
沉默SOCS1 对肝细胞癌患者树突状细胞表面抗原及其免疫调控的影响
2023-10-17 07:10董晓微张俊华樊瑞军
宁夏医科大学学报 2023年8期
董晓微, 郭 东, 柳 斌, 张俊华, 樊瑞军
(宁夏回族自治区人民医院临床医学检验诊断中心,银川 750002)
肝细胞癌占原发性肝癌的95%,居我国恶性肿瘤发病率第3 位,在全球范围内,每年可造成近100 万人死亡[1]。切除、移植、化疗栓塞、乙醇注射和冷冻消融术是治疗肝细胞癌的潜在方法,但仅对小且具有局限性的肿瘤有效[2]。不幸的是,大多数肝细胞癌患者在诊断时已到晚期,目前治疗效果不佳。因此,迫切需要为肝细胞癌研究新的治疗策略。近年来,在癌症治疗中,生物免疫治疗作用越来越突出,抗原提呈细胞之一的树突状细胞(dendritic cell,DC)不仅参与T 细胞的激活、免疫功能的恢复,并可促进相关细胞因子的释放等过程,从而发挥抗肿瘤效果[3]。细胞因子信号抑制因子1(suppressor of cytokine signaling 1,SOCS1)作为细胞因子信号抑制因子(SOCS)家族的一员,近年来得到广泛关注。SOCS1 不仅参与细胞信号传导,而且参与泛素化介导的蛋白质降解过程,与DC 分化、T 细胞功能以及肿瘤进展息息相关,这些功能在肿瘤细胞的生长和增殖中都起着重要作用[4]。微小RNA-618(micro RNA-618,miR-618)能够调节DC 以及T 细胞,作为抗癌免疫治疗的新方法,miR-618 在肝癌中也可能发挥着重要的抑癌因子作用。因此,miR-618 作为SOCS1的靶向miRNA,探究其转录调控和相关功能对DC 表面抗原及其免疫调控的影响,对于指导肝细胞癌临床免疫治疗意义重大。
1 材料与方法
1.1 材料
采集宁夏回族自治区人民医院2020 年10月至2021 年10 月收治的60 例肝细胞癌患者的外周静脉血,其中男性38 例,女性22 例;年龄52~75 岁,平均年龄(64.45±4.22)岁。通过淋巴细胞分离液离心得到外周血单核细胞(peripheral blood mononuclear cells,PBMC)。PBS 缓冲液、Lipofectamine3000 试剂盒、RPMI 1640 培养基和Gibco Opti-MEM 培养基均购自赛默飞世尔科技(中国) 有限公司;TransScriptⅡGreen One-Step RT-qPCR、SuperMix 试剂盒、TRIzol 试剂盒、RIPA 裂解液、BCA 试剂盒、白细胞介素(interleukin,IL)12、干扰素-α(interferon-α,IFN-α)和肿瘤坏死因子-γ(tumor necrosis factor-γ,TNF-γ)、ELISA试剂盒均购自上海碧云天生物技术有限公司;HEK-293T 细胞来自国家中心细胞库,所有序列均购自生工生物工程(上海)股份有限公司;所有抗体均购自美国Abcam 公司;pmirGLO 质粒、质粒提取试剂盒均购自上海源叶生物科技有限公司;FC 500 MCL 型五色流式细胞仪、7500 型全自动PCR 仪及UniCel DxI 800 型全自动化学发光免疫分析仪均购自美国Beckman 公司。
1.2 方法
1.2.1 DC 孵育 在RPMI 1640 全培养基中培养PBMC,第2 天去原培养液,加入等量含有IL-4和粒细胞-巨噬细胞集落刺激因子(granulocytemacrophage colony-stimulating factor,GM-CSF)与肝细胞癌细胞裂解液的新培养液继续孵育约7 d,在其分化成未成熟的DC 时去原培养液,更换等量IFN-α 的培养液,孵育至第9 天时获得成熟的DC。
1.2.2 T 细胞孵育 使用含10%牛血清的培养液将细胞PBMC 配成计数为4×107个/mL。使用37 ℃培养液洗涤尼龙毛柱,以约6 滴/min 放出Hanks 液,在尼龙毛柱中加入0.5 mL 的PBMC 悬液后,马上加0.2 mL 培养液后封闭塑料管,放入CO2培养箱中使PBMC 充分黏附尼龙毛柱,约培养60 min。接着用5 mL 培养液洗脱尼龙毛柱并收集,2 500 r·min-1离心8 min,去上清,继续常规孵育。
1.2.3 分组与转染 根据Lipofectamine3000说明书,将DC 转染和细胞培养后进行分组:仅含mimic 脂质体阴性对照组(mimic NC 组)、仅含miR-618 mimic 脂质体组(miR-618 mimic 组)、仅含SOCS1 shRNA 质粒脂质体组(sh-SOCS1 组)、仅含shRNA NC 质粒脂质体对照组(shRNA-NC组)、含SOCS1 shRNA 与miR-618 mimic 脂质体组(miR-618 mimic+sh-SOCS1 组)。
1.3 检测方法
1.3.1 相关细胞表面标记物水平的检测 首先将培养基重悬细胞配成计数为2×106个/mL。接着在流式细胞管中分别加入50 μL 的CD3 抗原提呈细胞(CD3-APC)、CD4 异硫氰酸荧光素(CD4 fluorescein isothiocyanate,CD4-FITC) 或CD8 藻红蛋白(CD8 phycoerythrin,CD8-PE)以及相应的50 μL 细胞,混匀后冰浴20 min。在4 ℃下2 500 r·min-1离心8 min,去上清,加入500 μL PBS 吹洗,重复2 次。最后加入400 μL PBS,使用FC 500 MCL 型五色流式细胞仪分析结果。
1.3.2 沉默SOCS1 由生工生物工程(上海)股份有限公司合成野生型SOCS1 及突变型3’UTR,随后将其克隆到含荧光素酶报告基因pmirGLO上游,提纯备用。按Lipofectamine3000 说明书将对照序列miR-618 mimic 和阴性转染至HEK-293T 细胞,孵育48 h 后收获细胞。检测细胞的相对荧光素酶活性(萤火虫荧光素酶值/海肾荧光素酶值)。
1.3.3 miR-618 表达量 用TRIzol 试剂盒提取DC 中总RNA。用焦碳酸二乙酯(diethyl pyrocarbonate,DEPC)溶解RNA,使用7500 型全自动荧光定量PCR 仪测量260 nm 和280 nm 下吸光值,测定总RNA 含量。使用RT-qPCR SuperMix试剂盒及PCR 仪扩增测定(上游引物:5’-GGGGAAACTCTA-CTTGTCCTT-3’,下游引物:5’-TCGTATCCAGTGC-GTGTCGT-3’)。依据试剂盒说明书完成反应体系与反应条件,miR-618表达水平用2-ΔΔCt法分析。
1.3.4 SOCS1 蛋白表达 将DC 培养基弃液,PBS 洗涤2 次,加入含1 mmol·L-1苯甲基磺酰氟的RIPA 裂解液,在冰上孵育5 min。随后4 ℃下10 000 r·min-1离心4 min。使用SDS-PAGE 凝胶电泳在聚偏二氟乙烯(polyvinylidene difluoride,PVDF)膜上测定蛋白浓度。室温下,在摇床上使用含5% 牛血清蛋白的Tris 盐缓冲液+TBST 缓冲液封闭1 h。弃液,以GAPDH 为内参,加入鼠一抗SOCS1,转膜后4 ℃冰箱中放置12 h。随后用TBST 冲洗,加入山羊抗鼠IgG 二抗,在4 ℃培养箱中孵育约5 h。PVDF 与化学发光液反应60 s,曝光后,采用Image J 软件分析蛋白相对表达量。SOCS1 蛋白质的相对表达量=目标带灰度值/内参照带灰度值。
1.3.5 DC表面抗原检测 将含DC的培养液与CD4-APC、CD11c-FITC、CD86-PE 分别加入Falcon管,2 500 r·min-1离心15 s,标记DC 后避光孵育15 min,随后添加100 μL 生理盐水,2 500 r·min-1离心90 s,将沉淀细胞转移至流式管,在流式细胞仪中检测,记录结果。
1.3.6 DC 上清液中细胞因子水平 4 000 r·min-1离心15 min 后收集DC 上清液,采用酶联免疫吸附试验法,按照对应的细胞因子试剂盒说明书操作,将其标准品梯度稀释后,制作标准曲线,调零,标准品为X 轴,吸光度为Y 轴,绘制标准曲线,计算测定IL-12、IFN-α 和TNF-γ 水平。
1.4 统计学方法
采用SPSS 22.0 统计学软件进行数据分析,计量资料以均数±标准差(±s)表示,多组间比较采用方差分析,两两比较采用Dunnett-t 检验。P≤0.05 为差异有统计学意义。
2 结果
2.1 光镜下观察DC
经裂解液处理后,DC 呈现聚集状态,多为圆形,失去树突状外观,细胞膜光滑且清晰,少数为星形,周围存在长短不等的突起,仍呈现梭形,见图1。

图1 光镜下观察DC(×40)
2.2 DC 表面抗原分析
采用流式细胞术检测DC 表面抗原情况。其中含CD3+的细胞约占94%,提示分离DC 表面抗原的T 细胞纯度较高。进一步分析,CD3+CD4+细胞约占36%,而CD3+CD8+细胞约占52%。CD1a+和CD83+分别为DC 特异性和成熟的细胞标记物,CD1a+细胞约占93%,CD1a+CD83+细胞约占85%。
2.3 DC 中miR-618 表达情况比较
在DC 诱导第0、1、3、5、7 天后进行miR-618表达水平比较,结果显示,诱导0、1、3 d 时miR-618 表达水平无明显变化,但经多种抗原刺激后,在第5、7 天miR-618 表达水平均比第0 天降低(P 均<0.05),见图2。

图2 DC 中miR-618 表达情况
2.4 miR-618 靶向沉默SOCS1 表达
双荧光素酶系统表明,miR-618 与SOCS1 表达存在靶向调控(P<0.05),见图3A。miR-618 过表达后,SOCS1 蛋白表达低于mimic NC 组(P<0.05),见图3B、图3C。
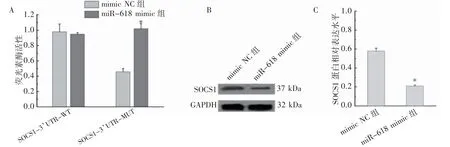
图3 miR-618 靶向沉默SOCS1 表达情况
2.5 沉默SOCS1 促进DC 活化
成熟DC 表面抗原CD4、CD11c、CD86、HLADR 的表达水平的结果表明,与各对照组相比,miR-618 mimic 组和shRNA-SOCS1 组CD11c、HLA-DR 和CD86 的阳性细胞均增多(P 均<0.05),CD4 阳性细胞无明显变化(P>0.05)。miR-618 mimic+sh-SOCS1 组CD11c、HLA-DR、CD86的阳性细胞数均增多(P 均<0.05),见图4。

图4 沉默SOCS1 促进DC 表面抗原表达
2.6 沉默SOCS1 促进DC 中细胞因子分泌
与各对照组相比,miR-618 mimic 组和shRNASOCS1 组TNF-γ、IL-12 和IFN-α 表达水平均上升(P 均<0.05),miR-618 mimic+sh-SOCS1 组上述因子均增加(P 均<0.05),见图5。
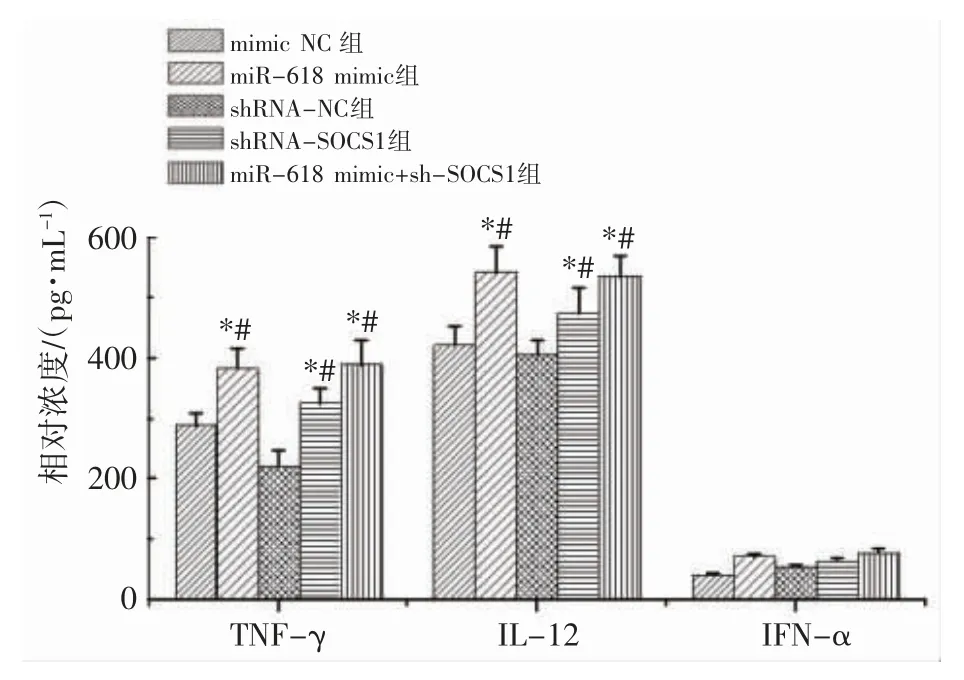
图5 沉默SOCS1 促进DC 中细胞因子分泌
3 讨论
SOCS1 是SOCS 家族的原型分子,作为负反馈调节的一部分,通过直接抑制酪氨酸激酶和激活的细胞因子受体的信号级联来下调细胞因子信号,减弱细胞因子启动的信号转导[5-6],SOCS1还通过直接和间接机制下调Toll 样受体(Tolllike receptor,TLR)信号[7],多种人类癌症中检测到肿瘤细胞中SOCS1 的异常表达,这与细胞因子受体和TLR 信号的失调有关。对肿瘤细胞中SOCS1 功能的研究揭示了其在致癌中的重要作用[8]。DC 是最有效的抗原呈递细胞,是调节和维持对肿瘤细胞免疫应答的核心[9]。作为抗原的来源,全蛋白、肿瘤细胞裂解物(TCL)、HLA 限制性肽、RNA 或杂交细胞可以通过DC 呈递给T 淋巴细胞[10]。有研究[11-12]阐述了TL 脉冲自体DC 免疫治疗黑色素瘤、肾细胞癌、儿童实体瘤和卵巢癌。miRNA 参与了DC 的分化,与肿瘤的免疫调控有关,将miRNA 作为靶点进而增强DC 的免疫功能。本研究结果显示,miR-618 在DC 诱导期间表达下调。
细胞因子与细胞表面的特定受体结合,将生物信息传递给靶细胞,与之相关的信号通路控制和调节基本的生物学过程,包括造血、炎症、免疫和肿瘤的发展[13-14]。SOCS1 可能在调节肿瘤生长和增殖中发挥作用[15]。对SOCS1 缺陷小鼠的研究证实了SOCS1 在调节CD8+T 细胞稳态中的关键作用[16]。在胸腺中,SOCS1 可防止阳性选择失败的胸腺细胞存活和扩张,确保阴性选择,并防止其向CD8 谱系的不当发育倾斜。SOCS1 不仅能控制T 细胞刺激性细胞因子的产生,还能降低CD8+T 细胞对协同细胞因子刺激和抗原非特异性激活的敏感性。由于CD8+T 淋巴细胞的细胞因子刺激增加了其对低亲和力T 细胞受体(T cell receptor,TCR)的敏感性,SOCS1 可能通过抑制由炎性细胞因子驱动的异常T 细胞活化来促进外周T 细胞耐受[17]。Rossato 等[18]发现,系统性硬化浆细胞样DC 中miR-618 的表达较健康对照组低,其可能是对免疫系统稳态进行调节的重要靶点。本研究证实,SOCS1 功能的发挥受靶向miRNA 的调控,而沉默SOCS1 在肝细胞癌免疫调节中具有重要价值。SOCS1 还可通过调节DC 的产生、成熟、抗原呈递、共刺激信号和细胞因子的产生来调节其对T 细胞的激活作用[19]。SOCS1 对T 细胞、T 调节细胞和DC 的上述调控机制可共同协助免疫耐受并预防自身免疫发生。另外,在DC或CD8+T 细胞中沉默SOCS1 能有效地刺激抗肿瘤免疫。SOCS1 还可通过调节致癌信号转导途径在癌细胞中起抑癌作用[20],它可以被细胞信号因子调节,SOCS1的启动子包含STAT 结合区,STAT 结合区域可以被STAT 信号因子调节[20]。促红细胞生成素(EPO)刺激下调EPO 应答细胞系中GFI1B 的表达,从而激活SOCS1。SOCS1 也可由其他细胞因子诱导,如IL-6、IL-4、IL-12、IL-13、TNF-γ、IFN-α/β 等。事实上,细胞因子介导的信号丢失可能导致效应器功能受损和耐受诱导。其他几种细胞因子包括IL-2、IL-4、IL-6、IL-15、IL-17 和IL-21 也有助于细胞增殖、分化、获得不同的效应器功能、防止衰竭和记忆细胞的产生[21]。本研究也发现,过表达miR-618 或沉默SOCS1 能够促进DC 中TNF-γ、IL-6 和IFN-α 等细胞因子的表达。
综上所述,沉默SOCS1 可以促进肝细胞癌患者DC 活化,提高其免疫调控能力。
猜你喜欢
九江学院学报(自然科学版)(2022年2期)2022-07-02
昆明医科大学学报(2022年3期)2022-04-19
中学生物学(2021年8期)2021-11-02
中国果业信息(2019年1期)2019-01-05
中国畜牧兽医文摘(2018年6期)2018-07-28
中成药(2018年2期)2018-05-09
中成药(2018年2期)2018-05-09
中成药(2017年3期)2017-05-17
今日健康(2016年5期)2017-01-23
中国现代医学杂志(2015年26期)2015-12-23
