
沙地云杉 LBD 基因家族鉴定及盐胁迫响应分析
2023-10-17 14:05王亚萍隋明明白玉娥
西北植物学报 2023年9期
王亚萍, 隋明明,白玉娥
(内蒙古农业大学 林学院,呼和浩特 010019)
沙地云杉(Piceamongolica)为松科(Pinaceae)云杉属(Picea)常绿乔木,是中国珍稀濒危树种,也是北方地区防风固沙的优良树种,其自然分布在内蒙古浑善达克沙地,具有耐旱、耐寒、耐沙埋的优良抗性。因此吸引很多学者研究其分布、栽培、育苗和生理生化、体细胞胚胎发生等[1],但其分子生物学相关研究极少,特别是抗性基因挖掘研究有限,限制了其分子育种及抗性改良研究进程。
转录因子(TFs)通过调控基因表达来参与植物许多重要的植物发育过程,如细胞形态发生、信号转导和环境胁迫反应[2]。其中,LBD基因家族是高等植物一大类转录因子之一,具有高度保守的侧器官边界(LOB)结构域。LOB结构域长度约为100个氨基酸,包含3个特定的保守结构域,从N端到C端,分别是C结构域(C-block)、Gly-Ala-Ser区域(GAS-block)和类亮氨酸拉链(LX6LX3LX6L)。C结构域是DNA结合所需的,由高度保守的CX2CX6CX3C半胱氨酸基序组成;类亮氨酸拉链(LX6LX3LX6L)主要负责蛋白质二聚化过程;GAS基序由49个氨基酸残基组成,影响蛋白生物功能[3-4]。根据LOB结构域不同,LBD蛋白可分为ClassⅠ类和ClassⅡ类。Ⅰ类LBD蛋白都含有C结构域,GAS区域与亮氨酸拉链结构;而Ⅱ类蛋白只包含1个不完整的亮氨酸拉链结构,不能形成螺旋结构[5-7],然而,进一步将Ⅰ类细分为Ⅰa~Ⅰe 5个亚类,Ⅱ类分为Ⅱa和Ⅱb 2个亚类[8-9]。首个被发现的LBD基因是拟南芥中的AtLOB,该基因的表达存在组织特异性,只在侧生器官的近轴端及侧根基部表达[5-6]。近年来,随着植物基因组和转录组数据的不断公布,分别在拟南芥[10]、烟草[11]、玉米[12]、水稻[6]、杨树[13]、葡萄[14]、番茄[15]及大豆[16]中鉴定到43,98,44,35,57,30,46,46个LBD基因。目前的研究表明,LBD基因家族在植物叶、根和花发育、组织再生及胁迫响应过程起重要调控作用。如拟南芥LOB及LBD6参与调控叶片发育[3],LBD16和LBD29参与调控拟南芥侧根起始[17],LBD15通过调控WUS基因参与顶端分生组织分化[18],LBD16、LBD17、LBD18及LBD19促进愈伤组织形成[19];水稻IG1基因调控水稻颖壳、花及雌配子体发育[20];大豆LBD12响应非生物胁迫(干旱、盐、冷)和IAA、ABA及SA激素处理进程[16];番茄LBD40介导茉莉酸途径负调控其抗旱性[21];香蕉LBD5参与茉莉酸信号转导提高其耐寒性[22];拟南芥LBD37、LBD38和LBD39通过抑制花青素的生物合成来响应氮胁迫[23]。尽管LBD基因家族目前已在多个物种中被鉴定和研究,但在沙地云杉中的鉴定和研究还鲜见报道。
本研究以白音敖包自然保护区生长的沙地云杉为材料,通过参考挪威云杉基因组及前期沙地云杉转录组相关数据,筛选并鉴定出30条沙地云杉LBD基因序列,利用生物信息学软件分析所有LBD的理化性质;并运用多重序列比对对其进行结构和保守性研究;构建沙地云杉、拟南芥及杨树LBD蛋白系统进化树,分析其进化关系;利用qRT-PCR检测LBD在沙地云杉不同组织及盐胁迫下的表达水平,以期为进一步探究沙地云杉LBD基因家族非生物胁迫功能及分子育种奠定基础。
1 材料和方法
1.1 试验材料与处理
采取内蒙古赤峰市白音敖包自然保护区沙地云杉同一母树的成熟种子,将其播种于内蒙古农业大学试验田,培养条件为25 ℃,16 h光照/8 h黑暗条件,生长1年的幼苗分成叶片、茎、茎尖、主根、侧根(3次生物学重复),所有样品经液氮速冻置于-80 ℃冰箱保存。将生长1年的沙地云杉幼苗进行200 mmol/L盐胁迫处理,分别在处理0,6,12,24 h后采取整株样本,液氮速冻后放置于-80 ℃冰箱保存。
1.2 沙地云杉LBD基因筛选及理化性质分析
为识别沙地云杉中的LBD蛋白,将从模式物种拟南芥基因组数据库(https://www.arabidopsis.org/)获得43个LBD基因的氨基酸序列作为探针,利用BioEdit 7.2.0软件[24]通过BLASTP比对和HMM比对2种方法对前期沙地云杉叶片转录组数据进行了局部BlastP搜索,使用默认参数和E值小于0.001,初步鉴定LBD转录因子候选基因,然后用SMART在线软件对候选LBD蛋白序列进行保守结构域分析,保留具有完整LBD保守结构域的序列,从挪威云杉基因组(https://plantgenie.org/)中进行比较,最终获得沙地云杉云杉LBD基因成员,并根据它们的基因编号进行命名。使用在线软件ProtParam(http://web. Expasy.org/protparam/)对沙地云杉LBD基因编码蛋白的理化性质进行分析。利用ExPASy(http://www.expasy.ch/tools/pi_tool.html)计算LBD蛋白氨基酸残基数、等电点(pI)、分子量(MV),利用CELLO v.2.5在线软件预测LBD蛋白的亚细胞定位。
1.3 沙地云杉 LBD 系统进化树构建
从Phytozome13数据库(https://phytozome-next.jgi.doe.gov/)获取57个杨树LBDs的蛋白序列,利用MEGA11软件对43个拟南芥LBDs、57个杨树LBDs及30个沙地云杉LBDs蛋白进行多重序列比对,再将多序列比对结果利用 MEGA11软件采用邻接法(neighbor-joining,NJ)构建LBD系统进化树,校验参数设置成1 000次。系统进化树使用Evolgenius在线软件(http://www.evolgenius.info/evolview)及Adobe Illustrator 2022进行美化。
1.4 沙地云杉LBD保守结构域分析
采用DNAMAN软件沙地云杉30个LBDs序列进行保守域分析。为分析沙地云杉LBDs蛋白的保守Motif,通过MEME(http://meme-suite.org/)在线软件对沙地云杉LBD进行保守Motif分析,将Motif数目设置为10,其他参数都被设置为默认值。
1.5 沙地云杉LBD的表达水平分析
使用TAKARA植物总RNA提取试剂盒分别从沙地云杉不同组织及盐胁迫处理样本提取总RNA,通过1.0%琼脂凝胶电泳及Nanodrop2000测定其RNA完整性和浓度。逆转录的cDNA产物在-20 ℃中保存,将cDNA稀释5倍后作为模板使用。使用Primer3设计qRT-PCR引物(表1)。采用罗氏实时荧光定量PCR仪进行扩增,以EF1α(MA_505653g0010)作为内参基因,采用2-ΔΔCT法计算各基因的相对表达量,并使用SPSS软件进行显著性差异分析。用TBtools和GraphPadPrism8软件分别制作热图和柱状图。每个处理3个生物重复,每个样本3个技术重复。

表1 PmLBD 基因家族表达分析的实时荧光定量引物
2 结果与分析
2.1 LBD 基因鉴定与蛋白质理化性质分析
在沙地云杉中共鉴定到30个LBD基因,并根据基因在挪威云杉数据库的编号进行命名为PmLBD1~PmLBD30。沙地云杉LBD基因的CDS长度从360(PmLBD3)~930 bp(PmLBD3);蛋白序列长度为119(PmLBD3)~309 aa(PmLBD15);分子量为10 545.15(PmLBD22)~33 359.17 D(PmLBD15);等电点为5.15(PmLBD7,PmLBD12)~9.26(PmLBD9);亚细胞定位结果显示,30个沙地云杉LBD蛋白都位于细胞核内,见表2。
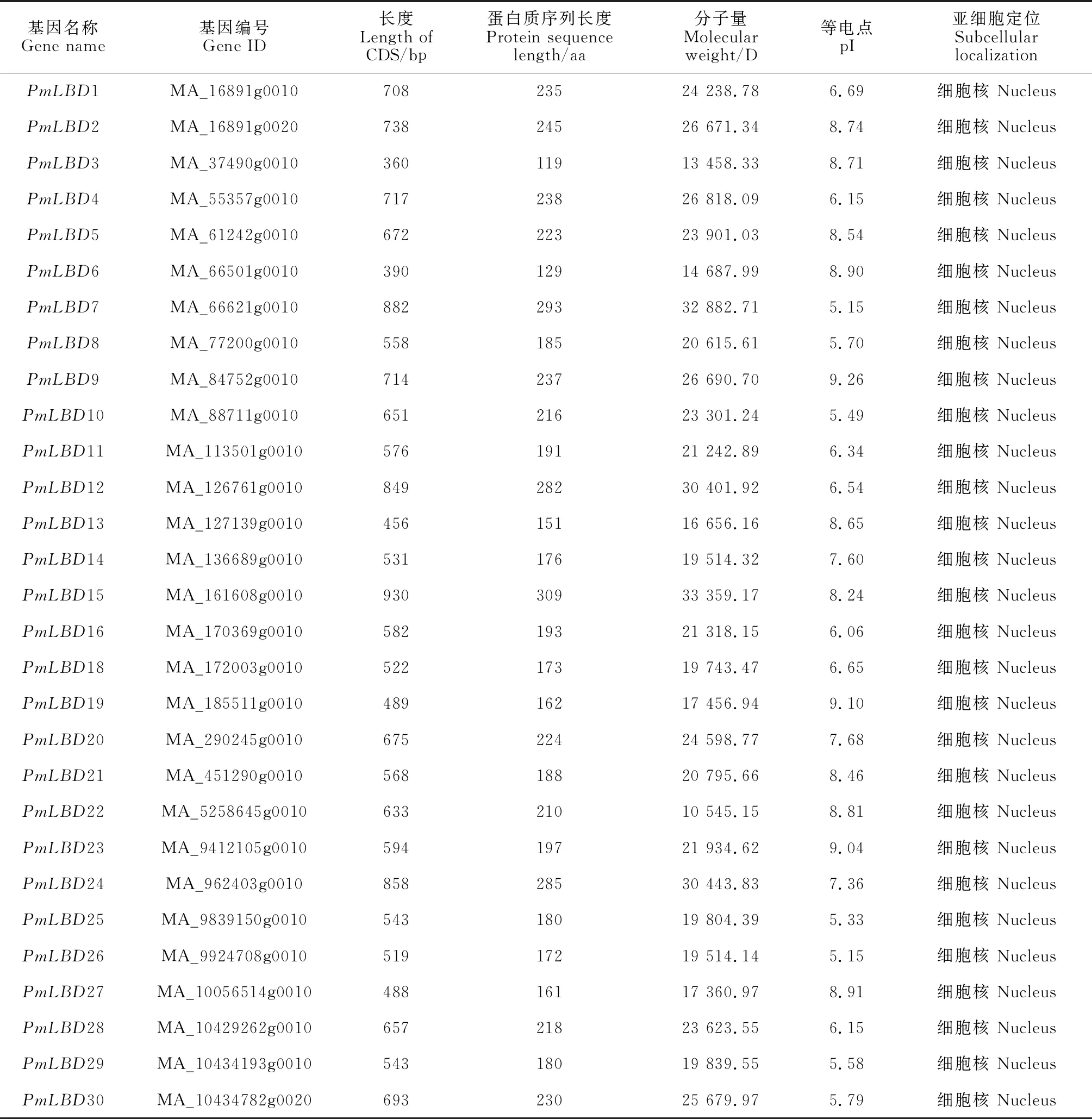
表2 沙地云杉 LBD 基因家族基本信息
2.2 沙地云杉LBD蛋白的保守结构域分析
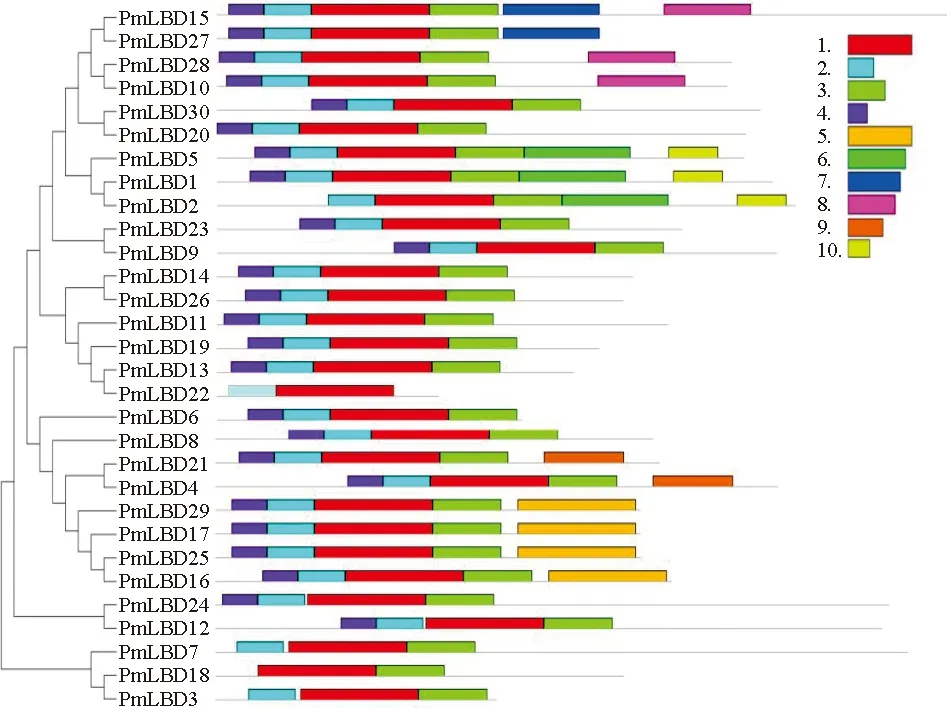
对30个PmLBD蛋白进行了多序列比对,所有PmLBD家族成员在N端都有1个高度保守的LOB区域,由约100个氨基酸组成(图1)。多序列比较表明,30个PmLBD均具有1个保守的c基序(CX2CX6CX3C)、GAS-block和亮氨酸拉链结构(LX6LX3LX6L)。说明PmLBD家族序列保守性较高,在沙地云杉内可能行使相似的生物学功能。为了进一步研究30个PmLBD结构的多样性,笔者利用MEME在线工具预测PmLBD蛋白的保守基序组成(图2)。结果表明,PmLBDs的序列特征识别出了10个基序,所有LBDs都包含motif1、motif2和motif3,构成了LOB结构域中高度保守的部分。除了PmLBD3/7/18外,其余27个PmLBD都具有motif4。基序在大多数序列中的相对位置相似,但不同PmLBD也有一定差异性。其中,motif5分布于PmLBD16/17/26/29中;motif6分布于PmLBD1/25中;motif7分布于PmLBD15/27中;motif8分布于PmLBD10/15/28中;motif9分布于PmLBD4/21中;motif10分布于PmLBD1/2/5中。
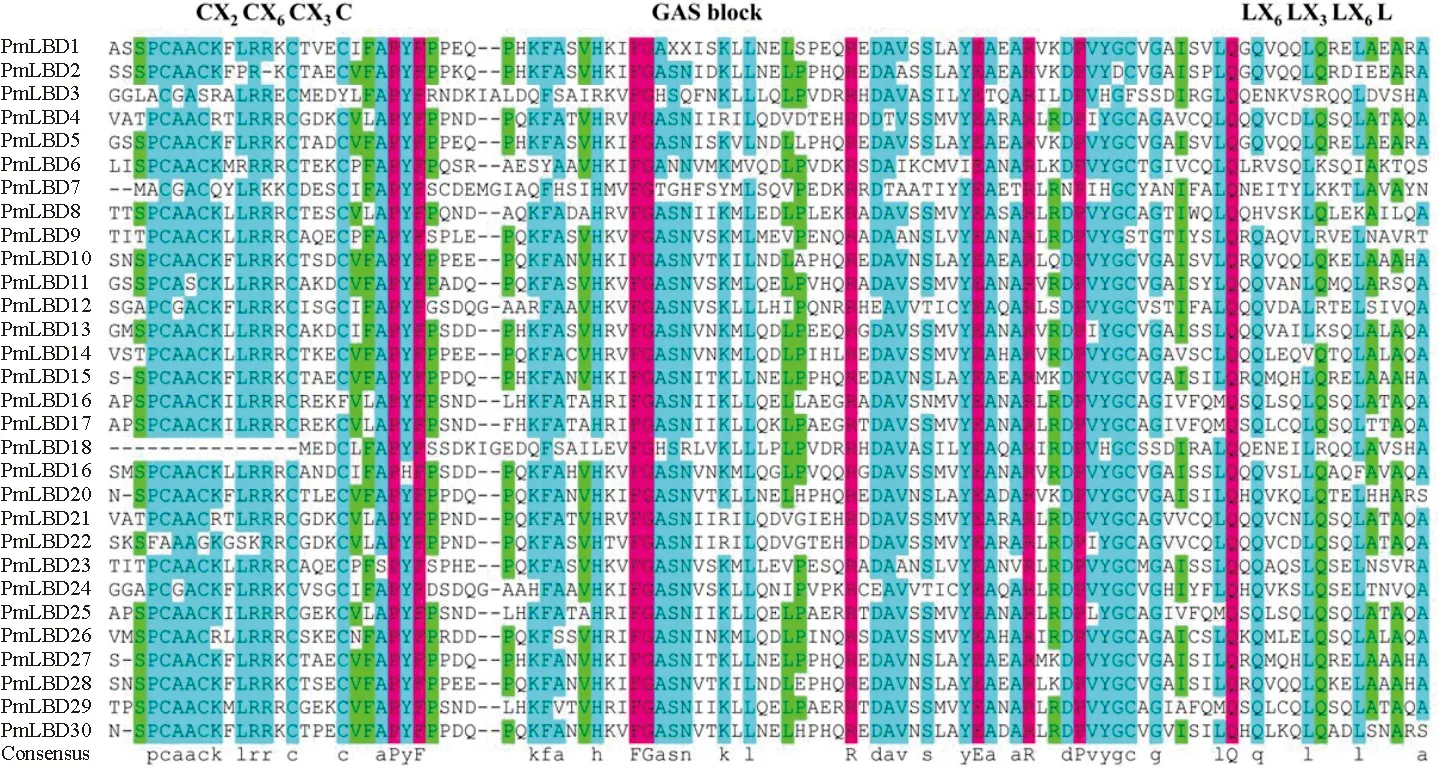
图1 PmLBD蛋白保守结构域序列比对
不同颜色的方块代表不同的基序(1-10)。
10个motif的氨基酸序列如图3,motif1为GAS保守结构域,motif2 为CX2CX6CX3C基序,motif3为LX6LX3LX6L。以上结果说明PmLBD具有多种生物学功能。
2.3 沙地云杉LBDs蛋白系统进化分析
为了深入了解沙地云杉PmLBD家族蛋白之间的进化关系,通过MEGA11将沙地云杉(30个)、杨树(57个)及拟南芥(43个)的130个氨基酸序列构建了系统发育树(图4)。根据模式植物拟南芥和杨树蛋白家族分类,将130个LBDs蛋白分为Class Ⅰ和Class Ⅱ,共7个保守的进化枝。其中,Ⅰ类有111个成员:杨树(41%)、拟南芥(32%)和沙地云杉(27%),分别为46,36,30个。Ⅱ类有19个成员:杨树(63%)和拟南芥(37%),分别为11,7个。进一步将Ⅰ类可以细分为5个子类:Class Ⅰa(14%),Class Ⅰb(27%),Class Ⅰc(23%),Class Ⅰd(24%),Class Ⅰe(12%);ClassⅡ可以细分为Ⅱa(58%)和Ⅱb(42%)。
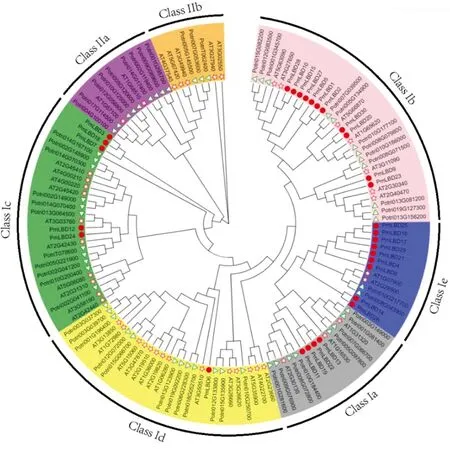
红色圆圈、绿色三角形和红色五角星分别代表沙地云杉、杨树及拟南芥。
其中,LBDs数量最多的是Class Ⅰb,分别包含12个PtLBDs,7个AtLBDs和11个PmLBDs;Class Ⅱb所含LBDs最少,仅有 3 个 AtLBDs 和5个PtLBDs;沙地云杉LBDs在Class Ⅰb中最多,有7个蛋白,但Class Ⅱ中无沙地云杉LBDs 蛋白,这可能与PmLBDs在3个物种中数量最少有关。
系统发育关系结果表明,沙地云杉的LBD蛋白与杨树的同源性高于拟南芥。
2.4 沙地云杉PmLBDs的组织特异性表达分析
为了进一步了解LBD基因在沙地云杉生长发育中的潜在功能,对LBDs在沙地云杉不同组织(茎尖、主根、侧根、茎和叶)中的表达水平进行分析。如热图(图5)所示,30个PmLBDs均被检测到,大部分LBDs在根和叶中具有较高的转录水平,而在茎尖中表达水平较低,表明它们在根和叶的发育中具有重要功能。此外,4个基因(PmLBD2/5/18/19)在茎中高表达,Class I b中的PmLBD9/20/23基因在侧根中强烈表达。以上结果表明,不同PmLBD在杨树各组织器官中功能具有差异。
2.5 沙地云杉PmLBDs在盐胁迫下的表达模式分析
为探究沙地云杉在盐胁迫条件下潜在的生物学功能,利用RT-PCR技术对沙地云杉30个LBD基因在盐胁迫处理下的表达情况进行了分析。结果(图6)表明在盐胁迫处理后,除了PmLBD3及PmLBD27基因外,28个沙地云杉LBD基因表达量均发生显著的变化,且同源性越近的PmLBD表达情况越相似。
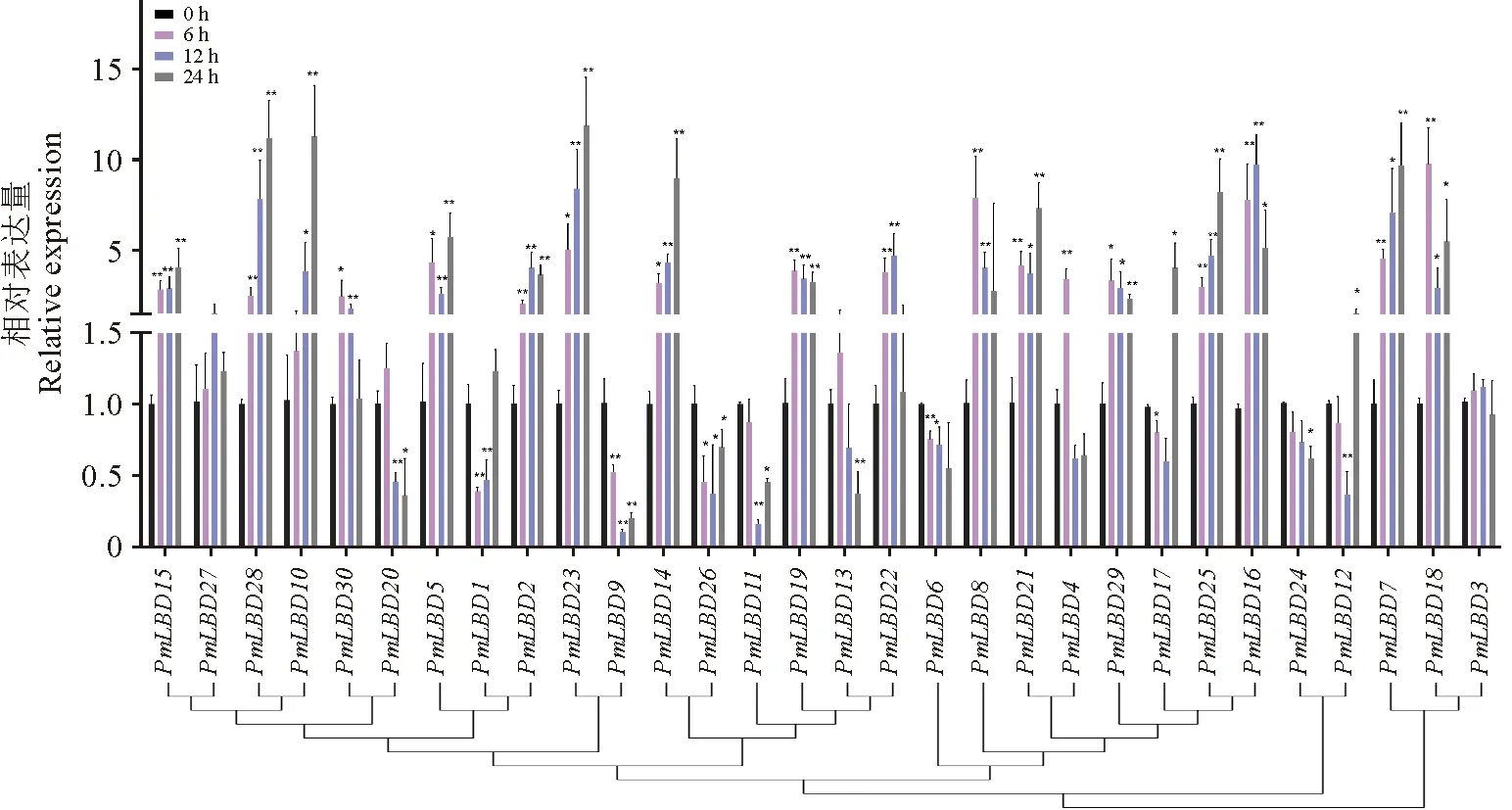
* P <0.05, ** P <0.01。
其中,17个PmLBD基因在盐处理后上调表达,不同的PmLBD基因在不同盐处理时间达到峰值,包括PmLBD8/18/19/29/30在6 h达到峰值及PmLBD2/16/22在12 h达到峰值,其余PmLBD基因在24 h达到峰值,特别是PmLBD10、PmLBD23及PmLBD28在盐处理24 h时上调10倍以上;而6个PmLBD基因在盐处理后下调表达,尤其是PmLBD9在12 h时下调表达10倍多。以上结果说明沙地云杉PmLBD基因家族成员在耐盐进程中可能发挥不同作用。
3 讨 论
LBD基因是1个植物特异性转录因子家族,已被证明在植物叶、根和花发育、组织再生及胁迫响应过程起重要调控作用;然而,目前尚未对沙地云杉进行LBD基因的系统研究。2013年,“云杉基因组项目”联合课题组在Nature上发表挪威云杉基因组草图,基因组大小为20 Gb,共注释约28 354个基因[25]。本研究通过参考挪威云杉基因组及前期沙地云杉转录组数据,筛选并鉴定了30条沙地云杉LBD基因序列(表2)。沙地云杉的基因组庞大,是拟南芥的100多倍,但在家族成员数量上,PmLBDs小于AtLBDs,这表明沙地云杉在进化过程中丢失了一小部分的LBD基因。多序列比较表明,30个PmLBD均具有1个保守的c基序(CX2CX6CX3C)、GAS-block和亮氨酸拉链结构(LX6LX3LX6L)(图1)。基因结构分析表明,不同LBD成员既有相似性也存在一定结构差异(图2)。这种差异可能是因为不同亚类的成员在进化过程中经历了基因片段的剪接或插入[26-27]。LBD亚类中相似的保守序列和基因结构通常意味着这些基因具有相似的生物学功能,这表明LBD家族的基序在整个进化过程中都很保守。
本研究构建了拟南芥、杨树和沙地云杉的LBD蛋白系统发育树,130个LBDs蛋白共分为Class Ⅰ和Class Ⅱ(图4)。以往的研究表明,拟南芥中86%的AtLBD基因、大豆中82%的GmLBD基因和玉米中84%的ZmLBD基因属于Ⅰ类[5,12,16]。同样,在本研究中,沙地云杉30个LBD均属于Ⅰ类。此外,Ⅰ类基因进一步被分为5个亚类。从分布看,Class Ⅰ a、Class Ⅰ b、Class Ⅰ c、Class Ⅰ d和 Class Ⅰ e中每个植物都存在LBD基因,且Class I b中的所有LBD基因共享1支较长的分支,说明这些LBD基因已发生了较大的变异,可能已经分化出新的功能。此外,同源PmLBD基因对在相同组织或相同胁迫下的表达丰度相似(图5和图6),表明沙地云杉中的PmLBD基因的重复可能导致了功能的冗余。
多项研究表明LBD蛋白参与调节植物分枝发育和花、茎、叶和根的形成[28-33]。基因的组织特异性表达可以为深入了解其在生长发育中的功能提供见解[34]。在本研究中,分析了PmLBDs在沙地云杉不同组织(茎尖、主根、侧根、茎和叶)中的表达水平模式,许多PmLBDs在特定组织中表现出较高的表达水平(图5)。例如,属于Class I b分支的PmLBD9、PmLBD20和PmLBD23在侧根中高表达,其在拟南芥中同源基因是AT2G30340(AtLBD13),研究表明AtLBD13、AtLBD16、AtLBD18和AtLBD29在调节拟南芥侧根发育中发挥重要作用[35-37],故推测其功能可能与AtLBD13相似,可作为调控沙地云杉侧根发育的候选基因。另外,笔者发现有4个基因(PmLBD2、PmLBD5、PmLBD18和PmLBD19)在沙地云杉茎组织中强烈表达,表明其在沙地云杉茎发育中起着重要作用。基于这些结果,推测沙地云杉PmLBD基因在其生长发育进程中起着关键作用,但具体的生物学功能其调控分子机制需要进一步验证。
据报道,MYB、WRKY、NAC等转录因子参与了抵抗非生物胁迫,帮助植物抵抗或适应环境的变化[38-40]。到目前为止,已有多项研究表明LBD基因在非生物胁迫反应中也发挥着重要作用[41-43]。在本研究中,qRT-PCR结果表明大部分沙地云杉LBD基因响应盐胁迫,其中57%的基因随处理时间表达上调,20%的基因表达下调(图6)。这些结果表明,PmLBD基因广泛参与沙地云杉的各种非生物胁迫反应。此外,众所周知,根系是响应干旱和盐胁迫的主要器官[44]。在本研究中,PmLBD10、PmLBD23及PmLBD28在根中强烈表达且在盐处理24 h时上调10倍以上,表明这些基因是调控沙地云杉盐胁迫的候选基因。以往的研究也表明,LBD基因在不同的非生物胁迫下显著上调表达。例如,马铃薯StLBD2-6和StLBD3-5是在干旱胁迫下显著上调[9];小麦Ta-6B-LBD81、Ta-4B-LBD51和Ta-U-LBD90在盐胁迫下表达上调,Ta-4A-LBD40和Ta-4D-LBD62在冷胁迫下表达上调,而Ta-2A-LBD13、Ta-2B-LBD15和Ta-2D-LBD18在干旱胁迫下表达上调[43]。值得注意的是在沙地云杉侧根中高表达的PmLBD9在12 h时下调表达10倍多,推测其可能是沙地云杉盐胁迫的负调控基因。在大豆中的研究也发现36个PvLBDs在盐及重金属胁迫处理中下调表达[45]。综上所述,PmLBD基因对盐胁迫表现出不同程度的反应,虽然其参与非生物胁迫反应的分子机制尚不清楚,但为开展沙地云杉进一步的抗性研究提供了候选基因。
4 结 论
沙地云杉 PmLBD蛋白家族包含30个成员,理化性质分析显示:蛋白序列长度在119~309 aa之间,分子量为10.5~33.4 kD,等电点为5.15~9.26,Cell-PLoc亚细胞定位显示均位于细胞核中。按照蛋白结构和聚类分析可以分为5个亚家族,Class I b与Class I e亚家族包含大部分成员。RT-PCR结果表明,沙地云杉PmLBD9、PmLBD20和PmLBD23也可能在侧根发生过程中发挥作用。另外发现大部分沙地云杉PmLBD基因响应盐胁迫,特别是PmLBD10、PmLBD23及PmLBD28在盐处理24 h时上调10倍以上,可作为调控沙地云杉盐胁迫的候选基因,但PmLBD9在盐胁迫中起相反作用。本研究初步揭示了大部分PmLBDs在植物生长发育及盐胁迫中发挥的作用,为沙地云杉PmLBDs基因功能进一步研究奠定了基础。
猜你喜欢
新医学(2023年10期)2023-12-09
农业现代化研究(2022年3期)2022-06-29
南方医科大学学报(2022年3期)2022-04-13
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
现代农村科技(2020年7期)2020-12-19
启迪与智慧·教育版(2018年12期)2018-02-26
山西林业科技(2017年4期)2018-01-23
甘肃林业(2016年2期)2016-11-07
华南师范大学学报(自然科学版)(2011年2期)2011-11-20
植物营养与肥料学报(2011年4期)2011-10-26
