
常规籼稻品种的稻瘟病抗性基因及穗颈瘟抗性分析
2023-10-12 04:02吴子帅陈传华刘广林罗群昌覃孙骞朱其南
华南农业大学学报 2023年5期
吴子帅 ,李 虎,陈传华,刘广林 ,罗群昌,覃孙骞,朱其南
(1 广西农业科学院 水稻研究所/广西水稻遗传育种重点实验室, 广西 南宁 530007; 2 来宾市农业科学院, 广西 来宾 546100)
由稻瘟病菌Magnaportheoryzae引起的真菌病害稻瘟病是水稻重要病害之一,一般情况下发病会造成10%~20%的减产,流行年份可造成50%以上减产,严重影响水稻产量和品质,被称为“水稻癌症”[1-2]。根据稻瘟病发生的时期和部位,可分为苗瘟、叶瘟、节瘟和穗颈瘟等,其中穗颈瘟对水稻生产的危害最大,穗颈瘟一般发生在抽穗破口前,病菌侵入穗颈部并逐渐蔓延,阻断养分通道,严重威胁产量甚至造成绝收[3-4]。通过大量使用化学农药防治稻瘟病,不仅成本高,还易造成环境污染和农药残留超标,不利于生态环境和食品安全[5],且应用较广的三环唑、春雷霉素等均为预防性药剂,对防治窗口期短的穗颈瘟难以取得良好效果[6],聚合多个稻瘟病抗性基因、培育高抗品种是公认防控稻瘟病最经济、环保、有效的方式[7]。挖掘抗性基因是开展多基因聚合育种的重要前提,目前已鉴定的稻瘟病抗性基因超100 个,已克隆的有39 个[8],这为抗性基因的检测和利用提供了重要保障。抗病品种的选育依赖于抗病资源的发掘与利用。李刚等[9]鉴定了544 份水稻种质资源的稻瘟病抗性水平及其携带的主效抗性基因,指出品种的抗性与其抗性基因种类密切相关,Pi5、Pita、Pi9和Pib对其研究所用的6 个强致病小种抗性表现较好;朱业宝等[10]对156 份外引水稻种质资源进行田间自然诱发鉴定和稻瘟病抗性基因检测,结果表明携带Pi9和Pi2的水稻资源的综合抗性较好,“Pi9+Pi5+Pik-m+Pia”、“Pi5+Pib+Pita+Pik-m+Pia”和“Pi2+Pi54+Pib+Pita+Pik-m+Pia”组合的稻瘟病综合抗性较好;王小秋等[11]分析了稻瘟病抗性基因在195 个粳稻新品种/系中的分布情况,指出供试品种对穗颈瘟的抗性主要与Pia、Pi5和Pita显著相关,抗性基因组合“Pia+Pita”在江苏粳稻抗穗颈瘟育种中有重要应用价值;此外,周坤能等[12]、陈晴晴等[13]、王晓玲等[14]、潘争艳等[15]分别对153 份安徽粳稻、252份长江中下游区试品种、82 份江西粳籼骨干亲本、260份辽宁粳稻进行稻瘟病抗性基因检测及抗性鉴定,这些研究结果为各地抗性亲本的选择和品种的合理布局提供了良好的理论参考。但目前针对两广籼稻区优质常规稻育种骨干亲本的稻瘟病抗性基因及穗颈瘟抗性分析的研究鲜见报道。本研究以121份广西、广东历年审定的常规籼稻为材料,进行田间穗颈瘟抗性鉴定及Pi2、Pi5、Pi9、Pi33、Pi46、Pi54、Pib、Pid3、Pigm、Pit、Pita、Pia、Pik-m和Pik等14个稻瘟病抗性基因的分子鉴定,分析它们的基因型与抗性的关系,旨在为抗性基因聚合育种的亲本选择提供支持,为常规稻的合理布局提供理论参考。
1 材料与方法
1.1 试验材料
试验材料为121 份广西农业科学院水稻研究所优质常规稻育种研究室收集保存的常规籼稻,包括75 份2000—2021 年广西审定品种和46 份1999—2020 年广东审定品种。这些材料一方面曾作为华南稻区的主栽或主推品种,另一方面也是目前广西优质稻育种的骨干亲本。‘桂育6 号’为广西水稻品种区试稻瘟病抗性鉴定所用的诱发材料。
1.2 试验方法
1.2.1 稻瘟病抗性基因检测 采用五引物扩增受阻突变体系(Penta-primer amplification refractory mutation system,PARMS) SNP 分型技术进行Pi2、Pi5、Pi9、Pi33、Pi46、Pi54、Pib、Pid3、Pigm、Pit、Pita、Pia、Pik-m和Pik等14 个稻瘟病抗性基因的检测,DNA 提取采用常规的CTAB 法,PCR 反应体系10 μL,35 个循环,分子特异性标记引物由武汉市景肽生物科技有限公司提供。PCR 完成后,以FAM和HEX 作为报告荧光,ROX 作为参比荧光,用TECAN infinite F200 酶标仪读取荧光信号,解析转换荧光信号得到清晰直观的分型图,并根据颜色不同,输出基因型结果[5,16]。具体检测委托武汉市景肽生物科技有限公司进行。
1.2.2 稻瘟病田间抗性鉴定 自然诱发鉴定于2022 年早稻在广西壮族自治区来宾市金秀瑶族自治县罗香乡琼伍村旱田屯(110.09°E、24.39°N)进行,该试验地处山谷丘陵,海拔270 m,雾大露重,稻分蘖盛期日平均气温为20~25 ℃,相对湿度为90%以上,具有诱发稻瘟病的有利环境条件。每份材料种植1 个小区,每小区10 行,每行4 株,小区两边各插1 列诱发材料,株行距20.0 cm×13.0 cm,小区随机排列,重复3 次。除不进行稻瘟病防治外,田间其他管理同一般水田。于黄熟初期按《水稻品种试验稻瘟病抗性鉴定与评价技术规程(NY/T2646—2014)》[17]的方法调查穗颈瘟,并按该标准中的“水稻穗颈瘟单穗损失率0~9 级分级标准”和“水稻穗颈瘟发病率群体抗性分级标准”对材料进行穗颈瘟抗性分级,以3 次重复中感病最严重的程度作为鉴定结果。
1.3 统计分析
采用办公软件WPS 对研究中获得的各类数据进行整理和作图。以DPS v9.01 进行统计分析,采用LSD 法检测差异显著性。
采用SPSS statistics 26 进行逻辑回归分析[11],将供试材料田间穗颈瘟抗级≤3 的定义为1, > 3 的定义为0,携带某抗性基因定义为1,不携带该抗性基因定义为0,得到数据表后对单个基因进行逻辑回归分析及卡方检测,获得各基因与抗性间的回归系数(B)、显著性检验的P值、优势比(Odds ratio,OR)和95% 置信区间(Confidence interval,CI)。
2 结果与分析
2.1 稻瘟病抗性基因检测
利用PARMS SNP 分型技术对稻瘟病抗性基因Pi2、Pi5、Pi9、Pi33、Pi46、Pi54、Pib、Pid3、Pigm、Pit、Pita、Pia、Pik-m和Pik的检测结果(图1)表明,所有供试品种均不携带Pi9、Pigm、Pik-m和Pik,其余10 个抗性基因则有不同程度的分布;供试品种中,Pi46和Pia的检出率较低,分别为3.3%和7.4%,Pi54检出率高达86.0%,Pi5检出率为67.8%,其他基因检出率在18%~46%之间;就品种而言(图2),携带了2~5 个基因的品种占总数的86.0%,携带1 个基因的品种有7 份,均为广西品种,携带7 个基因的品种有2 份,均为广东品种。以上结果表明,供试品种在携带已知稻瘟病抗性基因上存在明显差异,Pi54和Pi5的应用较广,Pi9、Pigm、Pik-m和Pik在供试品种中尚没有被利用。

图1 抗性基因在供试品种中的分布频率Fig. 1 Distribution frequency of resistance genes in tested varieties
2.2 稻瘟病抗性基因在不同类型供试品种中的应用情况
为分析不同地域环境、审定年份的品种间携带的抗性基因是否存在差异,比较除供试品种都不携带的Pi9、Pigm、Pik-m和Pik外其他抗性基因在不同类型供试品种中的分布,结果(表1)显示,2011年后的广西品种较2011 年前相比,Pi2、Pi5、Pid3和Pit出现频率分别提高22.7、22.8、18.2 和20.3 个百分点,特别是Pi5在2011 年后出现频率达84.1%,而Pi46和Pita出现频率分别降低12.9 和15.2 个百分点;2011 年后的广东品种较2011 年前相比,Pi2、Pi33、Pib和Pid3出现频率分别提高42.7、16.2、20.4 和35.4 个百分点,特别是Pi2在2011 年后出现频率达85.0%,而Pi5和Pita出现频率分别降低29.2 和22.7 个百分点;综合比较广西和广东品种间抗性基因分布频率,Pi2和Pid3在广西品种中出现频率比广东品种分别低47.6 和39.3 个百分点,而Pi5在广西品种中出现频率高出广东品种18.2 个百分点。以上结果表明,多数抗性基因在不同类型品种中应用基本相近,广东常规稻选育中主要是对抗性基因Pi2、Pi5、Pi54、Pid3和Pita的利用,广西常规稻选育中主要是对抗性基因Pi5、Pi54和Pita的利用,而2011 年后广西的常规稻选育中对Pi2和Pid3的利用也明显提升。

表1 不同类型供试品种间抗性基因分布频率Table 1 Distribution frequency of resistance genes in different types of tested varieties%
2.3 供试品种穗颈瘟抗性分析
穗颈瘟田间抗性调查结果(表2)显示,供试品种中没有高抗穗颈瘟的品种,中抗+抗的品种有25 份,占比20.6%,感+高感的比例高达到51.3%;广西的供试品种没有达高抗或抗水平的,中抗品种也仅占比10.7%,感+高感的比例高达到64.0%;广东的供试品种中抗+抗的品种有17 份,占比37.0%,感+高感的品种有14 份,占比30.4%。以上结果表明,供试品种的穗颈瘟抗性普遍较弱,相比之下,广东供试品种的穗颈瘟抗性总体明显好于广西供试品种。
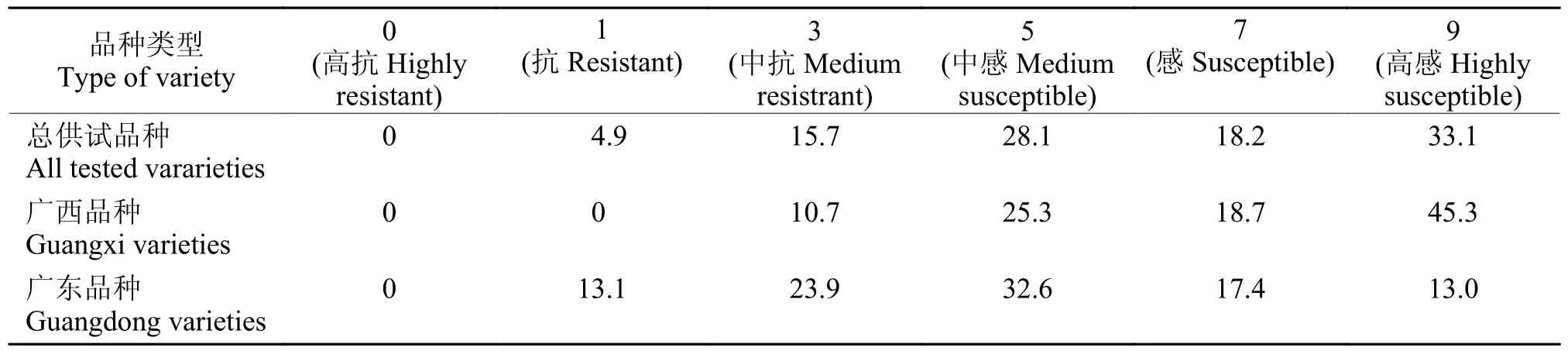
表2 供试品种穗颈瘟抗级分布频率Table 2 Distribution frequency of resistance level to panicle neck blast in tested varieties%
2.4 携带的抗性基因数量与穗颈瘟抗性相关分析
供试品种携带的抗性基因数量与抗级间的相关分析显示(图3a),二者间的决定系数R2=0.439 3,未达统计显著水平(P=0.105),表明抗性基因数量与穗颈瘟抗性间的相关性不显著。进一步比较抗的(抗级≤3)和感的(抗级≥5)供试品种间携带的抗性基因数差异(图3b),发现2 类品种携带的平均抗性基因数分别为3.92 和3.38,差异不显著(P=0.085)。同时在供试品种中也发现携带的抗性基因数量少却能达到中抗及以上水平的品种,及携带抗性基因数量多却表现中感以下的品种。因此对品种的穗颈瘟抗性改良,除简单的抗性基因聚合外,还应考虑不同遗传背景下抗性基因的强弱及组合效应。
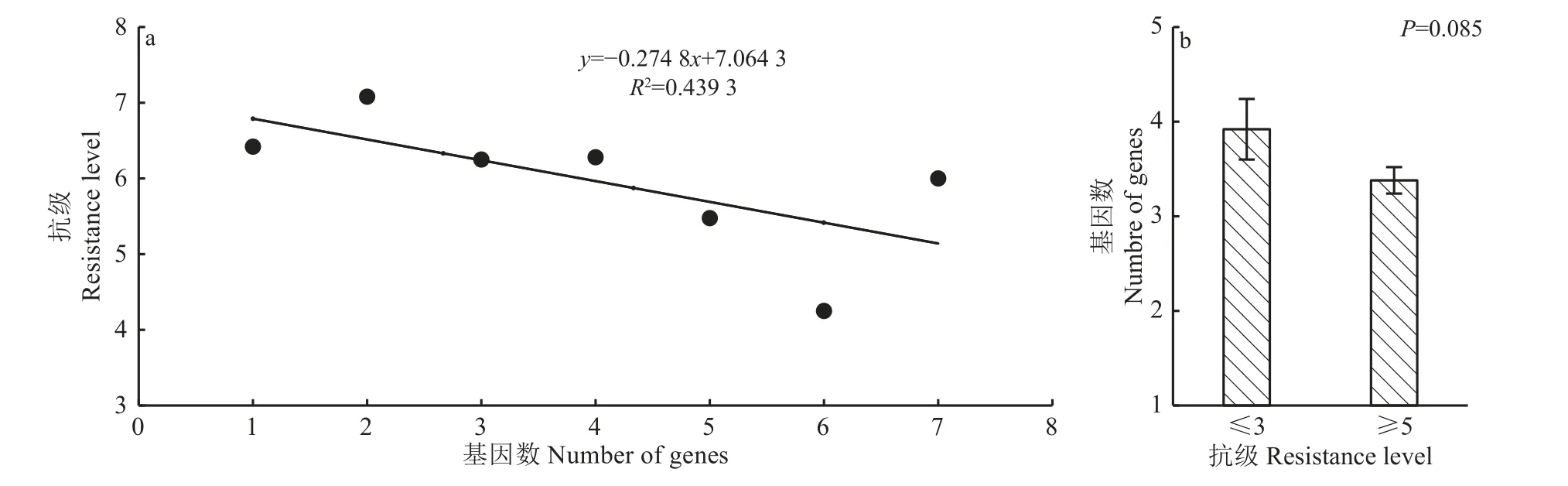
图3 品种携带抗性基因数量与抗级间的关系分析Fig. 3 Analysis of the resistance gene number carried by each variety and resistance level
2.5 稻瘟病抗性基因对穗颈瘟抗性贡献分析
采用逻辑回归法分析各基因对穗颈瘟抗性的贡献,结果见图4a,Pi2与穗颈瘟抗性间的回归系数(B)为1.79,达到极显著(P=0.00),Pid3与穗颈瘟抗性间的B为2.01,达到极显著(P=0.00),表明在单基因水平上Pi2和Pid3对抗性有显著贡献;Pi2和Pid3的优势比(OR)分别为5.98 和7.50,表明携带Pi2、Pid3对抗性的贡献是不携带Pi2、Pid3的5.98 与7.50 倍。进一步分析(图4b) 显示,携带Pi2(Pi2+) 的品种平均抗级为4.58,不携带Pi2(Pi2-)的品种平均抗级为6.90,差异极显著,携带Pid3(Pid3+)的品种平均抗级为4.35,不携带Pid3(Pid3-)的品种平均抗级为6.80,差异极显著,而其他基因的携带与否,造成的抗级差异均不显著。
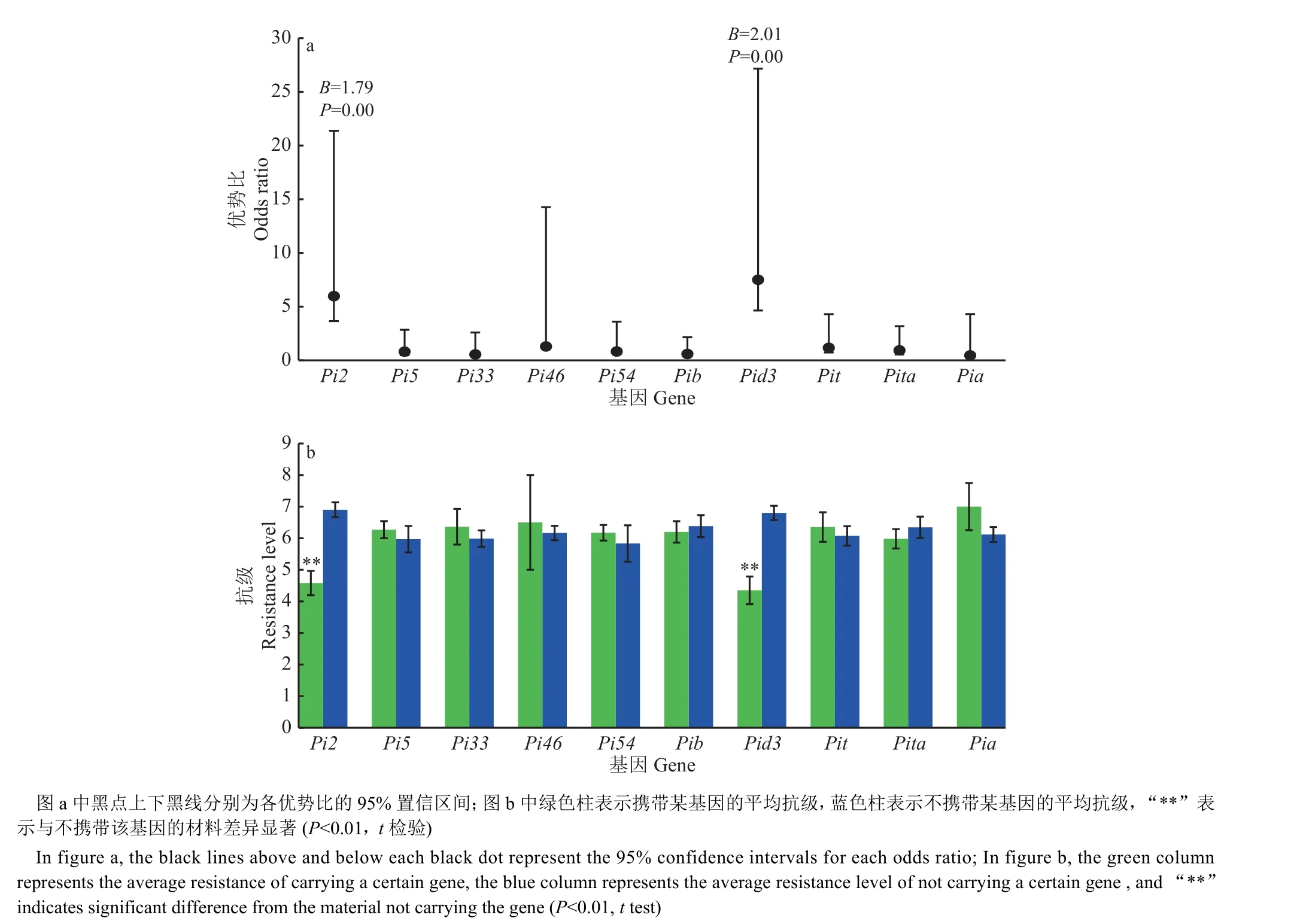
图4 单个抗性基因对穗颈瘟抗性贡献的逻辑回归分析(a)及平均抗级(b)Fig. 4 Logistic regression analysis of contribution to resistance (a) and average resistance (b) of single gene to panicle neck blast
2.6 稻瘟病抗性基因组合效应分析
分析抗性基因间的聚合叠加效应,结果(图5)显示,Pi2+Pid3+平均抗级4.35,Pi2+Pid3-的平均抗级5.57,Pi2+的平均抗级4.58,表明聚合Pi2和Pid3能增强抗病效应;Pi2+Pi33+平均抗级低于Pi2+Pi33-和Pi2+的平均抗级,Pid3+Pi33+平均抗级低于Pid3+Pi33-的平均抗级,表明在携带Pi2和Pid3的材料中聚合Pi33能一定程度上增强抗病效应。此外,5 个Pi2+Pi33+Pi54+的供试品种平均抗级3.0,田间穗颈瘟鉴定无感~高感的情况,1 个Pi2+Pi33+Pi54-的供试品种抗级9.0,表明Pi54也可能使Pi2+Pi33+的抗病效应增强。
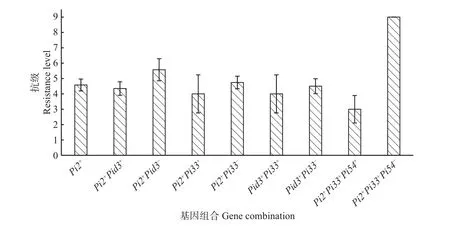
图5 不同稻瘟病抗性基因组合的抗级差异Fig. 5 Resistance level of different rice blast gene combinations
3 讨论与结论
稻瘟病生理小种复杂多变,不同的水稻种植区域优势小种构成往往不同,稻瘟病抗性基因在面对不同的生理小种时表现的抗性也不尽相同,培育广谱、持久抗性的水稻品种是防控稻瘟病的重要措施。而抗性基因鉴定及田间抗性分析,能较快掌握目标基因的分布状态和潜在的育种价值,是水稻育种资源鉴定的重要基础。
陆展华等[18]对广东省主栽品种的稻瘟病抗性基因及田间抗性鉴定表明,Pi2的检出率为17.6%,且抗性贡献显著,Pib和Pita的检出率虽高,但抗性贡献低;Pi9作为广谱抗性基因,对大多数国家的生理小种均表现出较好的抗性[19-20],但Pi9在国内育种上的应用差异较大,在东北稻区粳稻品种中检出率高达45.8%[21],而在广东籼稻主栽品种中未鉴定到[22],Tian 等[23]研究也认为Pi9还未广泛应用于我国籼稻品种。本研究结果发现,来源于两广地区的供试籼型常规稻品种均不携带Pi9,Pi2的检出率为31.4%,逻辑回归分析表明Pi2对穗颈瘟抗性贡献极显著,Pib和Pita的检出率分别为37.2%和45.5%,均对穗颈瘟抗性贡献不显著,本研究与上述研究结论是一致的,说明Pib和Pita对两广籼稻区的生产应用价值降低,应持续加强对Pi2的应用,并挖掘Pi9在两广籼稻区抗病品种选育及生产上的应用潜力。主效稻瘟病抗性基因Pik及其等位基因Pik-m对国内多个生理小种都有良好的抗性[24],Pik-m和Pik对广东、湖南、四川、江苏等稻瘟菌株的抗性频率达到90%以上[25],广谱抗性基因Pigm是一个与Pi2等位、抗谱差异显著的基因,对稻瘟病抗性改良有重要应用价值[26],本研究结果发现供试品种均不携带Pik-m、Pik和Pigm,暗示着两广籼稻区抗病品种选育及生产可尝试对Pik-m、Pik和Pigm的利用。Pid3是从‘谷梅2 号’中克隆的一个广谱稻瘟病抗性基因,在抗病品种改良中具有重要的利用价值[27],本研究结果表明,Pid3对穗颈瘟抗性贡献显著,供试品种中Pid3的平均检出率为25.6%,2011 年前后广西供试品种中Pid3的检出率从0 提高到18.2%,2011 年前后广东供试品种中Pid3的检出率从34.6%提高到70.0%,说明对Pid3在抗病育种中的重视和利用逐渐加强。
本研究结果发现,对穗颈瘟抗性贡献显著的Pi2和Pid3在广东品种中的应用明显高于广西品种,这是广东品种田间穗颈瘟抗性表现明显好于广西品种的重要原因之一。2011 年以前广西品种对Pi2和Pid3的利用为0,2011 年之后广西常规稻品种选育中加强了对这2 个基因的利用,这也是广西常规稻品种稻瘟病抗性提升的重要原因之一。
利用分子标记辅助选择是水稻新品种选育的高效手段,聚合多个稻瘟病抗性基因是改良品种抗性的常用方法,陈晴晴等[13]研究表明,聚合3 个抗性基因的穗颈瘟抗病率比1 个基因时上升54.3%,但更多的研究认为,携带的抗性基因数与稻瘟病抗性之间并不呈显著的负相关[11-12],抗性基因的聚合并非简单的抗谱叠加,应选择抗性好、抗谱互补的基因进行聚合,不仅能拓宽抗谱,还能提高对一些生理小种的抗性[28]。与前人研究结果一致,本研究结果表明,携带的抗性基因数量与穗颈瘟抗性间相关性不显著,对穗颈瘟的抗性改良,除简单的抗性基因聚合外,还应该充分考虑不同遗传背景下抗性基因的强弱及基因间的互作效应,而本研究对稻瘟病抗性基因组合效应分析发现,Pi2+Pid3+、Pi2+Pi33+和Pid3+Pi33+组合的穗颈瘟抗性表现较好。此外本研究中6 个Pi2+Pi33+Pi54+的供试品种平均抗级3.0,唯一一个Pi2+Pi33+Pi54-的供试品种抗级9.0,是否是Pi54的存在增强了Pi2+Pi33+的抗性,还需更多的样本支撑。
猜你喜欢
作物学报(2022年6期)2022-04-08
空间科学学报(2021年1期)2021-05-22
中成药(2018年12期)2018-12-29
环境保护与循环经济(2017年5期)2018-01-22
河南农业(2016年6期)2016-11-26
中国果菜(2016年9期)2016-03-01
现代农业(2016年5期)2016-02-28
中国蔬菜(2015年9期)2015-12-21
Asian Journal of Urology(2015年3期)2015-12-16
江西化工(2015年5期)2015-04-17
