
我国小麦田禾本科杂草对精唑禾草灵的抗药性研究进展
2023-10-16 03:35董立尧高海涛冯致科
植物保护 2023年5期
董立尧, 王 豪, 高海涛, 冯致科
(南京农业大学植物保护学院,农业农村部华东作物有害生物综合治理重点实验室,南京 210095)
小麦作为我国三大主粮之一,是重要的商品粮以及战略储备粮,小麦的产量直接关系到我国的粮食安全,据国家统计局统计,2021年全国小麦种植面积高达2 356万hm2,产量达1.37亿t,约占我国粮食总产量的20%[1]。农田杂草与作物争夺光照、水分、营养等而抑制作物的生长,制约粮食的高产与稳产。杂草是造成小麦产量损失的主要生物因素,我国每年因杂草危害造成的小麦减产达15%[2]。我国小麦生产主要分布在黄淮海和长江流域的河南、山东、河北、江苏、安徽和湖北等省[1],因气候环境及耕作方式的不同,小麦田优势杂草在各个地区不尽相同。近年来对小麦田杂草发生情况的调查显示,我国黄淮海和长江流域的小麦田优势禾本科杂草为日本看麦娘AlopecurusjaponicusSteud.、菵草Beckmanniasyzigachne(Steud.) Fern.、看麦娘AlopecurusaequalisSobol.、多花黑麦草LoliummultiflorumLam.、棒头草PolypogonfugaxNeesexSteud.、耿氏假硬草Pseudosclerochloakengiana(Ohwi) Tzvel.、野燕麦AvenafatuaL.、雀麦BromusjaponicusThunb.exMurr.、节节麦AegilopstauschiiCoss.、早熟禾PoaannuaL.等,优势阔叶类杂草为播娘蒿Descurainiasophia(L.) WebbexPrantl、荠菜Capsellabursa-pastoris(L.) Medic.、猪殃殃GaliumaparineL.、泽漆EuphorbiahelioscopiaL.、鹅肠菜Stellariaaquatica(L.) Scop.、繁缕Stellariamedia(L.) Villars等[2-6]。禾本科杂草的发生逐年加重,而且田间抗性杂草种类与种群不断增加,给杂草防除带来了新的挑战。
我国小麦田杂草防除主要依靠化学药剂,目前虽然登记证有1 410个(根据中国农药信息网统计),但实际上有效成分种类不是很多,同质化程度高。过去多年来常用的播后苗前土壤处理剂产品主要有异丙隆、绿麦隆、吡氟酰草胺等,防除禾本科杂草的苗后茎叶处理剂则主要是乙酰辅酶A羧化酶(ACCase)抑制剂和乙酰乳酸合酶(ALS)抑制剂类产品。ACCase抑制剂类除草剂的作用机制是抑制乙酰辅酶A羧化酶的活性,干扰脂肪酸的生物合成,最终导致植物死亡,我国小麦田广泛应用的品种主要有精噁唑禾草灵、炔草酯、唑啉草酯及其复配剂。ALS抑制剂是通过抑制植物支链氨基酸的合成导致植物生长受阻、死亡,用于小麦田的品种主要有甲基二磺隆、啶磺草胺、氟唑磺隆等及其复配剂。在生产中使用广泛的其他除草剂如苯磺隆、氯氟吡氧乙酸、2甲4氯、乙羧氟草醚等是用于防除小麦田阔叶杂草的[7]。综合来看,小麦田常用的土壤处理剂主要是异丙隆,防除阔叶杂草的茎叶处理剂主要是苯磺隆和双氟磺草胺等,而防除禾本科杂草的茎叶处理剂在过去很长时间内则以精噁唑禾草灵等为主[8]。随着苯磺隆和精噁唑禾草灵的长期使用,小麦田杂草抗性问题日益严重,一批高活性的新型除草剂的使用量在2010年前后迅速增加,如炔草酯、双氟磺草胺、唑啉草酯及其复配制剂成为中国除草剂市场主流。
精噁唑禾草灵自20世纪80年代后期开始在我国长江流域的小麦田使用,并成为防除小麦田禾本科杂草的主导药剂。由于这一药剂的长期单一使用,导致近年来禾本科杂草对它的抗性问题日益突出[9]。从我国多地小麦田采集的菵草[10]、日本看麦娘[11]、看麦娘[12]、多花黑麦草[13]、耿氏假硬草[14]、棒头草[15]和野燕麦[16]等种群已经对精噁唑禾草灵产生了明显的抗性。为了明确我国小麦田禾本科杂草对精噁唑禾草灵的抗药性整体情况,本文梳理了对精噁唑禾草灵产生抗性的主要杂草种类及相应抗性水平,归纳了其对精噁唑禾草灵的靶标抗性机理及代谢抗性机理,分析了抗精噁唑禾草灵杂草对不同除草剂的交互抗性及多抗性情况,提出了我国小麦田抗性杂草治理的初步方案。
1 我国小麦田主要禾本科杂草对精噁唑禾草灵的抗性现状
截至目前,我国小麦田发现抗精噁唑禾草灵的禾本科杂草至少有以下6种。
1.1 菵草
20世纪80年代末涂鹤龄报道了菵草在长江流域的稻茬麦田的发生量呈上升趋势[17]。从90年代起,菵草在小麦田迅速蔓延,成为小麦田主要杂草之一,形成了草害[18]。2007年张锁荣报道,苏南地区连续使用精噁唑禾草灵的稻麦连作田的菵草对精噁唑禾草灵均产生了一定的抗性,使用3年和5年的抗性指数(RI)分别为5.27和8.01,使用9年的RI为21.59[19]。吕波等发现2009年采集于江苏、安徽、上海小麦田的17个菵草种群中14个种群对精噁唑禾草灵产生了抗性,抑制杂草生长的有效中剂量(GR50)为0.133~19.198 g/hm2,句容种群抗性水平最高,RI为144.24[20]。李凌绪发现2010年-2012年采集于安徽、江苏、山东、湖北的59个小麦田菵草种群中15个表现出抗药性,庐江种群抗性水平最高,GR50达3 870 g/hm2[21]。潘浪发现2012年-2013年采集于江苏、安徽及上海的70个菵草种群中有27个对精噁唑禾草灵产生了抗性,抗性水平最高种群的GR50也达到2 560 g/hm2[10]。可见,长江中下游江苏、安徽及上海等地麦田的部分菵草对精噁唑禾草灵产生了明显的抗药性。
1.2 日本看麦娘
日本看麦娘在我国分布较广,几乎遍布全国,由于其较强的适应能力和喜湿特性,在长江流域的稻茬麦田和油菜田以及黄淮海流域等麦田中成为优势杂草。2007年杨彩宏等报道了句容油菜田的日本看麦娘对高效氟吡甲禾灵产生了抗性[22]。2009年郭峰在河南、湖北、江苏、山东和陕西等地麦田采集了11个种群,发现江苏练湖农场的种群对精噁唑禾草灵抗性水平最高,GR50达212.61 g/hm2[23]。毕亚玲发现2009年-2011年采自安徽、江苏、山东、河南和湖北5省的小麦田48个种群中24个对精噁唑禾草灵产生了抗性,安徽天长种群的抗性水平最高,GR50为1 182.30 g/hm2[8]。徐洪乐发现2010年-2012年采自江苏、安徽两地22个种群中安徽肥东种群抗性水平最高,GR50为1 556.08 g/hm2[24]。2019年-2020年许锦程等发现江淮地区的数十个种群均产生了抗性,GR50在223.89~ 429.78 g/hm2[25]。上述进展表明,我国长江流域小麦田的日本看麦娘对精噁唑禾草灵抗性十分严重。
1.3 看麦娘
看麦娘与日本看麦娘同属禾本科看麦娘属,也是我国农田主要杂草之一。2004年黄世霞最早报道江苏油菜田看麦娘对高效氟吡甲禾灵、精喹禾灵等产生了抗性[26]。郭文磊报道2013年-2015年采自安徽、江苏、山东和河南四省的77个看麦娘种群中有37个对精噁唑禾草灵产生抗性,高抗种群主要集中分布在安徽寿县、天长和江苏盱眙、姜堰,天长种群抗性最高,GR50为1 286.3 g/hm2[12]。卢红等发现2013年采自江苏、上海地区的23个看麦娘种群中有12个对精噁唑禾草灵产生抗性,江苏高邮种群的抗性水平最高,GR50为1 835.21 g /hm2[27]。陈秀涛测定了15个看麦娘种群,有10个产生抗性,RI在11.14~234.51之间[28]。看麦娘的抗性分布区域与日本看麦娘抗性种群的分布比较重合,有些田块抗性日本看麦娘、菵草、看麦娘等多种杂草混生,这可能与这些地区用药情况相似有关。
1.4 多花黑麦草
多花黑麦草又名意大利黑麦草,属于禾本科黑麦草属。多花黑麦草原产地中海区域,是我国引进的一个主要冬季牧草品种。近年来,在中国河南、江苏及湖北地区逐渐蔓延到小麦田,成为小麦田的一种恶性、抗性杂草。张佩发现2014年-2016年采自河南、江苏、山东等地的50个多花黑麦草种群中有42个对精噁唑禾草灵产生了不同程度的抗性,其中河南驻马店种群的抗性水平最高,GR50为2 218.41 g/hm2[13]。徐洪乐等也发现2016年-2017年采自河南新乡和驻马店小麦田的多花黑麦草对精噁唑禾草灵产生高水平的抗性, GR50最高为1 592.24 g/hm2[29]。Zhu等还发现抗啶磺草胺的多花黑麦草对精噁唑禾草灵产生了高水平的多抗性,GR50高达2 030.86 g/hm2[30]。Li等发现安徽地区的多花黑麦草对精噁唑禾草灵产生抗性,GR50在84.88~689.21 g/hm2[31]。
1.5 耿氏假硬草
耿氏假硬草,是亚热带和温带地区一种广泛分布的常见杂草,喜湿耐旱,抗盐耐瘠和耐阴,能在许多环境条件下生存,常见于潮湿的山地和低洼湿润地带。在国内其主要分布于上海、江苏、安徽、河南、山东等地区,主要存在于稻茬麦田,对小麦的危害非常严重。袁国徽报道2011年-2013年采自山东、河南、江苏等地62个耿氏假硬草种群中25个对精噁唑禾草灵产生高水平抗性(RI在14.7~103.2之间),GR50为438.1~3 033.1 g/hm2[14]。而高海涛检测了2014年-2015年随机采集于安徽、山东、江苏、河南、上海等地的30个耿氏假硬草种群,发现只有江苏江都种群达到高抗水平,GR50为159.03 g/hm2,抗性指数RI为15.68,其余29个种群对精噁唑禾草灵依旧敏感,抗性指数RI在1.00~2.40之间,GR50在10.14~24.32 g/hm2之间,也远小于田间推荐剂量(62 g/hm2)[32]。说明小麦田耿氏假硬草对精噁唑禾草灵抗性水平差别较大,袁国徽等调查的地块可能具有更长的精噁唑禾草灵用药历史[33]。
1.6 棒头草
棒头草属禾本科棒头草属,在我国,除东北地区外的其余地区均有发生,尤其在长江流域及其以南各地发生较严重。在部分地区已成为优势杂草。我国小麦田棒头草对精噁唑禾草灵的抗性尚缺乏系统研究,目前只有Zhao等报道了安徽怀远县的种群对精噁唑禾草灵产生较高水平的抗药性[34]。
2 杂草对ACCase抑制剂的抗药性机理
杂草对除草剂的抗药性机理复杂多样,但一般来说可以分为靶标抗性和非靶标抗性。靶标抗性包括靶标酶基因的突变以及靶标酶基因的过量表达。非靶标抗性即除草剂作用靶标酶以外的其他方式,非靶标抗性包括杂草解毒代谢能力增强、除草剂吸收减少、传导变慢、杂草抗氧化胁迫能力增强等(图1)[35]。
2.1 杂草抗ACCase抑制剂的靶标机理
精噁唑禾草灵是乙酰辅酶A羧化酶(ACCase)抑制剂类除草剂,用于防除禾本科杂草。由于其长期单一使用,目前禾本科杂草对精噁唑禾草灵的抗性问题比较严重。研究发现乙酰辅酶A羧化酶基因突变、表达量上调是其产生抗性的靶标酶机理。
2.1.1ACCase基因突变导致杂草对精噁唑禾草灵产生抗性
ACCase是一类生物羧化酶,结构上有3个功能区域,分别是生物素羧化酶(BC)、生物素羧基载体蛋白(BCCP)、羧基转移酶(CT)。该酶参与脂肪酸生物合成的第一步,即依赖三磷酸腺苷(ATP)催化乙酰辅酶A生成丙二酰辅酶A(malonyl-CoA),该反应也是植物脂肪酸生物合成的首要关键步骤。在当前大多数研究中ACCase基因发生特定突变会导致ACCase活性发生变化继而导致杂草对ACCase类抑制剂产生抗药性。ACCase氨基酸的变化被认为是杂草对ACCase抑制剂产生抗药性的重要机制。目前为止,已经发现7个位点的14种氨基酸取代与抗性相关(图2),分别是Ile-1781-Val,Ile-1781-Leu,Ile-1781-Thr,Ile-1781-Glu,Trp-1999-Cys,Trp-1999-Leu,Trp-1999-Ser,Trp-2027-Cys,Ile-2041-Asn,Ile-2041-Val,Asp-2078-Gly,Cys-2088-Arg,Gly-2096-Ala和Gly-2096-Ser[36-41]。

图2 ACCase 羧基转移酶(CT)区的抗性突变位点[39]Fig.2 Herbicide-resistant mutation sites in carboxyl transferase domain of ACCase
Xu等对我国日本看麦娘抗精噁唑禾草灵的机理研究发现,ACCase氨基酸发生了Trp-2027-Cys,Asp-2078-Gly,Ile-1781-Leu,Ile-2041-Asn,Trp-1999-Cys,Trp-1999-Leu的5个位点6种突变[42-43]。通过对抗性及敏感日本看麦娘ACCase的提取及测定,发现在精噁唑禾草灵不同浓度下,抗性和敏感日本看麦娘ACCase活性变化不同,敏感日本看麦娘ACCase活性快速显著下降,而抗性日本看麦娘的ACCase活性下降幅度小、速度慢,其活性被精噁唑禾草灵抑制程度显著低于敏感型。Pan等筛选出的27个抗精噁唑禾草灵菵草种群中均存在靶标位点ACCase的突变, 共有5种突变类型,分别为I1781L,W2027C,I2041N,D2078G,G2096A[36- 37, 44]。这些位点突变是导致菵草对精噁唑禾草灵产生抗性的主要原因。同样的,在对这些不同突变的抗性种群ACCase进行酶活测定时,发现抗性种群ACCase对精噁唑禾草灵的敏感性均低于敏感种群。Xia等[45]、Guo等[46]和Wang等[47]在抗精噁唑禾草灵看麦娘种群中发现4处突变,分别为I1781L, I2041N, I2041T和D2078G。Zhang等对我国抗精噁唑禾草灵多花黑麦草的基因序列分析,发现氨基酸Ile-1781-Leu、Asp-2078-Gly、Ile-2041-Asn、Cys-2088-Arg的突变是导致多花黑麦草对精噁唑禾草灵产生抗性的原因[48]。高海涛[32]、袁国徽[33]在抗精噁唑禾草灵耿氏假硬草中发现了Trp-1999-Cys和Trp-1999-Ser氨基酸突变。Zhao等[34]在抗精噁唑禾草灵棒头草种群中发现了Trp-1999-Ser突变。由此可见,ACCase基因突变是麦田禾本科杂草对精噁唑禾草灵产生抗性的重要机理之一。
通常来说,靶标酶ACCase基因特定位置的突变均会引起ACCase蛋白结构的改变从而导致对精噁唑禾草灵敏感性下降。但是也有研究发现,ACCase在未发生基因突变的情况下,ACCase活性改变亦会导致杂草抗药性的发生。如张佩[13]发现采自江苏省的多花黑麦草种群JLJY-6对精噁唑禾草灵的抗性指数高达27.96。但对其ACCase基因序列进行研究时未发现任何已知导致抗性的突变位点,且对抗性靶标基因表达量的研究中,抗性和敏感种群的表达量也没有显著性的差异。并且江苏省抗性种群对除了精噁唑禾草灵外的其他ACCase类抑制剂如精喹禾灵、高效氟吡甲禾灵、炔草酯、烯草酮、烯禾啶和唑啉草酯等药剂敏感;对ALS类、光合抑制剂类等除草剂都没有产生抗性。但是在对其抗性种群的ACCase活性研究发现,抗性种群的IC50是敏感种群的9.85倍。这说明江苏省多花黑麦草抗性种群JLGY-6对精噁唑禾草灵产生抗药性的主要原因是靶标酶活性的降低引起的。由此可见,在ACCase未发生基因突变的情况下,ACCase酶活性的改变也会导致对精噁唑禾草灵产生抗性。
2.1.2ACCase的过量表达导致杂草对精噁唑禾草灵产生抗性
2001年有学者首次提出ACCase的过量表达与此类除草剂抗性相关。Bradley等在研究石茅Sorghumhalepense(L.) Pers.对芳氧苯氧基丙酸酯类(APPs)和环己烯酮类(CHDs)除草剂的抗性机理时发现抗性和敏感种群之间ACCase的相对活性没有差异,但是抗性种群的绝对活性高于敏感种群[49]。他们认为导致这种情况发生的原因是抗性石茅的ACCase过量表达。徐洪乐利用筛选的内参基因检测了抗性和敏感日本看麦娘在精噁唑禾草灵处理前后,根、茎和叶中ACCase基因表达量,发现在药剂处理后2 h的叶中,药剂处理后3、5 d和7 d的茎中,抗性日本看麦娘ACCase基因的表达量显著高于敏感日本看麦娘[24]。潘浪通过分析敏感和抗性菵草ACCase基因表达量的变化,发现在精噁唑禾草灵处理前和处理后,5个抗性菵草种群中ACCase基因的表达量均高于敏感种群[10]。张佩发现多花黑麦草敏感和抗性种群ACCase表达量变化趋势都是先经过一个缓慢的上升过程到药后24 h达到最大值,然后下降,5~7 d之后已不再变化。抗性突变种群的ACCase基因表达量在药后24 h上升到最大值,是敏感种群的2倍,之后又缓慢下降,但是仍然比敏感种群表达量高2~3倍[13]。
2.2 杂草抗ACCase抑制剂的非靶标机理
非靶标抗性机理(non-target site resistance,NTSR)越来越受到重视,逐渐被认为也是导致杂草对ACCase抑制剂类除草剂产生抗药性的重要原因之一。非靶标抗性机理非常复杂,抗精噁唑禾草灵的非靶标机理研究很少,而且集中在杂草解毒代谢能力方面,20世纪90年代,也有发现个别杂草可以通过膜的去极化对ACCase抑制剂类除草剂产生抗性[50]。
2.2.1相关代谢酶解毒代谢能力增强导致杂草对精噁唑禾草灵产生抗性
2.2.1.1代谢酶解毒代谢的一般过程
目前的研究认为,代谢抗性是非靶标抗性中最重要的部分。除草剂在植物体内的代谢是一个复杂的过程,涉及不同种酶之间的协作(表1)。这一过程大致可以分为3个阶段(图3):1)除草剂分子被转化成一个更加亲水的代谢物,这一阶段涉及细胞色素P450氧化酶系、酯酶、水解酶、氧化酶和过氧化物酶;2)与植物体内受体分子结合,涉及的酶主要有谷胱甘肽S-转移酶和糖基转移酶;3)进一步轭合和氧化,代谢物被排到液泡或者细胞壁进行进一步降解,主要涉及ABC转运蛋白[51-52]。

表1 植物对除草剂代谢过程中涉及的相关酶1)Table 1 Enzymes involved in the metabolism of herbicides in plants

图3 精噁唑禾草灵在杂草体内代谢涉及的相关酶[51]Fig.3 A variety of metabolizing enzymes involved in metabolism of fenoxaprop-P-ethyl in weeds
2.2.1.2细胞色素P450氧化酶系与杂草抗精噁唑禾草灵
1985年和1986年分别首次报道对靶标位点不同的除草剂产生交互抗性的大穗看麦娘AlopecurusmyosuroidesHuds.和硬直黑麦草LoliumrigidumGaud.的抗性是由于细胞色素P450氧化酶系活性增强而对除草剂的代谢增强所致[53-54]。目前已有个别细胞色素P450氧化酶系介导的抗性杂草生物型被报道。Wang等发现早熟禾P.annuaL.对精噁唑禾草灵具有天然耐药性。在施用精噁唑禾草灵后细胞色素P450和b5的含量显著高于施用吡唑解草酯和未做处理的对照植株。用模式底物方法间接测量细胞色素P450酶活性,发现其在施用精噁唑禾草灵后活性升高。因此认为,细胞色素P450氧化酶系可能在早熟禾对精噁唑禾草灵耐药性方面起重要作用[55]。但是在其他ACCase和ALS类抑制剂的抗性研究中发现了几个重要的CYP代谢酶基因并验证了其功能。Zhao等在研究看麦娘对甲基二磺隆的抗性时,利用转基因拟南芥Arabidopsisthaliana(L.) Heynh.验证了CYP709C56过量表达会导致看麦娘对甲基二磺隆产生抗性[56],Pan等在抗五氟磺草胺光头稗Echinochloacolona(Linnaeus) Link研究中利用转基因水稻证明了CYP81A68过量表达会导致其抗性[57]。Bai等在抗ALS抑制剂菵草中发现了CYP99A44 和CYP704A177两个CYP基因在代谢抗性中起到重要作用[58]。
2.2.1.3谷胱甘肽S-转移酶与杂草抗精噁唑禾草灵
GST可催化生物体内源或外源化合物与谷胱甘肽(glutathione,GSH)的巯基轭合,改变亲脂分子的亲电子中心而形成低活性的最终产物。另外,GST还具有依赖于GSH的过氧化、异构化(isomerization)功能及一些其他非催化活性功能,如非底物配体、胁迫引导的信号处理、细胞凋亡预防等[59]。Cummins等研究发现GST对精噁唑禾草灵代谢活性及GSH过氧化酶(glutathione peroxidase,GPOX)活性的加强导致大穗看麦娘A.myosuroides对精噁唑禾草灵产生抗性[60]。
在GST介导的代谢除草剂安全剂应用中的数据显示,GST基因可被诱导性表达。此外在杂草研究中发现,作物中GST基因在代谢除草剂中发挥了一定作用。Pan等研究发现菵草GST基因GST-T3、GST-U1和GST-U6在精噁唑禾草灵处理后呈现表达量上调[52]。基因的表达上调可增强杂草对除草剂的代谢能力。
2.2.1.4ABC转运蛋白与杂草抗精噁唑禾草灵
很多研究证明除草剂在植物液泡中代谢,未见将ABC转运蛋白与杂草抗精噁唑禾草灵相联系的研究报道。但是,在作物和模式植物中已经确认ABC转运蛋白参与代谢除草剂。有研究显示,拟南芥和其他物种中的一些ABC转运蛋白能够转运不同除草剂和除草剂代谢产物。ABC转运蛋白同样在安全剂诱导下上调表达,因此可以协调过多的谷胱甘肽轭合物。一个参与GS-轭合物转运的ABC转运蛋白AtOPT6在氟嘧磺隆处理后也可出现上调[61]。近期的研究发现,对草甘膦(glyphosate)处理的光头稗进行转录组测序,通过RNA-Seq和RT-qPCR双重验证筛选出了2个重要ABC转运蛋白基因ABCC8和ABCC10,通过转基因水稻进一步验证了该基因可以通过将杂草体内草甘膦转运至液泡减少药剂到达靶标的量从而导致抗药性[62]。
印刷行业现在最缺的可能就是,数字化的人才,这类人才,不单要熟练操作数字化设备,还应娴熟运用各类数字化软件,李新立坦陈自己的忧虑。诚如他所说,传统印刷在国内经过这二十多年的大发展,已经储备了大量的人才,但在数字化的时代,一切应用都跟数据有关,行业及企业要想赢得未来,精通数字语言的人才就是关键。而如何破局,还需群策群力。
2.2.2杂草对除草剂吸收减少或传导变慢
已有的研究表明,对除草剂及其有毒代谢物的屏蔽作用和隔离作用被认为是杂草对除草剂抗性的一个重要机理。如飞蓬属ErigeronL.的抗药性生物型中,发现百草枯的移动受到了限制,并且叶绿体的功能如CO2固定和叶绿素荧光猝灭可以迅速恢复。这说明除草剂在其作用位点的结合可能被阻止[63]。但是目前未见杂草对精噁唑禾草灵吸收和传导的差异导致抗药性的报道。
2.2.3杂草抗氧化胁迫能力增强
有研究认为小RNA等调控因子参与的调控机制在杂草体内发挥着重要的作用,也极有可能参与杂草对除草剂的非靶标抗性并导致抗性的产生。
目前对植物中的漆酶(laccase)研究甚少,漆酶的许多生物学功能未知。已知植物中漆酶是小RNA397(miR397)的靶基因。Pan等[52]在抗精噁唑禾草灵菵草种群中发现4个氧化酶和过氧化物酶基因表达上调。而ptr-mir397a可调节其靶基因漆酶基因,随后漆酶基因再调控其下游的转录因子及氧化酶/过氧化物酶基因的表达,从而形成调控网络调节木质素生物合成。鉴于氧化酶/过氧化物酶在杂草对除草剂的抗性中可以发挥重要的作用,推测抗性菵草中也形成了这样一个miR397调控的网络,并最终导致其对精噁唑禾草灵产生抗药性。
3 抗精噁唑禾草灵杂草的治理策略
3.1 加强抗精噁唑禾草灵杂草的检测
抗性杂草的治理首先要发现抗性杂草、评价杂草的抗药性水平。通常使用整株生物测定法确定某杂草是否对精噁唑禾草灵产生抗性及其抗性水平。通过剂量反应曲线计算出除草剂抑制杂草生长50%的剂量(ED50或GR50)。ED50(effective dose of 50% inhibition in plant growth)即抑制植物生长的有效中剂量。GR50(herbicide dose required 50% growth reduction)即鲜重抑制中量。求出疑似抗性杂草与敏感杂草的ED50或GR50比值,即是某杂草种群对敏感杂草生物型的抗性指数(resistance index,RI)。2005年南京农业大学除草剂毒理及抗药性实验室根据杂草抗药性的实际,制定了杂草抗药性研究的分级标准:敏感RI≤2;敏感水平下降2
随着杂草抗药性研究的不断深入,杂草抗药性检测方法也不断创新,目前广泛应用的生物测定方法包括整株生物测定法、种子生物测定法和培养皿快速检测法,还有快速分子生物学测定方法,如靶标酶编码基因测序法、衍生酶切扩增多态性(derived cleaved amplified polymorphic sequence,dCAPS)法和环介导等温扩增PCR(loop-mediated isothermal amplification,LAMP)法等。目前在我国小麦田杂草对精噁唑禾草灵抗性研究中最常用的方法为整株生物测定法、靶标酶编码基因ACCase测序法、dCAPS, LAMP等。其中整株生物测定法、靶标酶基因ACCase测序是研究抗精噁唑禾草灵杂草必需步骤,而dCAPS和LAMP方法是快速准确诊断抗药性突变型杂草的方法。李凌绪[21]、潘浪[10]对抗精噁唑禾草灵的菵草,徐洪乐[24]对抗精噁唑禾草灵的日本看麦娘,郭文磊[12]对抗精噁唑禾草灵的看麦娘,袁国徽[14]对抗精噁唑禾草灵的耿氏假硬草,张佩[13]对抗精噁唑禾草灵的多花黑麦草,分别建立了dCAPS分子检测技术。潘浪针对菵草对精噁唑禾草灵产生抗性的I1781L,I2041A,G2096A突变建立了LAMP检测技术[10]。
杂草抗药性的检测方法各有利弊,研究中要根据除草剂的作用机理,杂草的基本属性,研究目的等综合分析决策选用合适的抗性检测方法。
3.2 避免使用与精噁唑禾草灵有交互抗性的除草剂
交互抗性指抗性杂草生物型对化学结构相似或作用机理相同的其他除草剂产生的抗性,这在抗性杂草中普遍存在。抗精噁唑禾草灵杂草生物型产生后,ACCase类的除草剂则对该类抗性杂草防效较低,甚至完全无效,严重威胁粮食的丰产丰收,给杂草科学和除草剂科学研究提出了新的挑战。目前,国内抗精噁唑禾草灵杂草对ACCase类除草剂均产生了不同程度的交互抗性,特别是APP类的炔草酯、精喹禾灵及精吡氟禾草灵的交互抗性较为严重(表2),在防除抗精噁唑禾草灵的抗性杂草中要避免使用此类药剂加重杂草抗药性。为便于大家回避交互抗性的除草剂,我们归纳了ACCase氨基酸上的7个位点14种突变会导致杂草对ACCase抑制剂类除草剂产生的抗性及其交互抗性情况(表3)。
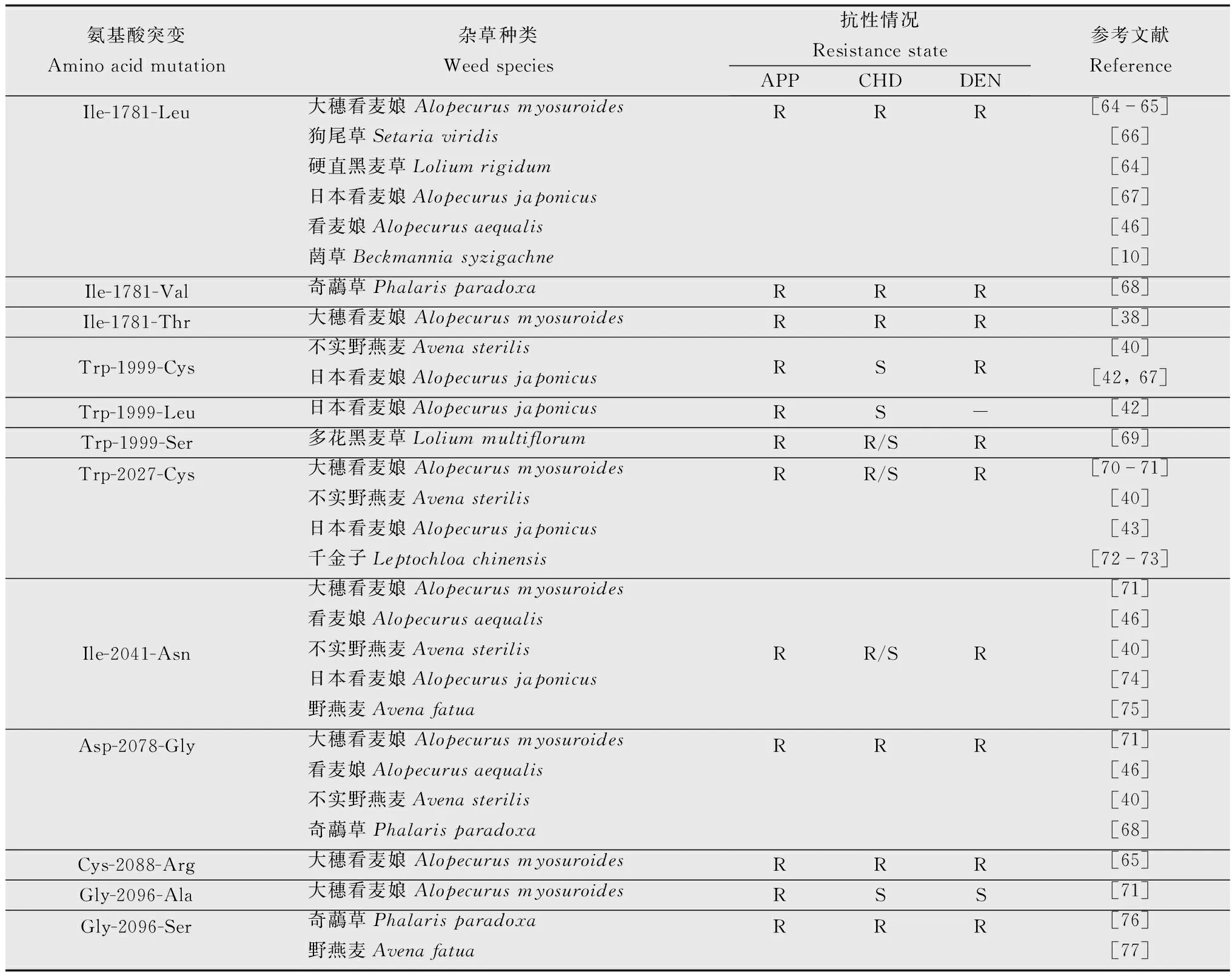
表3 ACCase氨基酸突变导致杂草对ACCase类抑制剂类除草剂的交互抗性1)Table 3 Cross-resistance to ACCase-inhibiting herbicides caused by amino acid mutations in ACCase
3.3 筛选能够治理抗精噁唑禾草灵杂草的除草剂
科学合理地选用除草剂防除抗精噁唑禾草灵杂草需要在明确杂草抗药性机理的基础上,清楚杂草的交互抗性及多抗性,科学界定针对该抗性杂草应禁用或限用的除草剂类别或产品,最终选择合适的除草剂。针对抗精噁唑禾草灵菵草种群,潘浪发现甲基二磺隆、啶磺草胺、异丙隆对抗性菵草具有良好的防效[10];徐洪乐等对抗精噁唑禾草灵日本看麦娘种群的多抗性研究发现,其对氟唑磺隆、甲基二磺隆、磺酰磺隆、啶磺草胺、乙草胺、扑草净、绿麦隆表现敏感[24]。抗精噁唑禾草灵的耿式假硬草[32]和抗精噁唑禾草灵的多花黑麦草[13]尚未对甲基二磺隆、啶磺草胺、扑草净、异丙隆等除草剂产生多抗性。尽管抗精噁唑禾草灵的杂草对上述ALS类抑制剂的多抗性不严重,对一些常用的ALS类抑制剂如甲基二磺隆、啶磺草胺等仍然敏感,但是这类药剂的抗药性风险很高,在防治时尽量回避这类药剂。
作为防除小麦田禾本科杂草的主要除草剂精噁唑禾草灵及同类除草剂炔草酯等的抗药性及其对ACCase类除草剂的交互抗性和对其他除草剂的多抗性不断被报道,设计开发新型作用机理的除草剂成为当前防除抗性杂草的急切要求。由青岛清原作物科学有限公司开发的新型化合物环吡氟草酮是HPPD抑制剂类除草剂,对小麦田的各种看麦娘属杂草(含抗性)表现出很好的防除效果[78]。
4 问题与展望
我国学者在麦田抗精噁唑禾草灵杂草研究方面取得了很大的进展,明确了抗性杂草的种类、揭示了抗性机理以及初步提出了抗性杂草治理的策略。但是在杂草对精噁唑禾草灵的非靶标机理研究方面不够系统、不够深入,在精噁唑禾草灵吸收传导差异方面未见报道,代谢精噁唑禾草灵的具体途径、代谢产物,筛选并验证能够代谢精噁唑禾草灵的功能基因及功能基因的调控途径等方面尚处空白。近年来,随着精噁唑禾草灵对小麦田杂草的防效不断下降,代替性除草剂啶磺草胺、甲基二磺隆等除草剂在小麦田的使用规模不断扩大,抗精噁唑禾草灵杂草的多抗性问题开始逐渐显现,这种现象值得我们警惕。2020年李君君等报道了日本看麦娘对精噁唑禾草灵的代谢抗性导致了其对高效氟吡甲禾灵、甲咪唑烟酸、双草醚、甲基二磺隆等产生中高水平的抗性[79]。周志恒等发现2020年采集的抗精噁唑禾草灵的野燕麦对ALS类啶磺草胺、PSⅡ 类绿麦隆和HPPD类环吡氟草酮表现出了敏感性下降[16]。因此,轮用不同作用靶标、较低选择压的除草剂是控制抗精噁唑禾草灵杂草蔓延的重要措施,而且抗性杂草的治理还需要与耕作、栽培等农艺措施结合起来,如作物轮作能避免在农田中长期单一使用除草剂。
猜你喜欢
草业科学(2022年10期)2022-11-18
风流一代·青春(2021年7期)2021-08-09
草地学报(2021年7期)2021-08-03
世界农药(2019年3期)2019-09-10
农药科学与管理(2019年10期)2019-04-20
南方农业·下旬(2017年11期)2018-01-09
散文选刊·下半月(2017年3期)2017-11-07
新农业(2017年2期)2017-11-06
中国饲料(2017年15期)2017-08-22
杂草学报(2015年2期)2016-01-04
