
干旱对甘蔗及其近缘植物蔗糖代谢的影响
2023-10-19 05:20张倩倩龚茂健周鸿凯莫俊杰
江苏农业科学 2023年18期
杨 善, 方 欣, 张倩倩, 龚茂健, 周鸿凯, 莫俊杰
(广东海洋大学滨海农业学院,广东湛江 524088)
甘蔗(SaccharumofficinarumL.)作为最主要的制糖类经济作物之一,支撑了糖料产业发展。广东湛江凭借其独特的地理位置及水热条件,成为我国三大糖蔗产地之一,但湛江地区降雨量年际间变化大,年内季节分配不均,所以季节性干旱导致甘蔗的产量和品质大幅下降,影响甘蔗产业健康发展[1]。干旱是影响植物生长和发育的最主要因素之一,可使甘蔗平均产量减少31.7%[2]。在甘蔗生育期中,伸长期对水分的需求量最大,如果该时期遇到干旱,对甘蔗最终产量影响最大[3]。因此,研究干旱胁迫下甘蔗伸长期蔗糖代谢的变化特征,对今后培育抗旱甘蔗品种具有重要意义。
为应对干旱胁迫,植物会产生一系列适应性机制,如生长减缓、光合作用降低、气孔关闭、渗透调节物质合成积累增多、相关激素含量升高、抗逆基因上调表达等[4]。在干旱胁迫下,甘蔗正常生理代谢过程会遭到破坏,长时间干旱胁迫会导致甘蔗生长减缓、植株矮小,产量和品质显著下降[5]。有研究表明,干旱使得甘蔗叶片的细胞膜透性增大、丙二醛(MDA)含量增加、膜脂过氧化作用加剧、叶绿素含量下降、脯氨酸含量迅速增加,以提高甘蔗体内渗透势,利于根系从土壤中吸收水分[6-7]。吴凯朝等以甘蔗品种GT11为材料,研究认为,甘蔗为响应水分胁迫,除可溶性蛋白含量和ProDH活性外,叶片MDA、可溶性糖、超氧阴离子、脯氨酸、脱落酸(ABA)含量及过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、谷胱甘肽还原酶(GSH-R)、Δ′-吡咯啉-5-羧酸合成酶(P5CS)和多胺氧化酶(PAO)活性均明显高于对照处理[8]。由此表明,干旱胁迫下耐旱品种具有更强的调控能力,以提高甘蔗对干旱胁迫的适应性。
蔗糖是植物中碳水化合物运输的主要形式,从叶片的叶绿素中合成磷酸丙糖,运输到细胞质中参与蔗糖合成,也可用于氨基酸和脂肪酸的合成。在作物糖代谢生理过程中,蔗糖磷酸合成酶(SPS)、蔗糖合成酶(SS)、酸性转化酶(AI)、中性转化酶(NI)起着重要的调节作用[9]。SPS以果糖(6-磷酸F6P)和尿苷二磷酸葡糖(UDPG)为底物催化合成蔗糖,是一种不可逆的酶[10]。SS既能催化果糖和UDPG合成蔗糖,又能催化蔗糖分解,是一种可逆酶[11]。转化酶在蔗糖代谢中不可逆地催化蔗糖分解为果糖和葡萄糖,分为中性和酸性转化酶[12]。在自然生长条件下,节间蔗糖含量与酸性转化酶活性呈极显著负相关,与蔗糖磷酸合成酶活性呈显著正相关,表明酸性转化酶、蔗糖磷酸合成酶对甘蔗节间蔗糖含量有显著性调节作用[13]。叶燕萍等研究认为,干旱处理提高了甘蔗蔗糖糖分和蔗汁重力纯度而降低了蔗汁还原糖分,因而提高了甘蔗品质[14]。由此表明,甘蔗成熟期适当的干旱胁迫有助于提高蔗糖糖分,提高甘蔗品质。
前人研究侧重于在干旱胁迫下研究甘蔗生理生化变化特性,或利用干旱胁迫的生理指标对甘蔗材料进行抗旱性综合评价[15-17]。伸长期甘蔗的蔗糖积累及其代谢相关酶活性应答干旱胁迫的生理机理尚不清楚。本研究通过在甘蔗伸长初期,连续干旱胁迫0、2、4、6、8、10 d,分别测定甘蔗及近缘植物割手密、斑茅体内蔗糖代谢相关酶活性变化,以及可溶性糖、蔗糖含量变化,并对各测定指标进行相关性分析,探究不同甘蔗品种及近缘植物应对干旱胁迫蔗糖代谢的响应特性,为今后深入研究干旱胁迫下甘蔗糖代谢、培育抗旱高糖甘蔗品种提供理论参考。
1 材料与方法
1.1 试验材料
选用2个甘蔗栽培品种,分别是Badila(干旱敏感品种)、ROC22(耐旱,推广面积最大主栽品种),甘蔗近缘植物为湛江地区的10倍体割手密(x=8,2n=80,Saccharumspontaneum)和6倍体斑茅(x=10,2n=60,Erianthusarundinaceus),以上材料均由广东省科学院南繁种业研究所湛江研究中心提供。
1.2 试验方法
1.2.1 试验设计 采用桶栽(胶桶高50 cm、口径40 cm,桶底均匀开4个排水小孔)方式进行种植,所用土壤为砖红壤土,每桶装土25 kg,与20 g复合肥(N、P2O5、K2O含量均为17%)混合均匀后装桶,每桶种植双芽苗4段,每桶出苗分蘖后定植6株。每个品种种植3桶,共12桶。
试验地点在广东海洋大学农业生物技术研究所。2020年6月7日将种苗种植入土,入土深度为10 cm,浇水使土壤保持湿润。2020年6月20日出苗,出苗后每2 d浇500 mL水1次。干旱胁迫前,在研究所大院空地培育,所有材料种植管理水平一致。出苗60 d后,甘蔗处于伸长期初期(11~13张真叶),将全部桶搬至通风透气良好的玻璃温室内,进行干旱胁迫试验,不再进行浇水。搬至玻璃温室后,次日上午8时至9时取样,每次每桶剪取1株甘蔗的+1叶,迅速放入液氮中速冻,带回实验室放入-80 ℃冰箱冻存,用于相关指标测试分析。之后,每2 d(48 h)取样1次,共取样6次,分别为干旱胁迫后0、2、4、6、8、10 d。
1.2.2 测定项目及方法 用烘干法测定土壤含水量[18]。细胞膜透性的测定采用电导法,可溶性糖含量的测定采用蒽酮比色法,蔗糖含量以及蔗糖磷酸合成酶、蔗糖合成酶、酸性转化酶、中性转化酶活性测定均参照高俊凤的方法[19]。
1.3 数据统计分析
应用统计分析软件SPSS 19.0分析试验数据,用Duncan’s新复极差法分析同一品种不同干旱胁迫时间点测定指标的差异显著性(α=0.05)。
2 结果与分析
2.1 土壤含水量
由图1知,随着干旱处理天数的增加,各个品种的土壤含水量均成一定梯度且显著性地下降,表明本研究的干旱胁迫梯度设计具有典型性和可靠性,可以在此干旱胁迫水平下进行各项指标的测定和研究。

2.2 甘蔗植株细胞膜透性分析
由图2可知,干旱胁迫下,与0 d相比,4种材料细胞膜透性增加的最大倍数分别为9.19倍、9.47倍、3.75倍、13.76倍,表明土壤水分含量是影响各材料细胞膜透性增加的主要因素。随着干旱处理天数的增加,ROC22、斑茅的细胞膜透性呈一定梯度增加,且斑茅的各梯度间有显著性差异,ROC22除30 d和2 d之间无显著性差异,其他处理梯度间均有显著性差异。Badila和割手密的细胞膜透性在干旱胁迫后8 d达到最大,之后开始减少,其他材料则一直增加。说明不同品种的耐旱能力不尽相同。

2.3 干旱胁迫下甘蔗植株糖代谢的生理效应分析
2.3.1 蔗糖磷酸合成酶活性 由图3可知,干旱胁迫下,各材料的SPS活性变化趋势一致,先增后减,在胁迫6 d时达到最大值,然后显著下降,可能是干旱胁迫8 d达到了各材料SPS调控的极限,从而表现出下降的趋势。然而,干旱胁迫下,抗旱品种ROC22的平均SPS活性低于其他材料的SPS活性。

2.3.2 蔗糖合成酶活性 由图4可知,6次测定值中,各供试材料SS活性均呈现先增后减的趋势,ROC22在干旱胁迫6 d时SS活性达到最大值,其余3个材料均在干旱胁迫4 d时SS活性达到最大值。另外,Badila与斑茅的SS活性变化非常相似,均比其他材料小,说明Badila与斑茅的SS活性对干旱胁迫的响应没有ROC22和割手密积极。
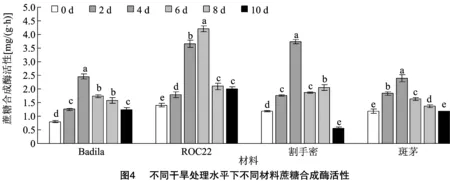
2.3.3 酸性转化酶活性 由图5可知,干旱胁迫下,各供试材料的AI活性在胁迫2 d后开始显著下降,Badila和割手密的AI活性在胁迫4 d时达到最低水平。ROC22和斑茅的AI活性在胁迫10 d时达到最低水平。胁迫6 d后Badila和ROC22的AI活性趋于稳定,没有显著性差异,而割手密在胁迫10 d后AI活性显著增加。在胁迫10 d时AI活性显著下降。值得注意的是,在干旱胁迫0 d和2 d时,斑茅的AI活性显著高于其他材料。
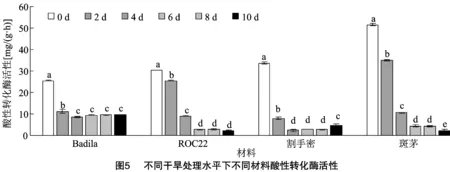
2.3.4 中性转化酶活性 从图6可知,干旱胁迫下,各个供试材料NI活性基本都呈现不断减小的趋势,但Badila胁迫4 d时先显著性增加,尔后减小,说明干旱胁迫会使得供试材料NI活性减小,以影响其体内蔗糖代谢的生理生化反应。6次测定值中,斑茅的平均NI活性最高,Badila次之,并且干旱胁迫下Badila的NI活性变化较其他材料稳定。

2.3.5 可溶性糖含量 由图7可知,除ROC22在胁迫6、8 d时可溶性糖含量下降,斑茅在胁迫6 d后可溶性糖含量基本不变外,干旱胁迫下各材料可溶性糖含量从胁迫2 d开始呈不断增长趋势,且有显著性差异。由此说明,供试材料可依靠大量积累可溶性糖来缓解水分胁迫,增加细胞内渗透势,以便根系从土壤中吸收水分。从整体观察各材料的可溶性糖含量变化,同样发现Badila、ROC22与割手密的变化趋势基本一致。然而,斑茅与其他材料不同,虽然也是上升趋势,但是干旱胁迫下斑茅的可溶性糖含量增加幅度并不大,其应对干旱胁迫的生理机制可能与甘蔗材料存在较大差异。

2.3.6 蔗糖含量 由图8可知,随干旱胁迫天数增加,Badila和割手密的蔗糖含量整体呈不断上升趋势,均在干旱胁迫10 d时蔗糖含量最高,ROC22和斑茅的蔗糖含量则呈先上升后下降的趋势,分别在胁迫后6、4 d达到最高。由此说明,在干旱胁迫下,大量积累蔗糖是各材料启动干旱胁迫的响应机制之一。从整体上观察,甘蔗栽培种的变化趋势与割手密的趋势基本一致,而不同于斑茅。
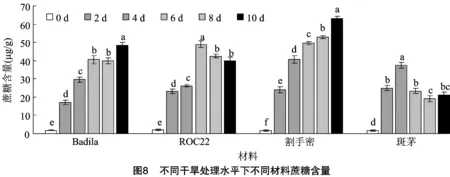
2.4 干旱胁迫下甘蔗及其近缘植物生理指标间的偏相关分析
由表1可知,通过偏相关分析,土壤含水量除了与Badila和割手密的可溶性糖含量、与斑茅的细胞膜透性和NI活性没有显著性相关外,与各生理指标均显著或极显著相关。细胞膜透性,在Badila中与SPS活性、NI活性、蔗糖含量存在极显著负相关关系;在ROC22中只与可溶性糖含量不存在显著性相关;在割手密中与SS活性、蔗糖含量存在显著或极显著性正相关;在斑茅中只与NI活性存在极显著性正相关。由此表明,干旱胁迫下甘蔗及其近缘植物的细胞膜透性与蔗糖含量、蔗糖代谢相关酶活性存在显著性相关,但不同材料间存在一定差异。干旱胁迫下,在Badila、ROC22、割手密中蔗糖含量与SPS、SS、AI活性存在极显著性正相关,在Badila中与NI活性呈显著正相关,在ROC22、割手密中与AI活性、NI活性呈显著负相关,而在斑茅中蔗糖含量与SS活性、AI活性分别存在极显著性正相关、负相关。可溶性糖含量,在Badila中只与AI活性存在极显著性相关,在ROC22中只与SPS活性、NI活性分别存在显著性正相关、负相关关系,在割手密、斑茅中分别只与SS活性、NI活性不存显著性相关。在4种材料中,只有在割手密中可溶性糖含量与蔗糖含量存在极显著性相关,说明干旱胁迫下,甘蔗的可溶性糖含量与蔗糖含量并不存在密切相关。

表1 不同材料生理指标间的偏相关分析
3 讨论与结论
水分短缺和季节性干旱严重影响作物的生长、品质和产量,因此,干旱胁迫已成为严重阻碍农作物可持续发展的重要因素之一[20]。对干旱胁迫,作物会产生感知、信号转导、渗透调节、转录调控、蛋白质修饰和代谢物质调控等应答反应[21-22]。为缓解干旱胁迫带来的不利影响,甘蔗体内会合成大量脯氨酸、甜菜碱、可溶性糖等渗透调节物质[23-24]。本研究中,随着干旱天数的增加,各个材料体内可溶性糖含量、蔗糖含量整体呈一定梯度显著性增加,以提高渗透压,这与前人的研究结果[25-26]一致。在4个材料中,干旱胁迫下,虽然斑茅可溶性糖含量、蔗糖含量整体为升高趋势,但其整体含量较其他2个甘蔗品种和割手密的低。究其原因,可能是由于斑茅与其他材料不在同一个属,遗传背景相差较大[27]。从偏相关分析可知,4个材料的土壤含水量与蔗糖含量均达到了极显著性负相关。然而,只在斑茅中土壤含水量与可溶性糖含量的关系为极显著负相关,在ROC22中为显著性负相关,由此说明,在甘蔗中蔗糖积极应答干旱胁迫,为主要的渗透调节糖类。同时,也进一步证明了干旱处理可提高甘蔗蔗糖糖分和蔗汁重力纯度,从而提高甘蔗品质[14]。所以,在大田生产中,可以对甘蔗进行适当的干旱处理,以便提高甘蔗蔗糖糖分。
蔗糖合成代谢过程主要受SPS、SS以及转化酶的影响[13]。本研究偏相关分析发现,Badila、ROC22、割手密中蔗糖含量与SPS、SS活性均为极显著正相关,与AI、NI活性为极显著负相关,但Badila中蔗糖含量与NI活性为极显著正相关。由此表明,甘蔗蔗糖含量与SPS、SS、AI、NI的活性密切相关,蔗糖合成代谢受4种酶的共同调控。前人研究表明,蔗糖磷酸合成酶可能是蔗糖合成途径中的一个重要控制点,它的活性反映蔗糖生物合成途径的能力,SPS活性越高,蔗糖积累得越多[28]。本研究发现,随着干旱天数增加,4个材料的SPS活性响应迅速,其活性在6 d达到最高值,之后呈一定梯度下降,并且在偏相关性分析中,SPS活性与蔗糖含量均呈极显著正相关,由此表明,甘蔗中SPS活性显著性地调控蔗糖合成。相关研究发现,在植物生长过程中,SS既可催化蔗糖合成又可催化蔗糖分解,但主要起分解作用[11]。赵丽宏等通过在甘蔗自然生长条件下,对蔗糖合成分解代谢进行研究认为,SS活性与蔗糖积累呈负相关[29]。这与本研究结果存在差异,随着干旱天数增加,SS活性显著增加,并在胁迫后4 d或6 d时活性达到最强,之后SS活性下降,并且在偏相关性分析中,各供试材料SS活性与蔗糖含量均呈极显著正相关。由此认为,干旱胁迫下,SS在甘蔗中主要起催化蔗糖合成的作用。转化酶分为AI和NI,在植物中主要催化蔗糖分解成果糖和葡萄糖,为植株的生长和发育提供碳源和能量,以保证植株的正常生理代谢活动[30-31]。本研究结果表明,随干旱天数增加,AI、NI活性呈一定梯度显著性下降,而蔗糖积累量则呈一定梯度显著升高,与蔗糖含量为显著负相关;而且,NI平均活性比AI平均活性低。有研究认为,在甘蔗生长过程中AI活性对蔗糖积累调控作用非常重要,此外,逆境条件下,AI活性对维持细胞的正常功能也起一定的作用[9]。由此说明,在甘蔗干旱胁迫中AI的调控作用更强。以上分析发现,SPS和SS活性受干旱胁迫激活,而转化酶活性受抑制。由此表明,干旱胁迫对甘蔗的蔗糖分解代谢活动产生了抑制作用,为了抵御干旱带来的渗透胁迫,甘蔗减少蔗糖分解以积累更多的蔗糖含量调节细胞渗透压。
干旱胁迫使得甘蔗及其近缘植物的细胞膜透性增大,可溶性糖和蔗糖含量上升,SPS和SS活性先升后降,AI和NI活性下降。偏相关分析表明,蔗糖含量与SPS、SS活性呈极显著正相关,与AI、NI活性呈极显著负相关。由此表明,甘蔗及其近缘植物蔗糖代谢积极响应干旱胁迫,主要以蔗糖合成为主,蔗糖分解为辅,缓解渗透胁迫。
猜你喜欢
天津医科大学学报(2021年1期)2021-12-05
广西糖业(2020年3期)2020-09-25
延边大学学报(自然科学版)(2019年2期)2019-10-08
浙江工业大学学报(2017年5期)2018-01-22
中国医学影像技术(2017年11期)2017-01-16
中国糖料(2016年1期)2016-12-01
考试周刊(2016年63期)2016-08-15
广西林业科学(2016年2期)2016-03-20
生物加工过程(2015年2期)2015-11-11
中国果业信息(2015年11期)2015-01-23
