著(P<0.05),与CK1处理间差异不显著。速效磷、速效钾含量的变化规律与速效氮一致,但以NHA3处理最大,分别较CK2处理增加了46.55%、31.35%,与CK2处理间差异显著(P<0.05)。有机质含量则随腐殖酸用量的增加呈现逐渐升高的变化规律,在NHA4处理下最大,较CK2处理增加了21.64%,与CK2处理相比差异显著(P<0.05)。pH值随着腐殖酸用量的增加呈现逐渐降低的变化规律,在NHA4处理下最小,较CK2处理降低了12.97%,与CK2处理相比差异显著(P<0.05)。另外,施用腐殖酸的4个处理(NHA1、NHA2、NHA3、NHA4)的土壤速效氮、速效磷、速效钾和有机质含量虽高于单纯的盐胁迫对照(CK2处理),但均低于非盐胁迫的对照(CK1处理)。
2.3 腐殖酸对盐胁迫下苦瓜幼苗根际土壤微生物群落结构的影响
由表4可以看出,在15、30 d处理时间内,盐胁迫下的细菌、真菌数量均随腐殖酸浓度的增加,呈现先增加再降低的单峰变化规律。其中,在NHA1处理下,细菌、真菌数量虽高于CK2处理,但与其差异均不显著,说明低浓度的腐殖酸对盐胁迫环境下微生物数量的影响较小;在NHA3处理下,细菌、真菌数量均最大,分别较CK2处理增加了48.79%、64.89%;在NHA4处理下,腐殖酸浓度高于NHA3处理,但细菌、真菌数量却降低,表明过高浓度的腐殖酸并不能为微生物生长繁殖提供相对适宜的环境。在15、30 d处理时间内,盐胁迫下的放线菌数量随腐殖酸浓度的增加而呈现逐渐升高的变化趋势。其中,放线菌数量在NHA1处理下与CK2处理间的差异不显著,在NHA2、NHA3、NHA4处理下与CK2间的差异显著。另外,在同一处理时间内,腐殖酸虽然提高了盐胁迫下苦瓜幼苗根际土壤微生物数量,但仍均低于非盐胁迫(CK1处理)条件下的微生物数量,说明腐殖酸对盐胁迫下微生物数量的调控作用仍存在一定限度。同时,随着处理时间延长,不同处理条件下的细菌、真菌、放线菌数量均呈现不同程度的升高趋势。
2.4 腐殖酸对盐胁迫下苦瓜幼苗根际土壤酶活性的影响
由表5可知,随着处理时间的延长,土壤酶活性均表现出增加趋势,不同处理的脲酶、蔗糖酶、磷酸酶活性在试验30 d时均高于15 d时的酶活性;在不同处理时间内,各土壤酶活性的变化规律一致,即脲酶、蔗糖酶、磷酸酶活性在处理15 d时的变化规律分别与处理30 d时一致。在处理15、30 d时,CK2处理的脲酶、蔗糖酶、磷酸酶活性均显著低于对照CK1,表明盐胁迫能够抑制土壤酶活性的提高。在处理15、30 d时,添加腐殖酸的4个处理的脲酶活性均高于CK2处理,表明腐殖酸能提高盐胁迫土壤中脲酶的活性。其中,NHA1处理与CK2处理之间差异不显著,NHA2、NH3、NH4处理之间差异不显著,但均显著高于CK2处理。在处理15、30 d时,添加腐殖酸的4个处理的蔗糖酶活性均随腐殖酸浓度的增加呈现先升高再降低的单峰变化规律,且以NHA3处理最大,较CK2分别增加了41.86%、43.56%,与CK2处理间差异显著(P<0.05)。在处理15、30 d时,添加腐殖酸的4个处理的磷酸酶活性随腐殖酸浓度的增加呈现逐渐升高的变化趋势,在NHA4处理下最大,分别较CK2处理增加了26.17%、51.34%,与CK2处理差异显著(P<0.05)。
2.5 盐胁迫下苦瓜幼苗根际土壤微生物数量与土壤养分之间的相关性分析
由表6可以看出,速效氮含量与细菌、真菌、放线菌数量之间存在正相关关系,其中速效氮含量与细菌数量之间存在显著正相关关系;速效磷含量与细菌、真菌数量之间存在极显著正相关关系,与放线菌数量之间存在显著正相关关系;速效钾含量与细菌数量之间存在极显著正相关关系,与真菌、放线菌数量之间存在显著正相关关系;有机质含量与细菌数量之间存在显著正相关关系,与放线菌数量之间存在极显著正相关关系;pH值与细菌数量之间存在显著负相关关系,与放线菌数量之间存在极显著负相关关系,表明微生物数量与土壤养分含量之间存在正向交互作用,与土壤pH值之间存在负相关关系。
3 讨论与结论
盐胁迫是影响植物生长发育的主要非生物胁迫因子之一,轻度盐胁迫会引起植株生长速度下降,而重度盐胁迫会造成植株生长明显停滞甚至死亡[13]。汪晓丽研究得出,植物遭受盐害时,首先表现为老叶叶缘或叶尖变黄枯萎,然后是新叶出现类似症状[14]。姚静等研究指出,盐胁迫通过抑制地下部根系的长度、根吸收面积等指标来影响地上部的生长[15]。本试验结果表明,盐胁迫显著降低了苦瓜幼苗株高、茎粗、最大叶面积、根长、地上部鲜质量和地下部鲜质量,结果与郭茜茜等在辣椒[16]、黄洁等在粳稻[17]上得出的研究结论一致,这是因为盐胁迫会降低植株渗透势,造成离子稳态的紊乱及单盐毒害。在盐胁迫下,植物中的活性氧积累增加,生物质膜受到破坏,细胞受到损伤,光合性能下降,碳同化减少,进而影响植株的生长[18]。腐殖酸是一种结构复杂的天然高分子物质,其分子内含有多种活性官能团,能与许多有机物、无机物发生相互作用[19]。研究发现,腐殖酸被植物吸收后能够有效刺激植株生长,并增强抗盐碱、抗干旱等抗逆境胁迫能力[20]。在本试验条件下,施用腐殖酸提高了盐胁迫环境下的苦瓜幼苗长势,且以160 mg/L腐殖酸对苦瓜幼苗的促进作用最佳,这是因为腐殖酸含有较多活性基团,增加了其自身的盐基交换量,通过吸附和阻留土壤可溶性盐中的阳离子,降低土壤中盐离子浓度,减轻了盐胁迫伤害。
盐渍化土壤的黏性增强,水稳性团粒结构减少,养分转化进程受到抑制,对植物生长不利[21]。前人研究指出,腐殖酸在土壤中形成团粒结构、改良土壤状况、增加土壤肥力、提高土壤养分利用率、提高植株抗逆性等方面有重要作用,可有效改善土壤理化性质[22-23]。本试验结果表明,与单一盐胁迫相比,腐殖酸能够提高鹽胁迫土壤的速效氮、速效钾、速效磷含量,试验结果与叶协锋等在植烟上的研究结论[24]一致,这可能是因为腐殖酸是有机胶体物质,含有多种功能基团,阳离子交换量大,具有较强的吸附缓冲能力,可通过吸附、络合土壤中游离的氮磷钾元素,减少雨水的淋洗和土壤的固定,在一定程度上提高土壤大量元素速效养分含量[25-26]。另外,腐殖酸通过氨化反应,将吸附的NH-3、NH+4转化成腐殖酸铵盐,可以有效降低氮肥以氨态氮形式流失,提高土壤速效氮含量。磷作为植物生长所必需的大量元素,在土壤中不易被吸收。腐殖酸通过解磷作用,使阴离子与从土壤中吸附来的磷酸根离子发生同晶置换,将土壤固定的游离态磷释放出来,从而提高了土壤速效磷含量[27]。腐殖酸提高盐胁迫土壤有机质含量的试验结果与韩剑宏等在腐殖酸对盐碱土理化性质的影响方面的研究结论[28]一致,这是因为腐殖酸本身就是一种大分子有机质,且腐殖酸为微生物创造了更加适宜的生活环境,促进微生物对土壤有机物质的腐化和分解,增加有机质含量[29]。pH值对土壤理化性质、微生物群落结构有重要影响。在本试验条件下,随着腐殖酸含量的增加,土壤pH值呈下降趋势,试验结果与胡明芳等在棉花上的研究结论[30]类似,这可能由于腐殖酸是一种有机酸,它含有的羧基、羟基等酸性官能团释放出的活泼H+能够与土壤中的碱性物质发生化学反应而生成H2O,降低土壤pH值。另外,腐殖酸能够提高根系活力,刺激根系分泌更多的有机质和H+,这在一定程度上进一步降低了土壤pH值[31]。
土壤微生物参与矿质元素的转化、有机质的分解、腐殖质的合成与养分的循环等多种生物化学反应,同时还能有效杀灭有害病原菌、降解土壤中残留的有害物质,对改良土壤结构、提高土壤肥力有重要影响[32-33]。以往的研究结果表明,尽管不同微生物的耐盐性存在差异,但在盐胁迫环境下,其生长普遍受到抑制。在本试验条件下,在单纯盐胁迫(CK2处理)条件下,细菌、真菌、放线菌数量在处理15、30 d时均显著低于非盐胁迫处理(CK1处理),试验结果与任红等在刺槐上的研究结论[34]一致,这是因为盐胁迫改变了土壤的理化性质,降低了土壤通透性,不适宜微生物的生长繁殖。另外,盐胁迫使植株生理机能遭受不同程度的破坏,植株生长受到抑制,根系分泌的有益物质减少,为微生物提供的营养物质也随之减少,这也在一定程度上限制了微生物数量的增加[35]。微生物在疏松透气、养分含量丰富、理化性质良好的土壤中含量相对较高[36]。腐殖酸能够增加土壤孔隙度、降低土壤容重,同时可以向外界提供活性因子和营养元素,对微生物的生理活动有直接影响[37]。袁婉潼等研究得出,450 kg/hm2 腐殖酸用量能够显著提高盐碱土壤细菌、真菌、放线菌数量[38]。孙世君等研究得出,适量的腐殖酸能够显著提高盐胁迫条件下黄瓜根际土壤的真菌数量[39]。本试验结果表明,适宜浓度的腐殖酸处理(NHA1、NHA2、NHA3处理)能够促进细菌、真菌数量的增加。这可能由于在适宜的浓度下,腐殖酸以电子受体的形式参与微生物能量的合成,促进微生物生长[40]。腐殖酸表面活性大,易被细胞膜吸附,而腐殖酸络合、螯合、离子交换过来的营养元素在其被吸附后更有利于微生物的吸收利用,从而为微生物的生长繁殖提供更加充足的养分[41]。另外,腐殖酸还可以通过改善土壤微环境,调节土壤中气相、液相物质状况,为微生物生长繁殖提供适宜的生长环境[42-43]。在本试验条件下,放线菌数量在高浓度腐殖酸条件下仍呈增加趋势,这可能是因为放线菌适宜在中性或微碱环境中生长,高浓度的腐殖酸使盐胁迫土壤pH值降低了1.05,pH值的降低对放线菌的增殖作用超过渗透胁迫对放线菌的抑制作用,从而表现出高浓度腐殖酸条件下放线菌数量不降反升的趋势。另外,盐胁迫下苦瓜幼苗根际土壤微生物数量与土壤养分之间的相关性分析结果表明,提高盐胁迫环境下土壤养分有利于根际土壤微生物数量的增加,而微生物数量的增加又反过来促进矿质元素的转化、有机质的分解,不但能够提高土壤肥力,还可以改良土壤结构,有利于促进盐胁迫环境下植物的生长。
脲酶是直接参与尿素转化的酶,它水解尿素生成的氮是植物氮素营养的主要来源,同时与土壤中的氮循环密切相关。蔗糖酶在土壤中广泛存在,它可以催化土壤中蔗糖的分解,与土壤碳循环密切相关。磷酸酶是一类可以催化土壤有机磷化合物矿化的酶,参与土壤有机磷的分解与转化,与土壤磷循环密切相关。本试验结果表明,盐胁迫显著降低了苦瓜根际土壤脲酶、蔗糖酶、磷酸酶活性,这一方面是因为微生物和植物分泌物是土壤酶的重要来源,盐胁迫降低了微生物数量和植株生长势,减少了土壤酶的含量,另一方面是因为盐胁迫产生的离子毒害和渗透胁迫进一步抑制了土壤酶的活性[44]。腐殖酸能够提高盐胁迫环境下苦瓜根际土壤脲酶活性,且当腐殖酸浓度为160 mg/L时活性最大,这可能由于脲酶活性位点上半胱氨酸含有的巯基(—SH)易被氧化,常导致脲酶活性降低,而腐殖酸含有具有不饱和结构和较多的芳香基团,这些结构和基团能够在一定程度上防止脲酶巯基(—SH)的氧化,从而保护脲酶维持一定活性[45]。但也有研究指出,在一定条件下,腐殖酸可以抑制脲酶活性,这是因为腐殖酸与脲酶结合后,与脲酶底物形成竞争关系,致使脲酶活性因活性位点的减少而降低。但这种情况多发生在pH值低于6时,而在pH值达到7以上时,腐殖酸与脲酶均带负电荷,二者相互排斥,此时的腐殖酸对脲酶活性位点的影响较小,对其活性的抑制作用随之降低[46]。在本试验条件下,腐殖酸处理的苦瓜根际土壤pH值介于7~8之间,腐殖酸对脲酶活性位点的影响较小。腐殖酸提高苦瓜根际土壤蔗糖酶活性,试验结果与靳志丽等的研究结论[47]一致,这可能由于腐殖酸可以与蔗糖酶形成酶-腐殖物复合体,酶的结构变得更加稳定,使得活性进一步提高。另外,根系分泌物是蔗糖酶的重要来源,腐殖酸可以刺激根际分泌物增加,从而提高蔗糖酶数量和活力[48]。在盐胁迫条件下,随着腐殖酸浓度的增加,苦瓜根际土壤磷酸酶活性呈现逐步提高的变化趋势,与脲酶和蔗糖酶活性的变化规律存在差异,试验结果与刘兰兰等在生姜土壤微生物上的研究结论[49]一致,这是因为磷酸酶活性与土壤腐殖质含量呈正相关[50]。腐殖酸是腐殖质的主要成分,腐殖酸浓度的增加直接使土壤腐殖质含量提高,这使得苦瓜根际土壤磷酸酶活性随腐殖酸浓度的增加而提高。另外,腐殖酸能够刺激植株根系向土壤分泌多种有机物质,有机质被微生物分解成腐殖质,进一步刺激土壤磷酸酶活性提高[51]。
参考文献:
[1]文方芳. 种植年限对设施大棚土壤次生盐渍化与酸化的影响[J]. 中国土壤与肥料,2016(4):49-53.
[2]陈 昆,张正亮,高 磊.碱蓬内生菌对盐胁迫西瓜幼苗形态建成及生理特性的影响[J]. 江苏农业科学,2022,50(9):116-122.
[3]陈 昆,王红军,周银芝.碱蓬内生菌对西瓜幼苗盐胁迫的缓解效应[J]. 江苏农业科学,2022,50(19):135-142.
[4]操 庆,曹海生,魏晓兰,等. 盐胁迫对设施土壤微生物量碳氮和酶活性的影响[J]. 水土保持学报,2015,29(4):300-304.
[5]曾宪成. 腐殖酸从哪里来,到哪里去[J]. 腐植酸,2012(4):1-10,30.
[6]Zaremanseh H,Akbari N,Eisvand H R,et al. The effects of humic acid on soil physicochemical and biological properties under salinity stress conditions in pot culture of Satureja khuzistanica Jamzad[J]. Ecopersia,2020,8(3):147-154.
[7]高云晓,庞元湘,毛培利,等. 黄腐酸有机肥对盐胁迫下刺槐幼苗生长的影响[J]. 西南林业大学学报(自然科学版),2019,39(2):26-43.
[8]金 萍,陶 波,滕春红,等. 腐殖酸提高大豆抗盐碱能力的生理机制初探[J]. 东北农业大学学报,2006,37(2):229-231.
[9]熊庆娥. 植物生理学实验教程[M]. 成都:四川科学技术出版社,2008:55-62.
[10]鲍士旦. 土壤农化分析[M]. 3版.北京:中国农业出版社,2000:31-34,99-109,202-205.
[11]林先贵. 土壤微生物研究原理与方法[M]. 北京:高等教育出版社,2010:362-364.
[12]李阜棣,喻子牛,何绍江. 农业微生物学实验技术[M]. 北京:中国农业出版社,1996:69-89.
[13]Takemura T,Hanagata N,Sugihara K,et al. Physiological and biochemical responses to salt stress in the mangrove Bruguiera gymnorrhiza[J]. Aquatical Botany,2000,68(1):15-28.
[14]汪晓丽.大叶棒新品种‘恨天高对盐碱胁迫的生理生化响应[D]. 长沙:中南林业科技大学,2016:1-87.
[15]姚 静,施卫明. 盐胁迫对番茄根形态和幼苗生长的影响[J]. 土壤,2008,40(2):279-282.
[16]郭茜茜,吴 鹏,陈柏杰,等. 钠盐胁迫对不同辣度辣椒的耐受性影响研究初探[J]. 东北农业科学,2021,46(4):69-74,87.
[17]黄 洁,黄 晶,梁青铎,等. 盐胁迫对粳稻品种生长和生理特性的影响[J]. 中国稻米,2021,27(3):37-40.
[18]Xu Y J,van Huystee R B. Assciation of calcium and calmodulin to peroxidase secretion and activation[J]. Journal of Plant Physiology,1993,141(2):141-146.
[19]杨 敏,王红斌,宁 平,等. 云南沼泽土中提取腐殖酸的研究[J]. 化学世界,2002(7):351-353.
[20]梅慧生,杨玉明,张淑运,等. 腐殖酸钠对植物生长的刺激作用[J]. 植物生理学报,1980,6(2):133-140.
[21]王德领,诸葛玉平,杨全刚,等. 3种改良剂对滨海盐碱地土壤理化性状及玉米生长的影响[J]. 农业资源与环境学报,2021,38(1):20-27.
[22]曾宪成. 腐殖酸从哪里来,到哪里去[J]. 腐植酸,2012(4):1-10,30.
[23]李 丽,武丽萍,成绍鑫. 腐殖酸对磷肥增效作用的研究概况[J]. 腐植酸,1998(4):1-6.
[24]叶协锋,凌爱芬,刘清华,等. 腐殖酸对植烟土壤理化性状影响的研究[J]. 中国农业科技导报,2010,12(6):120-125.
[25]刘宇锋,罗 佳,苏天明,等. 外源腐殖酸对栽培基质性状和辣 椒生长发育的影响[J]. 江苏农业学报,2016,32(3):647-655.
[26]白志浩,雷治兵,张智敏,等. 腐殖酸復合肥料在农作物中的应用研究进展[J]. 山东化工,2020,49(19):56-58.
[27]周 爽,其力莫格,谭 钧,等. 腐殖酸提高土壤氮磷钾养分利用效率的机制[J]. 腐植酸,2015 (2):1-8.
[28]韩剑宏,孙一博,张连科,等. 生物炭与腐殖酸配施对盐碱土理化性质的影响[J]. 干旱地区农业研究,2020,38(6):121-127.
[29]孔令冉,董雯昕,杨天一,等. 腐殖酸在改良和培肥土壤中的作用[J]. 腐植酸,2019(1):7-12.
[30]胡明芳,田长彦,王 平,等. 黑液腐殖酸液体肥料对棉花生长及土壤理化性质的影响[J]. 西北农林科技大学学报(自然科学版),2013,41(12):195-199.
[31]蔡宪杰,杨 义,马 永. 腐殖酸类肥料对碱性植烟土壤 pH及烤烟产量质量的影响[J]. 中国农学通报,2008,24(6):261-265.
[32]郁 洁,蒋 益,徐春淼,等. 不同有机物及其堆肥与化肥配施对小麦生长及氮素吸收的影响[J]. 植物营养与肥料学报,2012,18(6):1293-1302.
[33]卜洪震,王丽宏,尤金成,等. 长期施肥管理对红壤稻田土壤微生物量碳和微生物多样性的影响[J]. 中国农业科学,2010,43(16):3340-3347.
[34]任 红,唐 琪,韩丛聪,等. 盐胁迫对刺槐根系离子、根际土壤酶及微生物种群的影响[J]. 山东农业科学,2018,50(2):38-44.
[35]徐 强,程智慧,孟焕文,等. 玉米-线辣椒套作系统中土壤养分与根际土壤微生物、酶活性的关系[J]. 应用生态学报,2007,18(12):2747-2754.
[36]邓 欣,谭济才,尹丽蓉,等. 不同茶园土壤微生物数量状况调查初报[J]. 茶叶通讯,2005,32(2):7-9.
[37]梁婉婉,高灵旺. 腐殖酸在植物保护领域的应用研究进展[J]. 腐植酸,2015(6):9-14,20.
[38]袁婉潼. 生物腐殖酸对盐碱土壤微生物数量和酶活性的影响[J]. 安徽农业科学,2016,44(1):1-4,45.
[39]孙世君,马 博,王逸轩,等. 腐殖酸对盐胁迫下温室黄瓜根际土壤养分及微生物群落结构的影响[J]. 北方园艺,2022(5):83-90.
[40]高 亮. 腐殖酸在酵素农业上的应用研究进展[J]. 腐植酸,2017(6):10-16.
[41]智 明,黄占斌,单瑞娟. 腐殖酸对土壤改良作用探讨[J]. 环境与可持续发展,2013,38(3):109-111.
[42]林先贵,王一明. 腐殖酸类物质是土壤健康的重要保障[J]. 腐植酸,2010(2):1-10.
[43]雷 菲,王 莉,刘海林,等. 腐殖酸缓释氮肥对糯玉米产量、氮肥利用率及土壤细菌多样性的影响[J]. 江苏农业科学,2022,50(17):271-275.
[44]刘善江,夏 雪,陈桂梅,等. 土壤酶的研究进展[J]. 中国农学通报,2011,25(1):197-201.
[45]张水勤,袁 亮,林治安,等. 腐殖酸促进植物生长的机理研究进展[J]. 植物营养与肥料学报,2017,23(4):1065-1076.
[46]许 辉. 土壤腐殖酸对脲酶活性和稳定性的影响[D]. 武汉:华中农业大学,2011:1-49.
[47]靳志丽,刘国顺,聂新柏. 腐殖酸对土壤环境和烤烟矿质吸收影响的研究[J]. 中国烟草科学,2002(3):15-18.
[48]周礼恺. 土壤酶学[M]. 北京:科学出版社,1987:205-207.
[49]刘兰兰,史春余,梁太波,等. 腐殖酸肥料对生姜土壤微生物量和酶活性的影响[J]. 生态学报,2009,29(11):6136-6141.
[50]刘 茜,马飞跃,于建军,等. 腐殖酸对植烟土壤和烟草影响的研究进展[J]. 中国农学通报,2010,26(4):132-136.
[51]彭正萍,门明新,薛世川,等. 腐殖酸复合肥对土壤养分转化和土壤酶活性的影响[J]. 河北农业大学学报,2005,28(4):1-4.
收稿日期:2023-04-16
基金項目:河南省高等职业学校青年骨干教师培养计划(编号:2019GZGG064);河南省农业职业学院科技创新人才支持计划(编号:HNACSRH-2021-03)。
作者简介:陈星星(1983—),女,河南南阳人,硕士,副教授,主要从事园艺作物生产栽培教学及科学研究。E-mail:starstar710@sina.com。
通信作者:王盛荣,硕士,高级农艺师,主要从事农业技术推广、园艺新品种开发利用及逆境生理栽培研究。E-mail:wangsr1975@163.com。
猜你喜欢
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
中小企业管理与科技·下旬刊(2016年11期)2016-11-18
人间(2016年27期)2016-11-11
科技视界(2016年9期)2016-04-26
科技视界(2016年7期)2016-04-01
