
大豆GmGST7 基因耐酸铝功能研究
2023-10-12 04:02金晓雨王金玉程艳波连腾祥马启彬
华南农业大学学报 2023年5期
胡 康 ,金晓雨,张 雪,王金玉,程艳波,连腾祥,年 海,马启彬
(华南农业大学 农学院/国家大豆改良中心广东分中心/亚热带农业生物资源保护与利用国家重点实验室/广东省分子育种重点实验室/岭南现代农业科学与技术广东省实验室, 广东 广州 510642)
据统计,全球范围内有超过50%的潜在可耕地可能是酸性的,世界上多达60%的酸性土壤在发展中国家,我国南方土地酸化也面临严峻的形势[1]。铝毒是世界范围内限制作物生产的一个重要因素,当土壤pH 下降到5 以下时,铝在土壤中以八面体六水Al(H2O)63+形式存在,通常称为A13+。这种形式的铝溶解到土壤溶液中后,对植物的根系造成严重胁迫[2-3]。酸铝胁迫对植物影响最快最主要的是根系,尤其是对根尖生长的抑制[4-6]。植物根尖与酸铝接触后在极短的几分钟内便可快速进入细胞质[7-9]。当细胞质内进入大量铝元素后,会抑制细胞分裂,迅速破坏根系细胞的扩张和伸长[10]。植物根系若长期暴露在酸铝胁迫下,根系的延伸会受到抑制,根尖膨胀,侧根、根毛的数量显著下降[11]。酸铝胁迫下损伤后的根系对水和矿物质等养分的吸收能力显著下降[12-13]。酸性条件下的铝离子具有较高的活性,对细胞壁、细胞膜、细胞质和细胞核的生理功能都会产生影响[14]。
华南农业大学国家大豆改良中心广东分中心前期研究发现,GsMYB7受酸铝胁迫诱导表达,在酸铝敏感大豆品种‘华春6 号’中过表达,可增强植株对耐酸铝胁迫的抗性。对野生型和GsMYB7过表达株系进行酸铝处理后,RNA-Seq 测序结果分析和RT-qPCR 检测结果发现,有9 个候选基因被GsMYB7间接或直接调控,其中,GmGST7对酸铝胁迫响应比较明显[15]。
谷胱甘肽转移酶(Glutathione S-transferase,GST)家族成员起源于一个古老的祖先基因,在漫长的进化过程中,经历了外显子改组、基因复制、选择性剪接、交换、突变和其他未知机制,导致了GST 的序列多样化和功能多样性[16-18]。根据氨基酸序列的相似性和外显子的个数,GST 家族在植物中主要分为4 个类型:phi 型、zeta 型、tau 型和theta 型[19]。在结构上,GST 含有N 端和C 端保守域。GST 通过将谷胱甘肽偶联到疏水底物上,对内源性和外源性化合物进行解毒[20]。解毒包括3 个阶段:转化、结合和分隔[21]。经典的GST 有2 个结合位点,位于N 端的GSH 结合位点(G-site) 和由C 端形成的亲电底物结合位点(H-site)。
GST 在植物中参与除草剂解毒、激素生物合成、酪氨酸降解、过氧化物分解等过程,且环境应激反应比较敏感[22],如除草剂和低温诱导AtGSTU26表达[23],病原体感染诱导AtGSTU16表达[24]。研究发现,GST 可通过调解氧化还原信号参与植物根系的发育[25-26]。
1 材料与方法
1.1 试验材料
大豆GlycinemaxL.‘华春6 号’、本氏烟草NicotianaL.以及野生型拟南芥(Col-0)Arabidopsis thalianaL.,均由国家大豆改良中心广东分中心提供。所用菌株为大肠埃希菌EscherichiacoliDH5α、根癌农杆菌AgrobacteriumtumefaciensGV3101。
1.2 生物信息学分析
使用NCBI 大豆(https://www.ncbi.nlm.nih.gov/)和Phytozome 数据库(https://phytozome- next.jgi.doe.gov/),查询基因序列、蛋白家族,使用MEGA11软件对GmGST7进行cDNA 和氨基酸序列比对,完成进化树分析。
1.3 大豆试验样品处理
选取饱满无病害的‘华春6 号’种子,用营养土在人工气候培养间进行培育,在其幼荚期取幼荚、花、叶、茎和根样品,用锡纸包裹,迅速放置液氮中,-80 ℃保存。
挑选健康、同批收获的‘华春6 号’种子,放在吸水饱满的蛭石上萌发3~4 d,至芽生长至2~3 cm时,从蛭石中取出,用清水洗净根部,固定在漂浮板上,让根部完全浸泡在pH 为4.3 的0.5 mmol/L 简单钙离子溶液中,预培养24 h;在该培养液基础上,梯度设置AlCl3浓度为0、25、50、75 和 100 μmol/L,培养24 h。在pH 为4.3 的简单钙离子溶液且AlCl3浓度都为50 μmol/L 的基础上,设置0、4、8、12、16、24、48、36 和72 h 的时间梯度。选取预处理后根部长势一致的材料进行处理;上述材料处理完后,取大豆根部样品,用锡纸包裹后迅速放入液氮中,-80 ℃保存。
使用RNA 提取试剂盒(南京诺唯赞生物科技股份有限公司)分别提取各组织和处理根部RNA,统一浓度后完成反转录,将反转录后的cDNA 放于-20 ℃保存,作为RT-qPCR 和载体的模板。
1.4 RT-qPCR 分析
以各处理cDNA 为模板,试剂使用SYBR 荧光定量酶(苏州近岸蛋白质科技股份有限公司),仪器使用CFX96 实时荧光定量PCR 仪(Bio-Rad),PCR 程序设置为95 ℃ 3 min;95 ℃ 10 s、58 ℃10 s、95 ℃ 30 s,39 个循环;95 ℃ 30 s。相关引物序列见表1,以β-actin为内参基因。目的基因的相对表达量用2-ΔΔCt法计算。
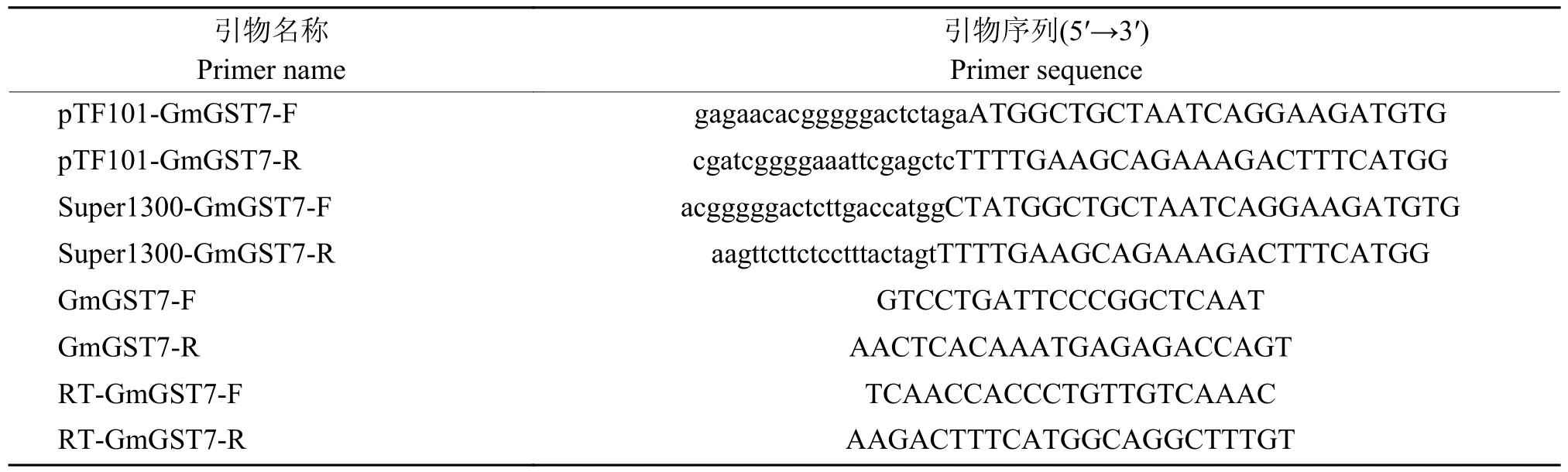
表1 引物序列Table 1 Primer sequence
1.5 载体构建
以‘华春6 号’根部的cDNA 为模板,用GmGST7的克隆引物(GmGST7-F/R)进行PCR 扩增。50 μL PCR 反应体系为:2×Phanta Max Buffer 25 μL、dNTP Mix 1 μL、正反向引物各2 μL、Phanta Max Super-Fidelity DNA Polymerase 1 μL、模板cDNA 2 μL、ddH2O 17 μL。PCR 反应程序为95 ℃预变性3 min;95 ℃变性15 s、57 ℃退火15 s、72 ℃延伸40 s,34 个循环;72 ℃保温5 min。
构建GmGST7过表达载体GmGST7-pTF101。使用XbaⅠ和SacⅠ对pTF101 进行双酶切,通过重组引物(pTF101-GmGST7-F/R)将GmGST7基因cDNA 序列连入pTF101,测序无误后转化农杆菌GV3101。
构建亚细胞定位载体GmGST7-Super1300,将GmGST7基因连接Super1300 亚细胞定位载体,使用XbaⅠ和SpeⅠ酶对Super1300 进行双酶切,通过重组引物(S u p e r 1 3 0 0-G m G S T 7-F/R) 将GmGST7基因cDNA 连到载体,测序无误后转化农杆菌GV3101。引物序列见表1。
1.6 亚细胞定位
将含有质粒GmGST7-Super1300 的GV3101农杆菌和Super1300 空载体的GV3101 农杆菌扩摇至D600nm=0.6,离心重悬。选择生长4~5 周、健壮的烟草,侵染前强白光照射2~3 h,使叶片气孔充分张开。从叶片下表皮注射菌液,注射完成后密封保湿、暗处理1 d,正常培养2 d 后,使用激光共聚焦显微镜(德国徕卡,激光共聚焦TCS-SP8 SR)观察,叶绿体荧光信号激发波长640 nm,检测波长675 nm,绿色荧光蛋白GFP 激发波长488 nm,检测波长510 nm。
1.7 拟南芥遗传转化
在营养土中种植野生型拟南芥。第1 次抽苔时剪去主苔,在侧苔快要开花时准备侵染,如第1 次侵染有结荚,则去除。侵染每周1 次,共侵染3 次。将载体GmGST7-pTF101 转化GV3101,鉴定阳性菌后扩摇至D600nm=0.6,离心后用重悬液重悬后准备侵染。将拟南芥花浸泡1 min 左右,密封保湿、暗培养1 d 后,正常培养。
1.8 拟南芥酸铝处理及表型鉴定
在营养土中种植侵染后收获的拟南芥种子,2 片真叶完全长开后定期多次喷洒除草剂,去除假阳性苗。待植株生长健壮后选取鲜嫩的叶片,用TPS 抽提液快提DNA,设计包含35S 启动子区域的引物进行DNA 分子水平鉴定。以此方法鉴定并加代至T3代。提取不同株系拟南芥RNA,通过荧光定量鉴定,选择表达量高的株系大量繁种。
将拟南芥种子用10%(φ)次氯酸钠溶液浸泡震荡10 min 消毒,然后用无菌水清洗5 遍。种植于1/2 MS 培养基上,将播种后的培养基放于4 ℃冰箱春化3 d,然后转移至培养室竖直培养,待根长至1 cm 左右,进行铝处理。
设置0、50、100 μmol/L AlCl3浓度梯度的1/2 MS 培养基,pH 调至4.3。将长势一致的拟南芥转移至该培养基上,保持根的伸长状态。竖直培养1 周左右,拍照记录,测量根长,计算根相对伸长。提取RNA,对拟南芥中酸铝胁迫响应的标志基因和GmGST7同源基因At2G29420的下游基因进行荧光定量分析。酸铝标志基因为铝激活苹果酸盐转运体AtALMT(At1G08430)[27];AtMATE(At1G51340)[27]参与铝离子外排;AtALS3(At2G37330)[27]编码ABC 转运蛋白,参与铝离子在体内的运转,减少铝对根的伤害;AtWAK1(At1G21250)[28]是一种细胞质相关激酶,参与细胞壁的修复。通过Phytozome 搜索At2G29420基因下游基因,得到3 个谷氨酰胺水解酶基因(At1G78660、At1G78670、At1G78680) 和1 个氨基肽酶基因(At4G33090)。引物见表2。测定植株中MDA 含量、H2O2含量、POD 活性(参考科铭生物试剂盒说明书)。

表2 荧光定量引物序列Table 2 qPCR primer sequence
2 结果与分析
2.1 GmGST7 蛋白保守结构域
GmGST7基因在大豆第7 号染色体上(基因登录号码为LOC547578),该基因周围存在基因簇。GmGST7基因含有2 个外显子、1 个内含子;基因组全长1 128 bp,转录本986 bp,cDNA 全长678 bp,编码226 个氨基酸。GmGST7 编码典型的tau 类GST 蛋白,含有2 个GST 保守结构域,分别为高度保守的N 端和不保守的C 端,如图1 所示。N 端三维结构的组成是β1α1β2(α2)β3β4α3,C 端是一个完全α-螺旋结构。

图1 GmGST7 蛋白结构域Fig. 1 Protein domain of GmGST7
2.2 GST tau 类家族进化树构建
GST 家族中tau 类在植物物种之间的差异较为明显。选择拟南芥和大豆中同源基因进行蛋白序列比对,在NCBI 中找到相似度最高的17 个基因进行蛋白序列比对,N 端和C 端序列相似度都较高,总体序列相似度为80%~99%。在不同物种中,只有N 端相对保守,C 端差异较大,总体序列相似度为44%~52%,如图2A 所示。在构建进化树时,大豆同源基因之间的遗传距离相对较近,和拟南芥的同源基因间的遗传距离相对较远,如图2B 所示。
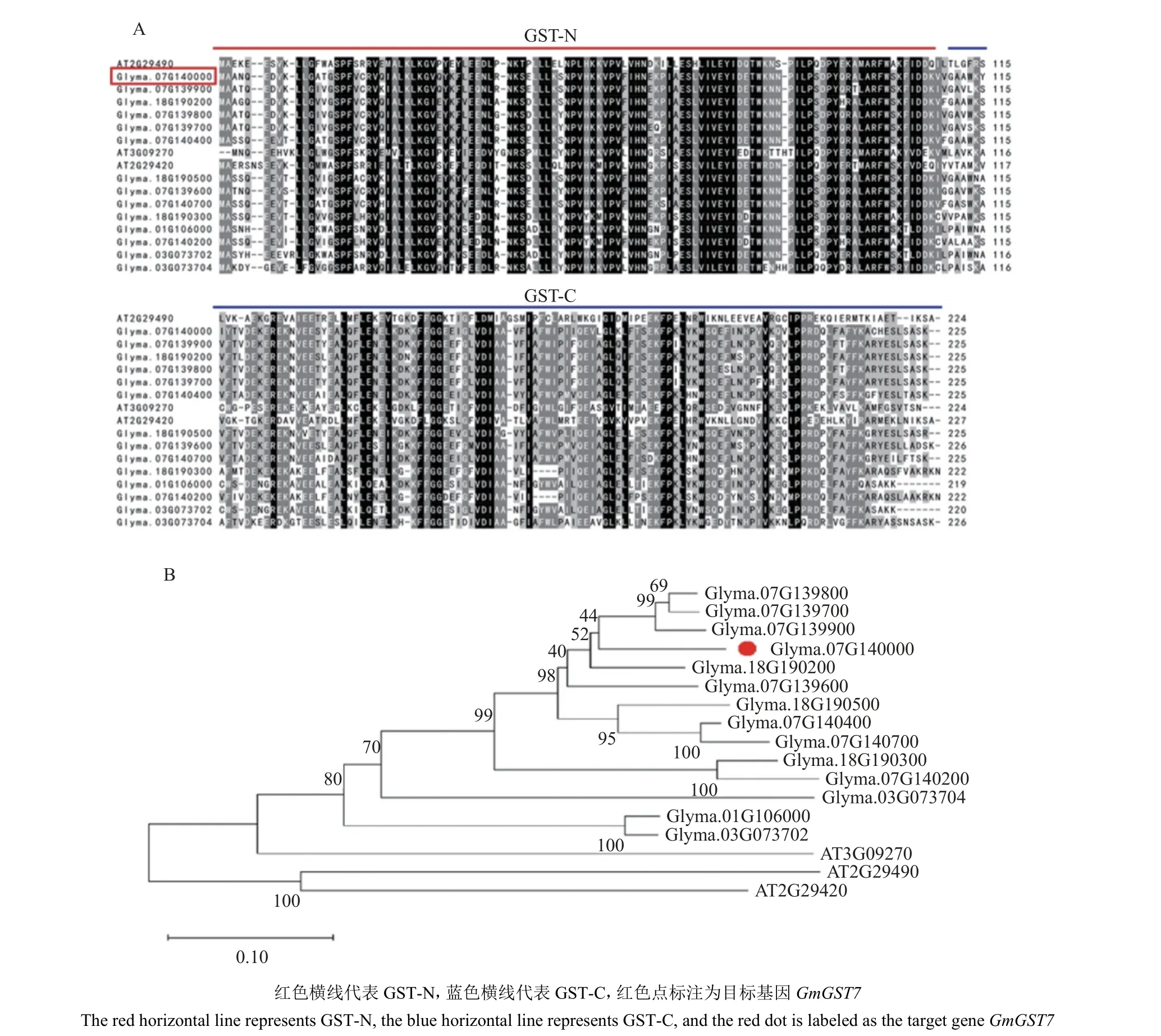
图2 GmGST7 蛋白序列对比(A)和进化树(B)分析Fig. 2 Sequence comparison (A) and evolutionary tree (B) analysis of GmGST7 protein
2.3 GmGST7 互作蛋白预测
通过STRING 网站对GmGST7 的互作蛋白进行预测,发现有5 个候选蛋白可能与GmGST7(LOC547578) 互作(图3)。其中,LOC732557(GLYMA07G16850)和DHAR1(GLYMA10G43730)属于GST 家族蛋白,和GmGST7 属于同一个家族。GLYMA03G40050 和HGS(GLYMA19G42610)属于HGS(谷胱甘肽合成酶)家族蛋白。

图3 GmGST7 互作蛋白预测Fig. 3 Interaction protein prediction of GmGST7
2.4 GmGST7 基因组织表达模式
为了检测GmGST7基因的组织表达模式,采集‘华春6 号’结荚初期的根、茎、叶、花、幼荚样品,定量检测结果(图4)表明,GmGST7基因呈组成型表达模式,GmGST7基因在叶、花和幼荚中的表达量相对偏低,在根中的表达量明显高于在其他组织中的;以幼荚的表达量为对照,GmGST7基因在根中的表达量是幼荚中的171.8 倍,是茎中的8.6 倍。

图4 GmGST7 基因的组织表达模式分析Fig. 4 Analysis of tissue expression pattern of the GmGST7 gene
2.5 GmGST7 基因酸铝胁迫响应
为了进一步分析GmGST7在铝胁迫下的表达特征,对GmGST7基因进行不同浓度梯度和时间梯度的表达分析。定量结果表明,随AlCl3浓度的增加,GmGST7的相对表达量先升高后降低,在50 μmol/L AlCl3溶液中的相对表达量最高,是对照的14.5 倍(图5A)。在时间梯度处理中,随着处理时间的推移,GmGST7基因表达量先升高后降低,在处理24 h 时表达量最高,为对照的24.9 倍(图5B)。
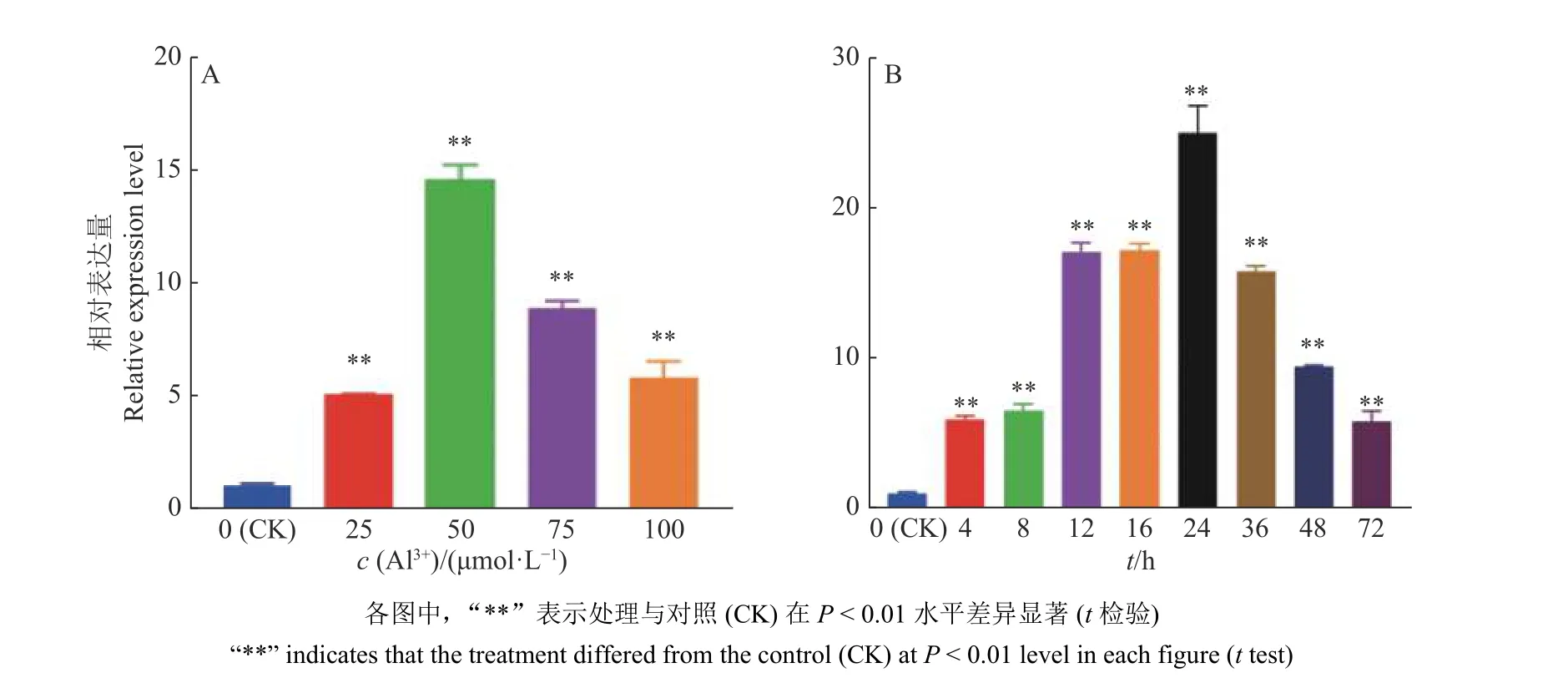
图5 GmGST7 基因酸铝胁迫响应Fig. 5 Response of the GmGST7 gene to acidic aluminum stress
2.6 GmGST7 基因载体构建
以‘华春6 号’转录组为模板,设计特异引物,PCR 扩增包含GmGST7整个cDNA 序列的产物。PCR 后通过琼脂糖凝胶电泳,获得大小为742 bp 的DNA 片段(图6A),符合引物设计产物长度。将PCR 产物纯化后进行测序,测序序列与NCBI 数据库中目标基因的序列一致。
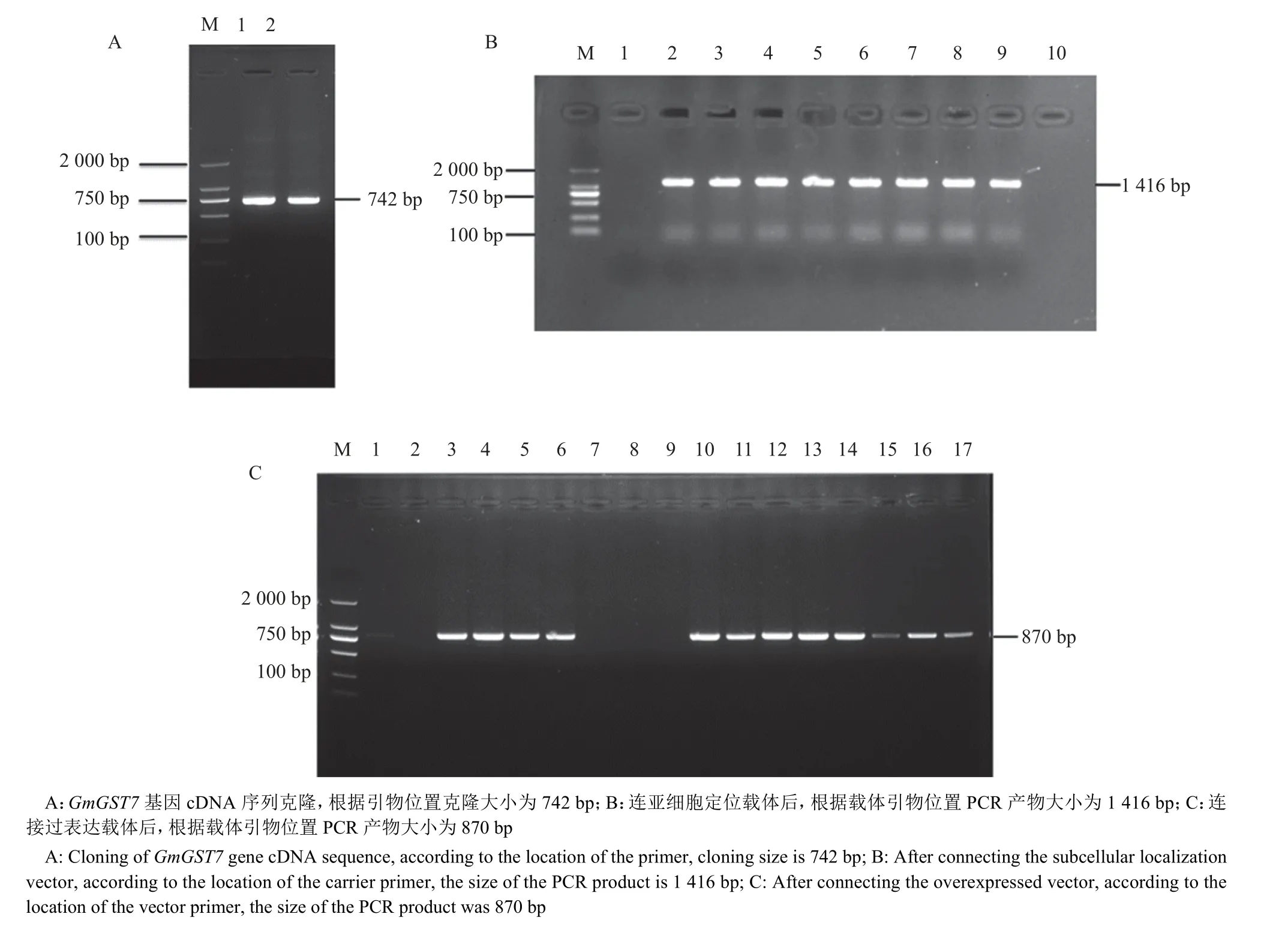
图6 GmGST7 基因克隆及载体构建Fig. 6 Cloning and vector construction of the GmGST7 gene
为验证亚细胞定位载体转化农杆菌GV3101 是否成功,选取培养皿上单菌落,扩摇至浑浊后用载体上的鉴定引物进行PCR 菌液鉴定,琼脂糖凝胶电泳结果(图6B) 显示具有预期大小的目的条带,DNA 片段大小为1 416 bp,选择阳性克隆测序,将测序序列与GmGST7基因cDNA 序列进行比对,全cDNA 序列都存在,且序列一致。
为验证过表达载体转化农杆菌GV3101 是否成功,选取培养皿上单菌落,扩摇至浑浊后用载体上的鉴定引物进行PCR 菌液鉴定,琼脂糖凝胶电泳结果(图6C)显示鉴定到预期大小的目的条带,片段大小为870 bp,表明这些菌落为阳性克隆。随后,挑选阳性克隆进行目的片段测序验证,将测序序列与GmGST7基因cDNA 序列进行比对,全cDNA序列都存在,且序列一致。
2.7 GmGST7 蛋白亚细胞定位
利用烟草研究GmGST7 蛋白的细胞定位,将GmGST7-GFP 和GFP 载体转化烟草,荧光检测结果(图7)表明,转化GFP 载体的细胞只有细胞核和细胞膜有荧光;在转化GmGST7-GFP 载体的细胞中,在细胞核和细胞质都存在荧光信号,由于细胞中央大液泡的存在,细胞质的荧光信号被挤压至细胞膜周围,且不与叶绿体信号重叠。荧光检测结果表明,GmGST7基因的蛋白定位于细胞质和细胞核,与网站(http://www.csbio.sjtu.edu.cn/bioinf/ Cell-PLoc-2/)预测的结果一致。

图7 GmGST7 蛋白亚细胞定位分析Fig. 7 Subcellular localization analysis of GmGST7 protein
2.8 拟南芥遗传转化鉴定
用除草剂喷施拟南芥,非转基因植株枯萎,GmGST7转基因阳性苗则正常生长(图8)。利用载体引物和GmGST7基因定量引物组成2 对引物,双向鉴定,PCR 产物经凝胶电泳检测后分别出现204 和753 bp 的目的条带(图9),通过该方法检测转基因阳性苗直至T3代。在T3代进一步通过定量PCR 检测转基因阳性株系,获得表达量较高的3 个株系(L5、L13、L14)(图10)。

图8 GmGST7 转基因拟南芥阳性苗除草剂喷洒鉴定Fig. 8 Herbicide spraying identification of GmGST7 transgenic Arabidopsis positive seedlings
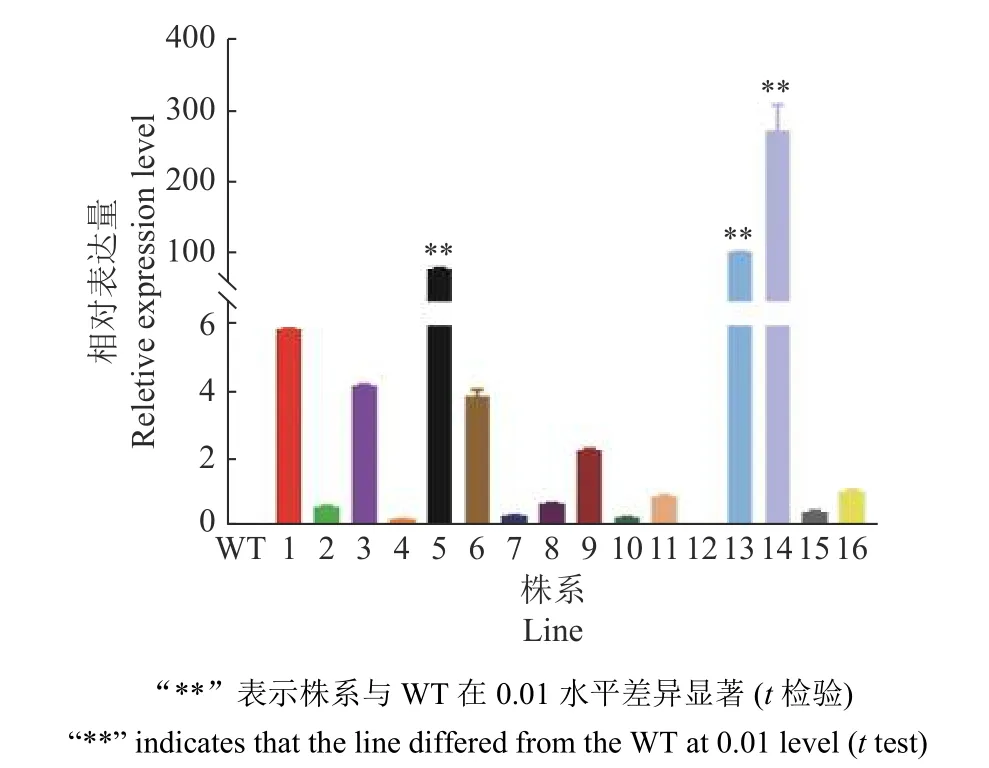
图10 转基因拟南芥T3 代株系中GmGST7 基因表达量Fig. 10 Expression of GmGST7 gene in T3 transgenic Arabidopsis thaliana
2.9 拟南芥耐酸铝表型鉴定
以WT 为对照,选择L5、L13、L14 3 个株系,设置3 个AlCl3浓度梯度进行耐酸铝表型鉴定。结果(图11)表明,随着AlCl3浓度的增加,拟南芥根系伸长受到抑制,野生型受到的抑制更加明显。在50 μmol/L AlCl3浓度下,GmGST7转基因株系的相对根伸长高于60%,在100 μmol/L AlCl3浓度下,相对根伸长均高于50%,均与野生型差异极显著。

图11 过表达GmGST7 拟南芥耐酸铝表型Fig. 11 Overexpressed GmGST7 Arabidopsis aluminiumresistant phenotype
在1/2 MS 培养基上,H2O2、MDA 含量和POD活性在野生型和转基因拟南芥中无显著差异。在酸铝培养基上,H2O2和MDA 含量在野生型和转基因拟南芥中均显著升高,其中,野生型H2O2含量增加幅度明显大于转基因株系,为对照处理的1.69 倍;转基因株系L5、L13、L14 H2O2含量分别为对照处理的1.29、1.38、1.23 倍。野生型MDA 含量比对照处理增加了1.80 倍,转基因株系比对照处理分别增加了1.30、1.23、1.21 倍。在酸铝培养基上,野生型POD 活性比对照处理增加了1.10 倍,3 个转基因株系比对照处理分别增加了1.22、1.19、1.37 倍。因此,在过表达GmGST7后,转基因拟南芥体内的氧化损伤要弱于野生型,增强了对酸铝胁迫的耐受能力。
2.10 拟南芥耐酸铝分子调控分析
利用酸铝胁迫响应的标志基因分析GmGST7转基因拟南芥耐酸铝的调控机制。在过表达GmGST7拟南芥中,酸铝条件下AtALMT、AtMATE、AtALS3表达量都有提高,分别为对照的1.36、1.33、1.34 倍,AtWAK1表达量提高最多,是对照的2.5倍。因此,推测GmGST7可能通过清除植株内有毒物质、降低植物体内的氧化水平、促进各耐酸铝基因的表达,进而加强植株在逆境下的耐受能力。
2.11 GST 下游基因的表达分析
使用NCBI 数据库,对GmGST7进行蛋白比对,在拟南芥中得到序列相似度最高的基因At2G29420。通过P h y t o z o m e 数据库搜索At2G29420,得到该基因下游主要有2 个家族,分别是谷氨酰胺水解酶(At1G78660、At1G78670、At1G78680)和氨基肽酶(At4G33090),这2 个蛋白在GST 催化谷胱甘肽和毒性物质结合后,参与下一步的分解工作。荧光定量分析结果表明,这4 个基因受酸铝胁迫诱导上调,并受GmGST7激活表达。在酸铝胁迫下,At1G78660、At1G78670、At1G78680、At4G33090在转基因株系中的表达量为野生型的1.71、1.45、1.44、1.26 倍。
3 讨论与结论
GST 是一个多功能的家族,是由硫氧化还原蛋白进化而来,目前已知的功能包括解毒、代谢、运输、隔离内源或者外源有毒成分等[29-30];对许多非生物胁迫产生应激反应,包括盐碱胁迫、低温高温胁迫、干旱胁迫[31-34],也对许多生物胁迫产生应激反应,如病原体攻击、重金属胁迫、过氧化物胁迫、紫外辐射等[35-38];参与催化或修饰各种酶促反应和小分子物质,包括谷胱甘肽(GSH) 偶联、过氧化物酶、巯基转移酶、谷胱甘肽酰化和脱氢抗坏血酸还原酶等[39-42]。
GST tau 类蛋白有2 个结合域,N 端序列高度保守,负责GSH 的识别和结合。C 端序列不保守,负责各种有毒物质或者植物体内一些信号的识别和结合[43-44];C 端在同物种间高度保守,不同物种间的氨基酸序列差异明显,当面对胁迫环境时,C 端会发生有利或有害突变,发生有利突变后,会增加其在基因组序列的高度重复,进而解释tau 类GST 在植物中的成簇出现和不同物种之间的差异[45-47]。在构建进化树时,大豆和拟南芥有许多相似的同源基因,由于C 端序列差异较大,拟南芥和大豆遗传距离相隔较远,而在大豆中的同源基因遗传距离较近。
GmGST7基因翻译典型的GST 家族tau 类蛋白,有2 个外显子分别是N 端和C 端。通过对GmGST7蛋白的氨基酸序列进行分析,发现N 端含有Ser 羟基,为GSH 巯基的氢键供体,对GSH 的催化激活起重要作用。C 端能促进高活性硫酸阴离子的形成和稳定,为亲电底物亲核攻击提供目标,这些亲电底物是植物在逆境胁迫中产生的过氧化物、活性氧等一些有毒物质。所以GmGST7 蛋白拥有GST 家族tau 类蛋白的典型功能,即能够促进氧化物与GSH 的结合,进而降低植物的氧化水平,这与过表达拟南芥的表型一致。在50 和100 μmol/L AlCl3处理下,GmGST7转基因株系比野生型的相对根伸长要高出50%。植株的氧化水平的测量结果也得出相同的结论,转基因株系的氧化水平要显著低于野生型,表现在H2O2和MDA 含量增幅低于野生型,POD 活性增幅低于野生型。当GmGST7基因过表达遗传转化拟南芥后,植株表现出酸铝胁迫,植株整体抗逆能力提高,相关下游基因表达量提高,氧化水平降低,进而减轻酸铝胁迫对植物生理功能的抑制。
许多研究都发现GST 家族基因响应逆境胁迫,并有相关研究显示通过遗传转化的手段能提高植物对逆境的耐受能力。如通过遗传转化野生大豆中GsGST14提高栽培大豆的盐碱胁迫能力;向日葵中HaGSTU1对核盘菌的侵染响应[48];在大豆中过表达GmGST12降低雄性不育大豆小孢子中的活性氧,提高其存活率[49];玉米中ZmGST23对干旱、高温、低温盐碱胁迫的响应[50];ZmGST31在玉米中对除草剂的解毒作用[51]。
研究表明,GST 家族基因在植物抵抗逆境胁迫时发挥了重要作用,大量关于GST 家族基因对作物品种改良的研究都取得了成功,因此,对GmGST7的深入研究将有助于解析大豆对酸铝胁迫的响应机制,为大豆抗逆分子育种提供理论依据和基因资源。
综上所述,GmGST7基因位于大豆第7 号染色体上,该基因含有2 个外显子和1 个内含子,2 个外显子分别编码GST 高度保守的N 端和不保守的C 端。GmGST7 蛋白定位于细胞质和细胞核中;GmGST7基因受酸铝胁迫诱导表达上调且具有时空表达模式;GmGST7基因呈现组成型表达模式,在根中富集表达;GmGST7基因过表达拟南芥后,可降低酸铝胁迫后的氧化水平,酸铝胁迫响应相关基因表达增强,根伸长的抑制得到缓解,拟南芥对酸铝胁迫的耐受能力显著提高。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17
学与玩(2022年10期)2022-11-23
今日农业(2022年3期)2022-06-05
亚热带植物科学(2022年1期)2022-05-17
四川农业科技(2019年5期)2019-07-01
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
浙江柑橘(2016年1期)2016-03-11
创新科技(2015年1期)2015-12-24
