
荔枝成花、坐果与现“白点”期和末次秋梢期成熟期的关系研究
2023-10-21 18:41:59苏钻贤杨胜男黄悦万志远申济源陈厚彬
果树学报 2023年8期
苏钻贤 杨胜男 黄悦 万志远 申济源 陈厚彬

摘要:【目的】探讨妃子笑、怀枝、桂味和糯米糍的最适宜末次秋梢成熟期和现“白点”期,为荔枝控梢促花管理提供生物学指引。【方法】于2009—2021 年跟踪调查妃子笑、怀枝、桂味和糯米糍品种的关键物候期、成花枝率和坐果量,利用箱线图、小提琴图、T 检验、Pearson 相关等方法对数据的分布、差异显著性和相关性进行分析,并研究荔枝秋梢期、现“白点”期与成花和坐果之间的关系。【结果】4 个品种共获得522 组数据。桂味等4 个品种在10 月16—31 日末次秋梢成熟、1 月15 日前现“白点”的植株,成花枝率约90%,成熟期坐果量等级最高;在11 月1 日后末次秋梢成熟、1 月16 日后现“白点”的植株,成花枝率多下降到60%以下,且数据波动较大,坐果量等级下降为最低。坐果量与现“白点”期和成花枝率均存在极显著相关性,相关系数分别为-0.51 和0.54;成花枝率与现“白点”期以及末次秋梢成熟期均存在极显著负相关性,相关系数分别为-0.59 和-0.20。【结论】在广州地区,调控妃子笑、怀枝、桂味和糯米糍4 个品种末次秋梢在10 月16—31 日成熟,在1 月15 日前现“白点”,是在复杂农业气象条件下获得高成花和高产量的最可靠策略。
关键词:荔枝;末次秋梢成熟期;现“白点”期;成花枝率;坐果量
中图分类号:S667.1 文献标志码:A 文章编号:1009-9980(2023)08-1628-12
荔枝(Litchi chinensis Sonn.)是重要的亚热带常绿果树,原产于我国西南部,主产于南北纬17°~32°之间[1]。成花不稳定导致荔枝“大小年”结果现象突出,严重影响产业健康发展。影响荔枝成花诱导的因素很多,通常认为冬季低温[2-4]、干旱[5]、枝梢状态[6]等因素最为重要。重测序分析表明,荔枝一对COL基因中的一段3.7 kb 区域不同程度缺失,导致不同品种对低温的需求量和开花期差异[1]。不同品种之间对低温的需求存在共性規律,即日最高温20 ℃以下或日最低温10 ℃以下,低温持续时间需要6~8周。花穗发育期理想的天气特点是:昼间日均气温回升(可达20 ℃以上)、夜间冷凉(可低至10 ℃以下)。根据成花“阶段性假说”[7],荔枝在秋末枝梢生长成熟、在冬季低温期间经历花诱导,而后在冬末春初气温回升时完成花分化和开花。叶片转绿、顶芽处于停止生长状态的荔枝末端秋梢才能对冬季低温诱导作出响应[8]。顶芽出现花穗原基(即“白点”)是荔枝完成成花诱导的标志。随着全球气候变化加剧,秋冬季低温来临时间、强度和持续时间在年际间不确定性越来越大,而秋梢状态是荔枝栽培管理中可调控的因素之一,因此明确各地区、不同品种荔枝品种末次秋梢达到成熟状态和“白点”显现的适宜时间十分重要。
物候学是研究植物或动物周期性重复的生长发育模式的科学,从最初观察和记录少数几个物种的几个关键年度自然事件时间的经验学科发展到当今融合观测网络[9]、实验和建模[10]的综合科学。近年来,卫星遥感通过检测与绿色相关的植被指数[11]、叶绿素荧光[12]或高频采集图像[13]等手段拓宽了传统植物物候观测的视野,但这些方法仍存在空间分辨率和图像解释准确率低等缺陷,而地面观测获得物候期数据仍然是常绿果树物候研究最可靠的手段[14]。
数据分析与挖掘被应用于物联网、电子商务、金融、健康医疗、舆情分析、生物技术等各个领域。其中相关性分析目的在于探寻数据集里所隐藏的相关关系网[15],在机器学习、生物信息、医学、信息检索、社会统计学、经济学等众多领域和学科都有广泛应用。
笔者在本研究中对华南农业大学荔枝园的关键物候进行连续观测记载,结合成花坐果数据,分析成花率、坐果量、现“白点”期与末次秋梢成熟期之间的关系,探讨妃子笑、怀枝、桂味和糯米糍最适宜的末次秋梢成熟时间和现“白点”时间,挖掘荔枝物候期、成花与坐果的共性规律,为荔枝控梢促花管理提供生物学依据。
1 材料和方法
1.1 试验地点与材料
试验地点为广州五山华南农业大学校本部教学科研基地荔枝园(113.366° E,23.164° N)。基地分两个地块,地块一于2003 年种植,密度为6 m×5 m;地块二于2013年种植,密度为6.0 m×2.5 m。试验材料为妃子笑、怀枝、桂味和糯米糍4 个荔枝品种的圈枝苗。2009—2015 年,在地块一每个品种选择3 株作为试材;2016—2021 年,在地块二选择试材数量分别为:妃子笑10 株,怀枝20 株,桂味39株,糯米糍22 株。所选试材生长势较一致,株高约3.0 m,冠幅约2.5 m,采用了较高水平的土肥水和植保管理。
1.2 数据采集
2009—2021 年期间,连续观测记载每棵单株的末次秋梢成熟期、现“白点”期、成花枝率和坐果量。物候期的记载标准如下。
末次秋梢成熟期:参考Wei 等[16]的标准,75%以上末次秋梢的最后一片小叶完全转成绿色(图1-A)的日期。
现“白点”期:参考Wei 等[16]的标准,全株25%或以上的枝梢顶芽鳞片完全打开后,露出内部白色茸毛体(称为“白点”,图1-B)的日期。
成花枝率和坐果量调查方法如下。
成花枝率:全树成花枝数量与全树枝梢的百分比,在花穗生长期调查。
坐果量:生理落果高峰结束后进入成熟采收期的全树结果个数,在果实成熟期调查。共获得522 组数据,其中妃子笑81 组、怀枝116组、桂味202 组、糯米糍123 组。每组数据均包含每株树每年的成花枝率、坐果量和相应的物候期。
把荔枝植株分为高成花(成花枝率≥75%)、中等成花(25%≤成花枝率<75%)和低成花(成花枝率<25%)三个等级。分析不同成花枝率级别植株的末次梢成熟日期。将荔枝植株分为高坐果(坐果量≥第三四分位数,440 个)、中等坐果(中位数,130 个≤坐果量<第三四分位数,440 个)和低坐果(坐果量<中位数,130 个)三个等级,分析不同坐果级别植株的现“白点”日期。
在珠三角地区,11 月平均气温下降到20 ℃以下[17],而20 ℃是妃子笑等低需冷量荔枝品种的成花诱导性低温,此时末次秋梢需要处于成熟状态并保持一定时长,因此,10 月份是荔枝末次秋梢成熟的关键时期。为研究荔枝末次秋梢成熟早晚导致的成花和坐果差异,将此物候期分为三个时段:早于10月15 日,10 月16—31 日和迟于11 月1 日。1 月份历史平均气温约12 ℃,是全年气温从最低开始上升的月份[17],也是荔枝现“白点”的关键时期。为研究荔枝现“白点”早晚导致的成花和坐果差异,将此物候期分为早于1 月15 日、1 月16—31 日和迟于2 月1 日三个时段。
1.3 数据分析
将末次秋梢成熟期和现“白点”期转化为年积日期,以便于对不同年份的数据进行平均值计算、相关性分析以及差异显著性分析。
应用箱线图(Box plot)对调查数据进行异常值判断。作为一种统计图,箱线图可以提供数据分布、多样性、偏向正态分布以及异常值的直觀视图[18],它不需要事先假定数据服从特定分布形式,能真实地表现数据分布的原始面貌,被广泛应用于样本数据特征的可视化。箱线图的结构和关系如图2-A 所示。该图显示了最小值(Minimum)、下四分位数(Lower quartile,Q1)、中位数(Median)、平均值(Mean)、上四分位数(Upper quartile,Q3)和最大值(Maximum)以及介于Q1 与Q3 之间的箱形区(四分位数范围)。
最小值= Q1 - 1.5 × IQR; (1)
最大值= Q3 + 1.5 × IQR; (2)
四分位距(IQR:Inter-Quartile Range)= 上四分位数(Q3)- 下四分位数(Q1)。(3)
四分位距IQR 即箱形区域的高,与数据集的多样性成正比。由IQR标度的中位数和平均值之间的距离是评估正态分布偏差的量化指标。最小值到最大值之外的任何数据都被归类为异常值,这些数据与给定数据的特征存在显著差异。异常值通常重复性较低,故将其删除后再作进一步的统计分析,以便更可靠地描述[19]。
应用小提琴图[2(0] violin plot)查看数据的分布情况。小提琴图除可以展示样本数据的最小值、下四分位数、中位数、平均值、上四分位数、最大值以及四分位距外,还能呈现数据的概率密度分布曲线(图2-B)。
应用T-检验分析数据组之间是否存在差异。p≥0.05表示无显著差异(ns);*、**和***分别表示统计检验为p<0.05、0.01和0.001 时的差异显著水平。应用桑基图(Sankey)[21-22]查看荔枝植株数量从末次秋梢成熟、现“白点”、成花到坐果的变化。桑基图主要由边、节点和流量组成,流量与边的宽度成正比,边越宽说明数值越大。
用Excel 进行数据整理,使用R 语言(3.6.1 版,https://www.r-project.org)完成差异显著性分析、用Pearson 相关性分析和作图。
2 结果与分析
2.1 荔枝末次秋梢成熟期、现“白点”期、成花枝率与坐果量的主要特征
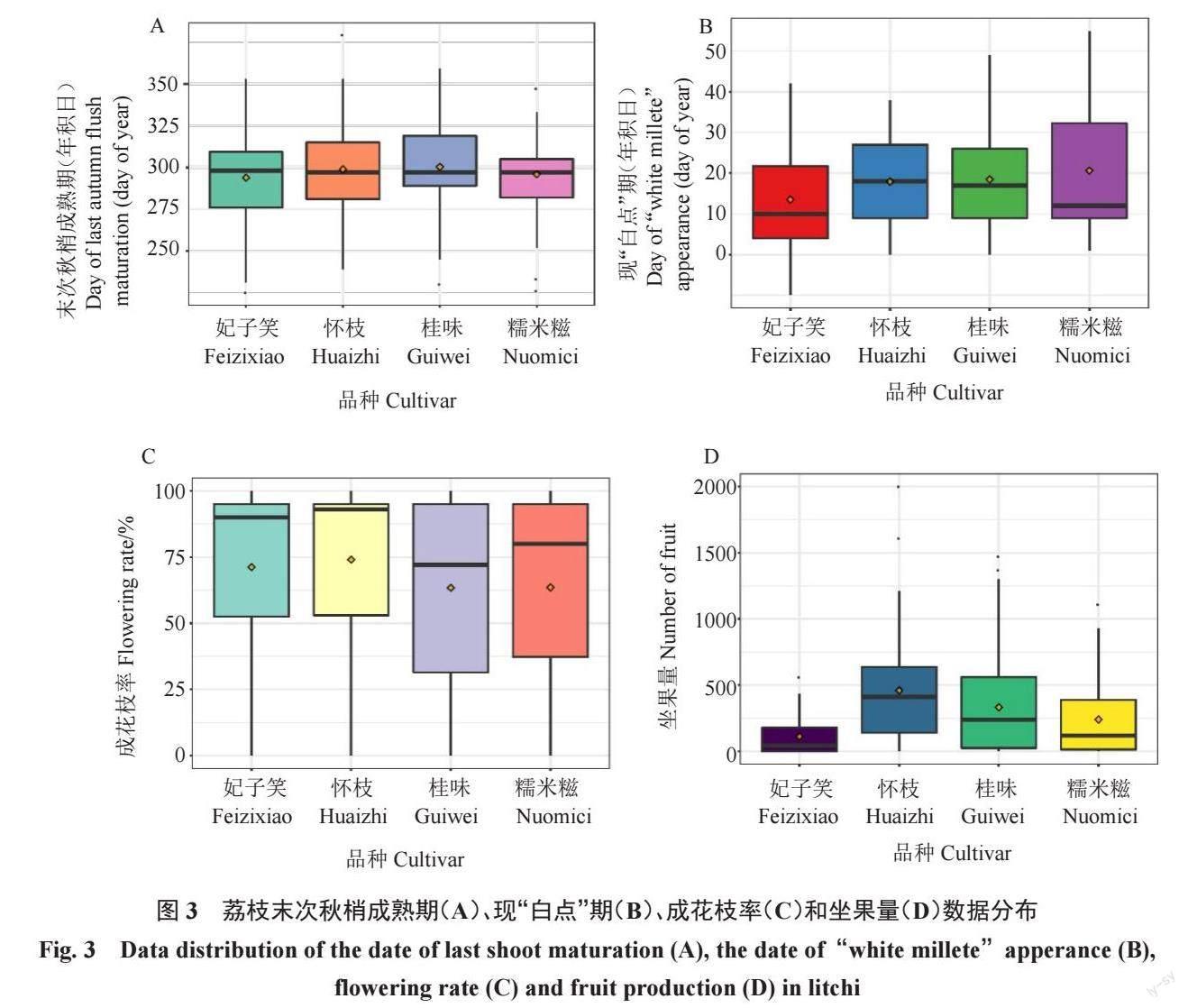
根据箱线图法原理,去除以下异常值:末次秋梢成熟期,妃子笑为225 d,怀枝379 d(即下一年第14天),桂味230 d,糯米糍226 d、233 d 和347 d(图3-A);现“白点”期和成花枝率未发现异常值(图3-B与图3-C);坐果量,妃子笑为559 个,怀枝为2000 个和1616 个,桂味为1470 个和1367 个,糯米糍1108 个(图3-D)。
去除异常值后将年积日转换为日期格式(2021年),妃子笑、怀枝、桂味、糯米糍4 个品种分别计算,末次秋梢成熟期平均值在10 月22—27 日间,中位数在10 月19—25 日间;现“白点”期平均值在1 月13—20 日间,中位数在1 月10—17 日间;成花枝率平均值57%~75%,中位数64%~90%;坐果量平均值93~372,中位数40~322。
2.2 末次秋梢成熟期对成花枝率和坐果量的影响
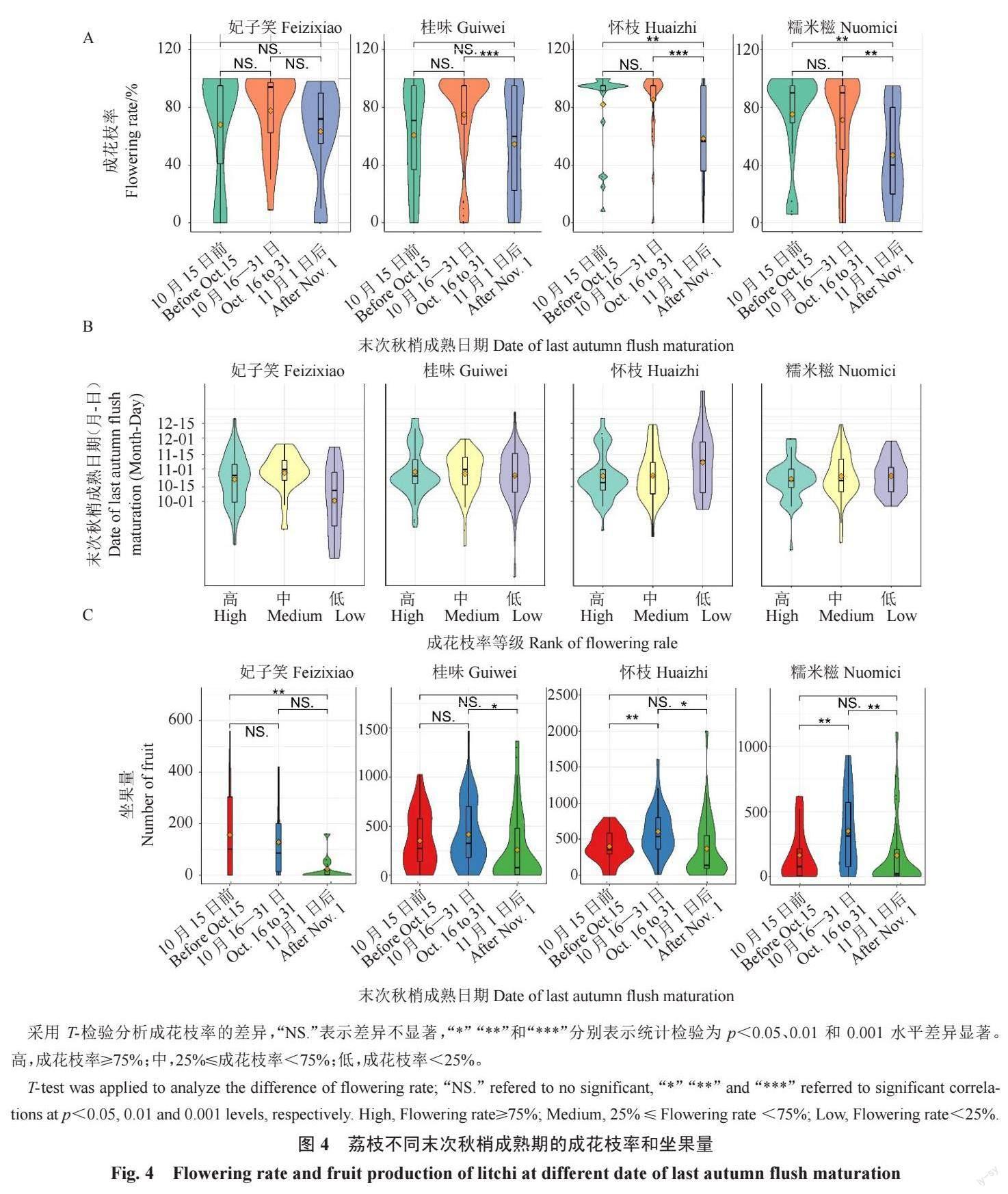
2.2.1 不同末次秋梢成熟期植株的成花枝率从图4-A可见,妃子笑在不同成熟期的成花枝率差异不显著,但在10 月15 日前或11 月1 日后成熟的植株,成花枝率低于20%的情况更多;11 月1 日前成熟的成花枝率高于80%的数据分布要比11 月1 日后的集中。怀枝在11 月1 日前成熟的平均值都大于80%,且集中分布在90%左右;11 月1 日后成熟的差异较大,平均值约50%。桂味在10 月16—31 日成熟时成花率平均值约80%,且集中分布在90%上下,显著高于11 月1 日后成熟的植株;11 月1 日后成熟的平均值低于50%,并且分散分布。糯米糍在10 月31 日前成熟的成花枝率都接近80%,且集中分布在90%上下,显著高于11月1 日后成熟的植株。
2.2.2 成花枝率与末次秋梢成熟期的关系妃子笑、怀枝、桂味和糯米糍高成花植株末次秋梢成熟时间主要分布在10 月16—31 日间。桂味中、低成花植株有较高比例早于10 月1 日前或晚于11 月5 日成熟。糯米糍中、低成花植株则有高比例早于10月1日前或晚于11月5日成熟,最晚接近12月份(图4-B)。从图4-C可见,妃子笑末次秋梢在10 月15 日前成熟的坐果量要显著高于11 月1 日以后的。怀枝和糯米糍在10 月16—31 日成熟的坐果量显著高于另外两时间段。桂味在11 月1 日后成熟的植株坐果量显著低于10月31日前成熟的。
2.3 现“白点”期对成花枝率和坐果量的影响
2.3.1 不同现“白点”期植株的成花枝率从图5-A可见,妃子笑的成花枝率,在1 月15 日前现“白点”的植株高于80%的情况多于其他时段。怀枝在1 月15日前现“白点”的植株成花枝率高于80%,显著高于2 月1 日后的;1 月16 日以后现“白点”的植株成花枝率差异较大。桂味和糯米糍在1 月15 日前现“白点”的植株平均成花枝率约80%,集中分布在90%左右,显著高于1 月16 日后现“白点”的植株;1 月16—31日现“白点”的植株成花枝率分布较为分散;2 月1 日后现“白点”的植株成花枝率多数低于40%。
2.3.2 坐果量与现“白点”期的关系如图5-B 所示,妃子笑高坐果量的植株较少,中等坐果量的植株现“白点”日期多在1 月10 日前或1 月20 日前后,低坐果量的植株现“白点”日期甚至达2 月10 日前后。怀枝高坐果量植株现“白点”日期主要集中在1 月10日前后,中低坐果量的植株现“白点”日期多数晚于1 月15 日,较集中在1 月30 日前后。桂味高坐果植株现“白点”日期主要在1月1—10日,中坐果量树现“白点”期主要集中在1 月20 日前后,而低坐果量植株现“白点”日期较分散,部分在1 月10日左右,也有部分推迟到2 月10 日左右。糯米糍高成花率植株“白点”日期主要集中在1月1—10日,中低坐果量的植株现“白点”日期较分散,从1 月初延续至2 月初。从图5-C可见,妃子笑在三个时间段的坐果量差异不显著,但2 月1 日后现“白点”的植株几乎无坐果。怀枝在2 月1 日后现“白点”的植株大部分坐果量都少于250 个。桂味和糯米糍在1 月15 日前现“白点”的植株坐果量都显著高于1 月16 日后现“白点”的植株;2 月1 日后现“白点”的多数植株几乎无坐果。
2.4 不同物候期荔枝植株的成花坐果表现
植株经历不同末次秋梢成熟期和现“白点”期后,其成花枝率和坐果量的走向如图6 所示。比较不同物候期可见,末次秋梢在10 月16—31 日间成熟的,在1 月15 日前现“白点”占61%,在1 月16—31 日现“白点”占34%,在2 月1 日后现“白点”占5%;有62%为高成花枝率,26%和12%为中、低成花枝率;有31%为高坐果量,有31%和37%为中、低坐果量。1 月15 日前现“白点”的植株,高、中、低成花枝率分别为67%、15%和18%;高、中、低坐果量分别为37%、19%和44%。10 月16—31 日前成熟、1 月15 日前现“白点”的植株,成花枝率和坐果量都最高。从植株成花坐果表现看,高成花枝率植株组中,末次秋梢在10 月16—31 日成熟的植株比例最高,为45%;1 月15 日前现“白点”的植株占65%,比例最高。高坐果量植株组中,末次秋梢在10 月16—31 日成熟的植株比例最高,为47%;1 月15 日前现“白点”的植株占74%,比例最高。
2.5 荔枝成花、坐果与末次秋梢成熟期和现“白点”期的相关性
如图7 所示,荔枝坐果量与成花枝率以及现“白点”期均存在极显著相关性,相关系数分别为0.54和-0.51;成花枝率与现“白点”期以及末次秋梢成熟期均存在极显著负相关性,相关系数分别为-0.59和-0.20。虽然坐果量与末次秋梢成熟期无直接相关,但它与成花枝率呈极显著相关,而坐果量与成花枝率呈极显著正相关。查看末次秋梢成熟期与坐果量的置信椭圆区,及与成花枝率以及坐果量相关性的置信椭圆区,现“白点”期在8~30 d(为1 月8—30日);末次秋梢成熟期在270~325 d(为9 月27 日—11 月27 日)。与坐果量相关性的置信椭圆区,成花枝率为35%~100%。
3 讨论
荔枝的生长阶段主要分为芽、枝、叶发育,花序出现、开花、果实发育和果实成熟等[16]。由于荔枝花芽分化具有階段性[7],精准控制荔枝末次秋梢成熟时间、培养早熟末次秋梢以及早促发“白点”是荔枝丰产的重要保障。有学者提出广西地区桂味末次秋梢在11 月下旬成熟成花率最高[23-24],但试验只设置了一年重复。陈厚彬等[25]建议粤中地区妃子笑于11月5 日前成熟、1 月5 日前现“白点”,桂味和糯米糍于10 月15 日前成熟、1 月25 日前现“白点”;在粤西地区上述日期应提前5~10 d,但未涉及成花率和坐果量的分析。植株末次秋梢期和现“白点”期控制在哪个时间段成花和坐果的可靠性最高,一直未有明确的答案。本研究中通过对妃子笑、怀枝、桂味和糯米糍4 个荔枝品种持续12 年的关键物候期、成花和坐果数据进行统计分析,挖掘不同品种的共性规律,结果发现,成花枝率、坐果量与末次秋梢成熟期和现“白点”期存在极显著相关性。对关键物候期分阶段比较其开花与坐果情况,发现4 个品种在广州地区10 月15—31 日末次秋梢成熟、1 月15 日前现“白点”的植株,成花枝率和坐果量都最高,成花枝率在75%以上,且集中分布在90%左右;在11 月1 日后末次秋梢成熟、1 月15 日后现“白点”的植株,成花枝率下降到60%以下,且数据波动较大,坐果量也显著下降。
荔枝末次秋梢成熟时,叶片通过光合作用大量积累碳水化合物(如淀粉)储存于枝梢和枝干中[26-27],以供来年开花坐果消费利用;未成熟的叶片光合能力不足,而新萌发的叶芽更是强大的库,而且这些状态的叶片都不能有效感受低温诱导[28]。在诱导性低温[3]来临时,如末次秋梢成熟期过迟,会导致枝叶生长量和碳水化合物储存不足[6],叶片中GA和IAA等抑制成花激素偏高[29],FLC 等成花抑制基因表达水平高[30],使叶片不能接受成花诱导,从而错过诱导性低温;如末次秋梢成熟期过早,则有抽发迟梢甚至冬梢的倾向。因此,通过营养和水分管理,调整末次秋梢充足的生长量和适宜的成熟时间窗口是荔枝有效接受成花诱导的前提。海南省荔枝产区在9 月下旬即开始对妃子笑进行控梢,使其末次秋梢保持成熟状态等候短暂的低温窗口期。广西产区在11 月下旬开始对桂味和糯米糍控梢。本研究中,广州地区的妃子笑在10月15 日前成熟的枝梢,之后可不同程度长出新梢,如不加严格控制,可能影响成花。
在完成花芽诱导后,适时促发“白点”,可使荔枝花芽随着早春和缓的升温过程完成形态分化,形成优质花穗。生产上较常出现“白点”期延迟的情况,对荔枝成花坐果不利:一是1 月中下旬之后现花穗原基的植株,花穗生长时易遭遇高温而成花逆转[26],导致不能成花或成花率下降;二是现“白点”后花穗发育时间缩短,形成短花穗或弱花穗;三是晚花荔枝多在清明前后开放,较大概率遭遇持续阴雨,不利于授粉受精。末次秋梢成熟期延迟[31]和土壤干旱[32]均导致“白点”期推迟;适当的末次秋梢成熟期、降水和足量灌溉[26,33]、施用生长调节剂[2]则有利于“白点”及时出现。
虽然“适宜秋梢成熟期”和“适宜现‘白点期”的植株成花和坐果都处于较高等级,但即便在最适宜物候期下,仍存在一定比例的低成花率和低坐果量情况。这可能是不适宜的天气条件造成的:(1)在秋梢成熟期,未有足够强度的持续低温,造成成花诱导不充分,花芽质量低;(2)现“白点”后,持续高温干旱会使荔枝出现成花逆转,降低成花枝率;(3)开花期持续低温阴雨会影响授粉受精,果实发育期的干旱或阴雨都容易加剧落果,影响最终坐果量。尽管如此,在复杂多变的天气条件下,通过培养健壮的结果母枝,提高树体的碳素积累量,使荔枝处于最适宜物候期,仍有最大概率获得高产。
农业气象变化复杂,难以精准预测,在多变的天气下采取最可靠的管理策略对荔枝获得高产量意义重大。本研究目的是在复杂多变的气象环境下,把同一地区正常管理的品种作为整体,从多年数据中探寻普适性规律。已有研究表明,2018—2019 年间广州地区妃子笑、怀枝、桂味和糯米糍4 个品种成花诱导期间,气温变化可分成三个阶段,阶段一多处于20~30 ℃之间;阶段二从20 ℃下降至10 ℃以下,随后缓慢上升;阶段三在20 ℃上下。年际间气温下降和上升出现的时间和强度有差异,但变化趋势一致性较高。在复杂的气候变化中,上述4 个品种的成花诱导持续期平均值没有显著差异[31]。
经过综合分析,笔者在本研究中明确提出在广州或珠三角地区,通过农艺措施,使妃子笑、怀枝、桂味和糯米糍4 个品种在10 月16—31 日末次秋梢成熟、1 月15 日前现“白点”,有较大可能提高成花率和获得高的产量。这与海南省荔枝产区在9 月下旬控制妃子笑末次秋梢成熟的策略一致,均是使荔枝植株保持最佳的树体状态进入相应的时间窗口。调控的农艺措施有结合环割、化学药物精准控梢、及时灌溉、喷施叶面肥促发“白点”等,在此基础上分析荔枝成花与坐果最适宜物候期的树体碳素、矿质元素和激素水平,制定秋梢的营养标准,实现精准调控。
4 结论
对2009—2021 年4 个荔枝品种522 组物候期和开花坐果数据的分布、差异显著性分析和相关性分析,结果表明,荔枝成花枝率与末次秋梢成熟期和现“白点”期存在极显著负相关性,坐果量和成花枝率与现“白点”期存在极显著相关性。明确在广州地区,调控妃子笑、怀枝、桂味和糯米糍4 个品种末次秋梢在10 月16—31 日成熟,在1 月15 日前现“白点”,是在复杂的农业气象条件下获得高成花率和高产量的最可靠策略。
参考文献References:
[1] HU G B,FENG J T,XIANG X,WANG J B,SALOJ?RVI J,LIU C M,WU Z X,ZHANG J S,LIANG X M,JIANG Z D,LIU W,OU L X,LI J W,FAN G Y,MAI Y X,CHEN C J,ZHANG X T,ZHENG J K,ZHANG Y Q,PENG H X,YAO LX,WAI C M,LUO X P,FU J X,TANG H B,LAN T Y,LAI B,SUN J H,WEI Y Z,LI H L,CHEN J Z,HUANG X M,YAN Q,LIU X,MCHALE L K,ROLLING W,GUYOT R,SANKOFFD,ZHENG C F,ALBERT V A,MING R,CHEN H B,XIA R,LI J G. Two divergent haplotypes from a highly heterozygous lycheegenome suggest independent domestication events for earlyand late-maturing cultivars[J]. Nature Genetics,2022,54(1):73-83.
[2] MENZEL C M. The control of floral initiation in lychee:A review[J]. Scientia Horticulturae,1983,21(3):201-215.
[3] CHEN H B,HUANG H B. Low temperature requirements forfloral induction in lychee[J]. Acta Horticulturae,2005(665):195-202.
[4] 陳厚彬,苏钻贤,张荣,张红娜,丁峰,周碧燕. 荔枝花芽分化研究进展[J]. 中国农业科学,2014,47(9):1774-1783.
CHEN Houbin,SU Zuanxian,ZHANG Rong,ZHANG Hongna,DING Feng,ZHOU Biyan. Progresses in research of Litchi floraldifferentiation[J]. Scientia Agricultura Sinica,2014,47(9):1774-1783.
[5] SHEN J Y,XIAO Q S,QIU H J,CHEN C J,CHEN H B. Integrativeeffect of drought and low temperature on litchi (Litchichinensis Sonn.) floral initiation revealed by dynamic genomewidetranscriptome analysis[J]. Scientific Reports,2016,6:32005.
[6] ZHENG Q,DAVENPORT T L,LI Y C. Stem age,winter temperatureand flowering of lychee in South Florida[J]. Acta Horticulturae,2001(558):237-240.
[7] 黃辉白,陈厚彬. 以阶段观剖视荔枝的花芽分化[J]. 果树学报,2003,20(6):487-492.
HUANG Huibai,CHEN Houbin. A phasic approach towards thefloral formation in Litchi chinensis Sonn.[J]. Journal of Fruit Science,2003,20(6):487-492.
[8] ROBBERTSE H,FIVAZ J,MENZEL C. A reevaluation of treemodel,inflorescence morphology,and sex ratio in lychee (Litchichinensis Sonn.) [J]. Journal of the American Society for HorticulturalScience,1995,120(6):914-920.
[9] SCHWARTZ M D. Phenology:An integrative environmental science[M]. Dordrecht:Springer,Netherlands,2013.
[10] DE BARBA D,ROSSI S,DESLAURIERS A,MORIN H. Effectsof soil warming and nitrogen foliar applications on budburst of black spruce[J]. Trees,2016,30(1):87-97.
[11] PIAO S L,FANG J Y,ZHOU L M,CIAIS P,ZHU B A. Variationsin satellite-derived phenology in Chinas temperate vegetation[J]. Global Change Biology,2006,12(4):672-685.
[12] SMITH W K,BIEDERMAN J A,SCOTT R L,MOORE D J P,HE M,KIMBALL J S,YAN D,HUDSON A,BARNES M L,MACBEAN N,FOX A M,LITVAK M E. Chlorophyll fluorescencebetter captures seasonal and interannual gross primary productivitydynamics across dryland ecosystems of southwesternNorth America[J]. Geophysical Research Letters,2018,45(2):748-757.
[13] SEYEDNASROLLAH B,YOUNG A M,HUFKENS K,MILLIMANT,FRIEDL M A,FROLKING S,RICHARDSON A D.Tracking vegetation phenology across diverse biomes using Version2.0 of the PhenoCam Dataset[J]. Scientific Data,2019,6:222.
[14] AONO Y,KAZUI K. Phenological data series of cherry treeflowering in Kyoto,Japan,and its application to reconstructionof springtime temperatures since the 9th century[J]. InternationalJournal of Climatology,2008,28(7):905-914.
[15] 李国杰,程学旗. 大数据研究:未来科技及经济社会发展的重大战略领域—大数据的研究现状与科学思考[J]. 中国科学院院刊,2012,27(6):647-657.
LI Guojie,CHENG Xueqi. Research status and scientific thinkingof big data[J]. Bulletin of Chinese Academy of Sciences,2012,27(6):647-657.
[16] WEI Y Z,ZHANG H N,LI W C,XIE J H,WANG Y C,LIU LQ,SHI S Y. Phenological growth stages of lychee (Litchi chinensisSonn.) using the extended BBCH-scale[J]. Scientia Horticulturae,2013,161:273-277.
[17] 封静,潘安定,李冰. 珠三角地区近40 年的气温变化特征分析[J]. 热带农业工程,2011,35(5):60-64.
FENG Jing,PAN Anding,LI Bing. Air temperature variation inrecent 40 years over Pearl River Delta region[J]. Tropical AgriculturalEngineering,2011,35(5):60-64.
[18] SPITZER M,WILDENHAIN J,RAPPSILBER J,TYERS M.BoxPlotR:A web tool for generation of box plots[J]. NatureMethods,2014,11(2):121-122.
[19] 郭德清,廖祥文. 基于箱線图的微博客热点话题发现[J]. 山西大学学报(自然科学版),2014,37(1):19-25.
GUO Deqing,LIAO Xiangwen. Microblog hot topic detectionbased on box-plot[J]. Journal of Shanxi University (Natural ScienceEdition),2014,37(1):19-25.
[20] HINTZE J L,NELSON R D. Violin plots:A box plot- densitytrace synergism[J]. The American Statistician,1998,52(2):181-184.
[21] LUPTON R C,ALLWOOD J M. Hybrid Sankey diagrams:Visualanalysis of multidimensional data for understanding resourceuse[J]. Resources,Conservation and Recycling,2017,124:141-151.
[22] 周忠凯,赵继龙,林佳潞,刘长安. 桑基图的可视化图式在建成环境领域的应用[J]. 山东建筑大学学报,2017,32(6):536-544.
ZHOU Zhongkai,ZHAO Jilong,LIN Jialu,LIU Changan. Applicationof Sankey diagram as a visualization graph in the builtenvironment[J]. Journal of Shandong Jianzhu University,2017,32(6):536-544.
[23] 侯延杰,李鸿莉,邱宏业,李冬波,徐宁,秦献泉,张树伟,朱建华,彭宏祥. 桂味荔枝2 种不同时期老熟秋梢营养变化研究[J]. 南方园艺,2021,32(6):11-14.
HOU Yanjie,LI Hongli,QIU Hongye,LI Dongbo,XU Ning,QIN Xianquan,ZHANG Shuwei,ZHU Jianhua,PENG Hongxiang.Study on nutritional changes of autumn shoots of Guiweilitchi matured at two different stages[J]. Southern Horticulture,2021,32(6):11-14.
[24] 李平,钟敏芝,覃燕,冯晓佳,潘介春,邓英毅,徐炯志,程夕冉,莫云川.‘桂味荔枝成花调控技术研究[J]. 中国南方果树,2018,47(6):65-68.
LI Ping,ZHONG Minzhi,QIN Yan,FENG Xiaojia,PAN Jiechun,DENG Yingyi,XU Jiongzhi,CHENG Xiran,MO Yunchuan.Study on flower- forming regulation of‘Gui Weilitchi[J].South China Fruits,2018,47(6):65-68.
[25] 陈厚彬,苏钻贤,陈浩磊. 荔枝“大小年”结果现象及秋冬季关键技术对策建议[J]. 中国热带农业,2020(5):10-16.
CHEN Houbin,SU Zuanxian,CHEN Haolei. The biennial bearingin Litchi and key management in autumn and winter[J]. ChinaTropical Agriculture,2020(5):10-16.
[26] MENZEL C M,RASMUSSEN T S,SIMPSON D R. Effects oftemperature and leaf water stress on growth and flowering of litchi(Litchi chinensis Sonn.)[J]. Journal of Horticultural Science,1989,64(6):739-752.
[27] MENZEL C M, SIMPSON D R. Temperatures above 20 ℃ reduceflowering in lychee (Litchi chinensis Sonn.)[J]. Journal ofHorticultural Science,1995,70(6):981-987.
[28] SU Z X,XIAO Q S,SHEN J Y,CHEN H B,YAN S J,HUANGW J. Metabolomics analysis of Litchi leaves during floral inductionreveals metabolic improvement by stem girdling[J]. Molecules,2021,26(13):4048.
[29] 梁武元,梁立峰,季作梁,李沛文. 荔枝花芽分化过程中内源赤霉素和吲哚乙酸的含量动态[J]. 园艺学报,1987,14(3):145-152.
LIANG Wuyuan,LIANG Lifeng,JI Zuoliang,LI Peiwen. Thefluctuation of fndogenous gibberellin and indole-3-acetic acid inLitchi chinensis shoot-tips during floral initiation[J]. Acta HorticulturaeSinica,1987,14(3):145-152.
[30] YANG H F,KIM H J,CHEN H B,RAHMAN J,LU X Y,ZHOU B Y. Carbohydrate accumulation and flowering- relatedgene expression levels at different developmental stages of terminalshoots in Litchi chinensis[J]. HortScience,2014,49(11):1381-1391.
[31] SU Z X,LIU L Y,LI Y Q,CHEN H B. Predicting flower inductionof Litchi (Litchi chinensis Sonn.) with machine learningtechniques[J]. Computers and Electronics in Agriculture,2023,205:107572.
[32] CHAIKIATTIYOS S,MENZEL C M,RASMUSSEN T S. Floralinduction in tropical fruit trees:Effects of temperature andwater supply[J]. Journal of Horticultural Science,1994,69(3):397-415.
[33] MENZEL C M,SIMPSON D R. Effects of temperature and leafwater stress on panicle and flower development of Litchi (Litchichinensis Sonn.) [J]. Journal of Horticultural Science,1991,66(3):335-344.
猜你喜欢
发明与创新·小学生(2023年11期)2023-11-02 16:04:31
科学大众(2022年23期)2023-01-30 07:03:24
东方企业家(2022年9期)2022-11-25 15:37:05
岭南音乐(2022年4期)2022-09-15 14:03:10
花火·慧阅读(2022年5期)2022-05-21 11:56:58
锻压装备与制造技术(2021年6期)2022-01-17 04:22:36
快乐作文(5.6年级)(2021年11期)2021-12-17 02:26:33
中国(俄文)(2019年8期)2019-08-24 09:34:38
意林·全彩Color(2019年4期)2019-05-11 09:06:20
岭南音乐(2017年2期)2017-05-17 11:42:07