
L-酪氨酸高密度流加发酵工艺研究
2023-10-10 12:54:00龚雨马零赵春光徐庆阳
中国调味品 2023年10期
龚雨,马零,赵春光,徐庆阳,3,4*
(1.天津科技大学 生物工程学院,天津 300457;2.宁夏伊品生物科技股份有限公司,银川750100;3.代谢控制发酵技术国家地方联合工程实验室,天津 300457;4.天津市氨基酸高效绿色制造工程实验室,天津 300457)
fermentation
L-酪氨酸(L-tyrosine,L-Tyr)是一种芳香族氨基酸[1],是人体必需氨基酸之一,对人体和动物的生长代谢都具有重要作用,有着广泛的市场需求[2],在食品行业中,因其具有极高的营养价值,被用作氨基酸类营养补充剂添加到食品中[3],L-酪氨酸及其衍生物还是许多保健品的重要成分[4];在医药方面,L-酪氨酸的衍生物广泛应用于医药载体,比如多巴胺是抗氧化药物合成的前体物质,用于多种类型休克药物的合成研究[5],L-酪氨酸亚硫酸盐合成的药物能治疗无力综合征、精神分裂症等疾病;在化工方面,能够降低黑色素的合成[6],是天然美白化妆品的合成原料。
当前L-酪氨酸生产主要以微生物发酵法为主,该方法具有成本低、无污染等生产优势,但其大量生产仍面临着巨大挑战[7]。在微生物大规模发酵生产过程中,菌体密度在一定程度上决定了目标产物的生成量[8],因此高密度培养无疑是提高产量的重要途径之一,其中培养基成分、发酵条件的控制等是至关重要的影响因素[9]。在大肠杆菌代谢途径中L-酪氨酸合成的前体是磷酸烯醇式丙酮酸(PEP)和4-磷酸赤藓糖[8],磷酸烯醇式丙酮酸还是芳香族氨基酸合成的前体物质[10-11],Mg2+是糖酵解途径中的能量传递者NADH合成过程的辅因子,是其合成的激活剂[12]。维生素H可以促进菌体生长,但过量会造成菌体生长过快进入衰亡期,过少又会导致生长过慢的问题[13];磷酸二氢大肠杆菌合成细胞膜磷脂提供了主要成分磷,还参与了大肠杆菌的生长代谢,控制其生长速率[14-15],直接影响了大肠杆菌的生长密度和重组蛋白的表达量[12],磷酸吡哆醛(PLP)广泛存在于细胞中,参与了脱羧反应、氨基转移、消除反应[16];在代谢过程中,维生素B12是许多酶的辅酶,参与了甲基化和化合物异构作用,对DNA和蛋白质的合成及细胞的生长也有着重要作用[17];甲硫氨酸能增大蛋白的可溶性表达量,减少包涵体蛋白的形成,使后续纯化工艺更简便[18];谷氨酰胺在细胞中充当氮的载体,还作为蛋白质合成的调节物,对细胞的生长具有重要作用[19]。
为促进高密度发酵过程中微生物细胞更好地生长,避免营养不足或浪费等问题,本研究对L-酪氨酸发酵过程中的生长因子进行优化,并且设计了3种不同的底物添加方式进行上罐生产,大幅度提高了菌体最高密度及L-酪氨酸产量。
1 材料与方法
1.1 菌种
本实验的供试菌株为大肠杆菌 TYR-05,保藏于天津科技大学代谢工程研究室。
1.2 培养基
1.2.1 活化培养基
葡萄糖25 g/L,酵母粉6 g/L,柠檬酸1 g/L,(NH4)2SO4·7H2O 2.4 mg/L,MgSO4·7H2O 0.4 g/L,谷氨酸 0.6 g/L,蛋氨酸0.5 g/L。
1.2.2 发酵培养基
葡萄糖25 g/L,酵母粉4.5 g/L,柠檬酸1.5 g/L,(NH4)2SO4·7H2O 2.8 mg/L,维生素H 0~2 mg/L;MgSO4·7H2O 0~3 g/L,KH2PO40~3 g/L;PLP 0~3 mg/L;维生素B120~3 mg/L。
1.3 仪器与设备
Biotech-15JS-3发酵罐 上海保兴生物工程设备有限公司;Aglient 1290高效液相色谱仪 美国安捷伦科技公司;SBA-40D生物传感分析仪 山东佰森泰克仪器有限公司;752PC紫外可见分光光度计 上海洪纪仪器设备有限公司。
1.4 培养方案
1.4.1 菌种活化
于-80 ℃冰箱中取出甘油管,使用接种环蘸取4环菌液涂抹于一代斜面上,37 ℃培养箱培养12 h,第2天再从一代斜面接种到二代斜面,继续37 ℃培养10~12 h。
1.4.2 摇瓶培养
以培养基总体积的10%为接种量置于摇瓶中,调节pH为7.0~7.2,温度37 ℃,220 r/min培养45 h(2%苯酚红作为指示剂,补加氨水调节pH)。
1.4.3 5 L发酵罐培养
1.4.3.1 发酵菌种活化
于-80 ℃冰箱中取出甘油管,使用接种环蘸取4环菌液涂抹于一代斜面上,于37 ℃培养箱中培养12 h,第2天再从一代斜面接种到二代斜面,继续于37 ℃培养10~12 h。
1.4.3.2 发酵罐种子培养
使用200 mL无菌水在超净台火焰旁洗脱茄形瓶中的菌体,在火焰旁从发酵罐进料口倒入种子培养基中。发酵过程中通过发酵罐参数将温度控制在37 ℃,pH控制在7.0~7.2,溶氧控制在45%~55%,生物量>18时接入发酵培养基,进行发酵培养。
1.5 方法
1.5.1 生长因子添加量优化单因素实验
在保证培养基等其他条件不变的前提下,按照表1中7种不同种类的生长因子,设计5组不同浓度梯度进行实验,实验结果取平均值。
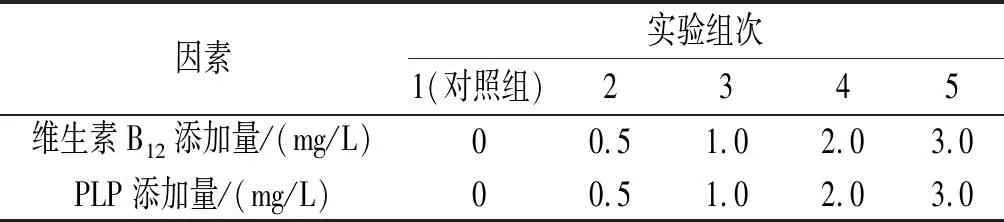
表1 生长因子单因素实验批次及添加量
1.5.2 正交实验确定生长因子的最适添加量
在实验2.1和2.2的基础上,综合菌体密度和产量考虑,各生长因子的影响顺序为KH2PO4添加量>维生素H添加量>MgSO4·7H2O添加量>维生素B12添加量>PLP添加量,其中甲硫氨酸和谷氨酰胺的影响非常小。因此本实验在单因素实验的基础上,以KH2PO4添加量(A)、PLP添加量(B)、维生素B12添加量(C)、维生素H添加量(D)和MgSO4·7H2O添加量(E)为研究对象,以菌体密度、L-酪氨酸产量为指标,进行正交实验进一步分析,正交实验因素水平表见表2。

表2 正交实验因素水平表
1.5.3 生长因子添加方式的测定
由2.4可知,在优化添加量的基础上,为了进一步达到实验预期,实验设计了3种生长因子添加方式,即一次性添加(a)、分时间段(10,15,20,25 h)添加(b)和10 h后持续流加(c)。3种添加方式添加总量一致 。
2 结果分析
2.1 生长因子添加种类及添加量对菌体密度的影响
由图1可知,不同生长因子对菌体生长有较大影响,其中KH2PO4和维生素H对菌体密度的影响较大,当KH2PO4添加量为2.5 g/L时,菌体密度值达到34.1 g/L,当维生素H添加量为1.0 mg/L时,菌体密度最高,为30.2 g/L;二者差距较小,并且均高于其他5种生长因子对应的菌体密度。菌体的生长及基因的表达很大程度上取决于磷酸盐的含量[20],细胞中磷酸盐的含量对大肠杆菌中质粒表达及复制的速率有一定影响,因而决定了菌体的蛋白表达量[9];磷酸盐是大肠杆菌合成细胞膜磷脂的主要成分,对菌体的生长有很大影响;它对代谢流分布有主要影响,控制着EMP途径中L-酪氨酸合成的关键节点,PEP(磷酸烯醇式丙酮酸)和E4P(4-磷酸赤藓糖)以及PRPP(磷酸核糖焦磷酸)是L-酪氨酸的合成原料[21],决定了EMP途径的碳流量,因此磷元素是大肠杆菌生产L-酪氨酸不可或缺的成分之一,当过量添加时,会使代谢流中前体物的供给达到一种失衡的状态,无法达到平衡点,进而使菌体密度降低[21];生物素参与了脂肪酸生物合成、氨基酸代谢和糖异生三大代谢过程[12],参与了脱羧、羧化、脱氢化等反应[12],是乙酰CoA羧化酶等的辅酶,还参与了CO2的固定作用,因此随着添加量的增加,菌体密度逐渐变大,但当菌体密度达到最高值30.2 g/L后呈下降趋势,分析得到添加量过高会导致菌体代谢过快而过早衰老,进而导致菌体密度下降。

图1 单一生长因子对菌体密度的影响
2.2 生长因子添加种类及添加量对L-酪氨酸产量的影响
由图2可知,7种生长因子对L-酪氨酸产量有显著的影响。其中维生素B12、MgSO4·7H2O和PLP的影响较大,Mg2+是EMP途径中关键酶的辅酶,还参与了甘油酸-3-磷酸和甘油酸-2-磷酸之间磷酸基团的转移过程[22],为原料磷酸烯醇式丙酮酸的生成打下了基础;磷酸吡哆醛促进了谷氨酸向谷氨酰胺的转化[22],而L-酪氨酸是由前体物质HPP(对羟基苯丙酮酸)与谷氨酸的转氨作用生成的,PLP为其生产提供了动力[23];维生素B12存在两种活化形式,不仅参与了核苷酸与脱氧核苷酸的相互转化,还参与了甲基的转移[22];当谷氨酰胺的添加量≤2 g/L时,L-酪氨酸的产量与谷氨酰胺的添加量呈正相关,当谷氨酰胺添加量为2 g/L时,L-酪氨酸产量达到11.20 g/L,当谷氨酰胺添加量>2 g/L时,L-酪氨酸产量呈下降趋势,维生素H的效果类似;随着KH2PO4添加量的增加,L-酪氨酸产量一直增大,并未像菌体密度呈先增后减的趋势,分析出KH2PO4对产量只呈促进作用,甲硫氨酸的效果类似,分析原因为甲硫氨酸能减少包涵体的生成,提升质粒的表达效果[18]。

图2 单一生长因子对 L-酪氨酸产量的影响
2.3 正交分析探究关键元素的最适用量
实验得出KH2PO4、维生素H、MgSO4·7H2O、维生素B12、PLP对 L-酪氨酸发酵过程产生了较大影响,因此继续优化5种物质的用量。
由表3中极差值的大小可以得出影响L-酪氨酸生物量的5个因素依次为A>D=E>C>B;通过比较5个因素的4个均值,即最大均值对应最适添加量,得到KH2PO4的最适添加量为2.5 g/L;维生素H的最适添加量为1.0 mg/L;MgSO4·7H2O的最适添加量为2.0 g/L;PLP的最适添加量为1.0 mg/L;维生素B12的最适添加量为2.0 mg/L。
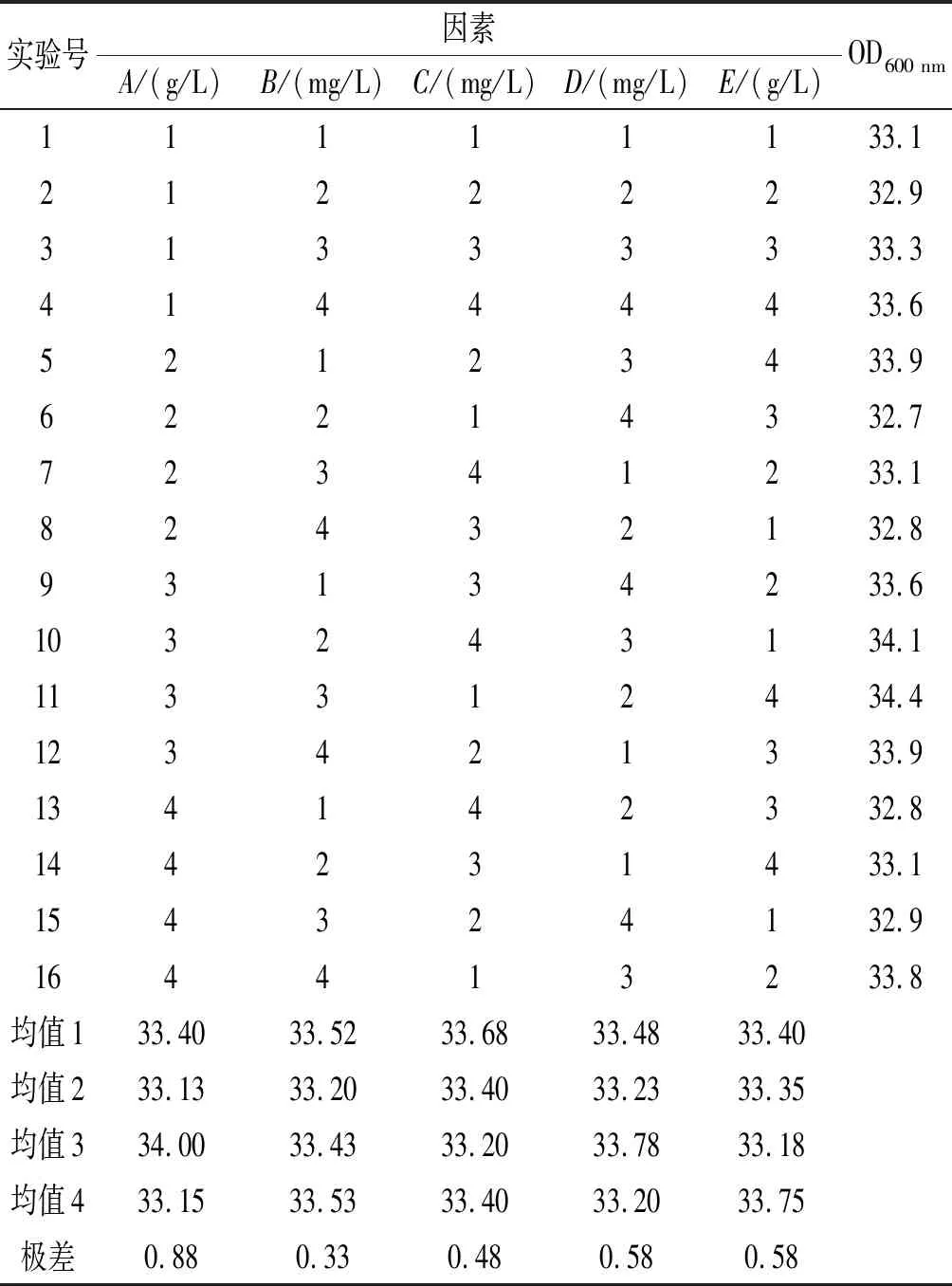
表3 L-酪氨酸生物量正交实验结果
由表4中极差值的大小可以得出影响L-酪氨酸产量的5个因素依次为B>C>A>D=E;通过比较5个因素的4个均值,即最大均值对应最适添加量,得到KH2PO4的最适添加量为2.4 g/L;维生素H的最适添加量为0.8 mg/L;MgSO4·7H2O的最适添加量为1.5 g/L;PLP的最适添加量为1.8 mg/L;维生素B12的最适添加量为2.0 mg/L。
综合来看,维生素H和MgSO4·7H2O的添加量对产量的影响较大,其他3个因素对生物量的影响较大,因此确定维生素H的最适添加量为0.8 mg/L;MgSO4·7H2O的最适添加量为2.0 g/L;KH2PO4的最适添加量为2.5 g/L;PLP的最适添加量为1.0 mg/L;维生素B12的最适添加量为2.0 mg/L。
2.4 5 L发酵罐验证
前期实验以L-酪氨酸的菌体密度和产量为指标,探究了7个生长因子对L-酪氨酸发酵的影响,最后确定了效果较好的5个生长因子的添加量,并通过正交实验进一步对添加量进行了筛选优化。但摇瓶实验具有一定的局限性,导致菌体的生长潜力大大受限,因此在其他培养基成分及培养条件一致的条件下,进行5 L发酵罐对比验证实验,以不添加生长因子的培养基作为对照组,以KH2PO4、维生素H、MgSO4·7H2O、维生素B12、PLP分别添加2.5 g/L、0.8 mg/L、2.0 g/L、2.0 mg/L、1.0 mg/L为实验组,进一步探究生长因子及添加量的优化效果。
由图3可知,优化后实验组的菌体密度和L-酪氨酸产量在发酵期间始终高于对照组,实验组最大菌体密度为63.2 g/L,高于对照组的菌体密度56.7 g/L,发酵过程中菌体密度一直处于上升趋势,但发酵后期菌体生长达到稳定期,所以增长较慢,趋于稳定。当不添加生长因子时,L-酪氨酸产量明显偏低,增长趋势较平稳,且最终产量为49.8 g/L。发酵中期(20~36 h)是L-酪氨酸积累速率最大的时期,且与菌体密度增长趋势相近,当发酵时间达到36 h之后,L-酪氨酸积累较缓慢,最后44 h下罐时L-酪氨酸产量为53.1 g/L,较对照组提高了6.6%。
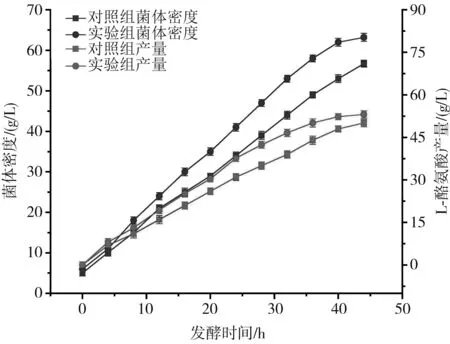
图3 菌体密度与产量的对比
2.5 不同生长因子添加方式对L-酪氨酸发酵的影响
在发酵过程前期,菌体密度可通过发酵条件进行控制,进而缩短发酵周期,若生长因子添加时间过晚,会造成营养不足,导致菌体生长缓慢,发酵产酸水平较低,最后导致产量下降。因此,生长因子的添加方式对L-酪氨酸生物量及产量也具有一定影响。适合的添加方式对L-酪氨酸发酵至关重要。
2.5.1 不同添加方式对产量和糖酸转化率的影响
由图4可知,发酵过程中具体时间监测到的糖酸转化率为实时糖酸转化率,因此能够通过数据呈现的转化率的变化曲线得出发酵过程中转化率最高的时期,如果设法延长这一时期,那么L-酪氨酸产量将会大大提高。a添加方式为一次性添加全部生长因子,因此发酵前期营养丰富,菌体能充分利用营养物质加速地进行菌体生长及L-酪氨酸的合成,糖酸转化率也较高,菌体生长前期8 h时糖酸转化率最高,8~20 h时菌体产酸速率逐渐降低,20~30 h产酸速率呈上升趋势,30 h之后产酸速率较低,分析可能是生长因子前期利用充足但中后期供应不够,不足以维持菌体正常的生长。添加方式b为分时间段(10,15,20,25 h)添加生长因子,可明显看出前期0~10 h未添加生长因子时期L-酪氨酸产率较低,而添加生长因子的10~30 h内菌体大量生产L-酪氨酸,30 h后产酸率大大下降,最终L-酪氨酸产量为50.3 g/L,分析原因为前期菌体生长时期营养物质不足,导致菌体生长受到抑制,进而无法较好地进行L-酪氨酸生产,中期添加生长因子后这一问题有所缓解但最终产量低于a添加方式。添加方式c为发酵8 h后持续流加,能够看出前期菌体生长速率较快,8 h时出现第一次转化率高峰,分析原因是前期营养物质较充足,8~35 h生产较稳定,36 h后糖酸转化率再次达到高峰,糖酸转化率达到一个较平稳的水平,最终产量为56.1 g/L,糖酸转化率达到23.2%,分析可得整个发酵过程中生长因子都处于一个相对稳定的水平,使菌体较好地吸收生长所需的各种营养物质,因此使L-酪氨酸发酵水平达到最高,较其他两种添加方式有明显优势。

图4 不同添加方式对L-酪氨酸产量及实时糖酸转化率的影响
2.5.2 不同添加方式对生物量和耗糖速率的影响
由图5可知,菌体密度和耗糖速率由大到小的顺序都为c>b>a,c组最大菌体密度为65.0 g/L,c组在24 h时耗糖速率最大,为12.3 g/(h·L),而a组和b组最大耗糖速率出现在16 h,分别为10.1,11.2 g/(h·L),较c组早出现了4 h,并且可明显看出c组在36 h之后菌体密度开始缓慢增长,而a组和b组在28 h之后菌体密度便开始出现缓慢增长的趋势,菌体活性明显下降,最终a组和b组的菌体密度分别为53.1,56.0 g/L,远低于c组,因此添加方式c明显优于另外两种添加方式。
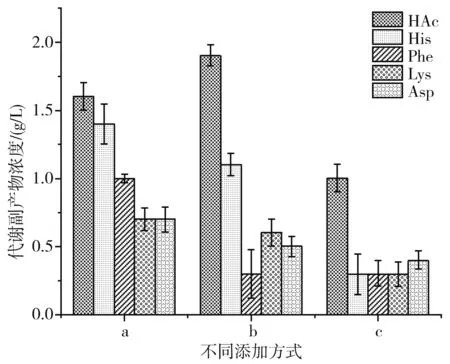
图6 不同添加方式对代谢副产物的影响
2.5.3 不同添加方式对副产物的影响
重组大肠杆菌高密度发酵过程中,很容易出现代谢副产物,其中乙酸最常见,它对蛋白的表达影响较大,研究表明,当菌体的生长速率达到一定值时,菌体密度达到最大值后摄氧能力不再继续增强,进而导致乙酸的生成[24],方式b乙酸最终含量为1.9 g/L,分别比方式a和方式c增加了18.7%和60%,分析方式b会造成菌体过快衰老,菌体摄氧不足导致副产物乙酸含量较高,还会造成原料的浪费。限制比生长速率是减少乙酸生成的方法之一。综合来看,方式c中副产物组氨酸(His)等均少于另外两种添加方式,因此方式c有较大优势。
3 结论
培养基的营养成分和发酵条件控制是大肠杆菌高密度发酵的两个关键因素,实验中选用5个生长因子并对其添加量及添加方式进行了探索,实验表明5个生长因子的最优添加量为维生素H 0.8 mg/L,MgSO4·7H2O 2.0 g/L,KH2PO42.5 g/L,PLP 1.0 mg/L,维生素 B122.0 mg/L,最优添加策略为10 h开始持续流加。最终L-酪氨酸产量为56.2 g/L,最高菌体密度为65.0 g/L,糖酸转化率为23.1%,副产物乙酸积累量减少到1.0 g/L,证明了生长因子的添加使菌体活性大大增强,强化了L-酪氨酸生产的通路代谢,L-酪氨酸产量明显提高,使大肠杆菌生产L-酪氨酸的高密度培养达到一个较高的水平,也为L-酪氨酸发酵生产中各种生长因子的调控提供了依据[25]。
猜你喜欢
当代水产(2022年1期)2022-04-26 14:35:38
科学大众(2022年12期)2022-02-18 11:17:38
河北果树(2021年4期)2021-12-02 01:14:38
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:02
天然产物研究与开发(2018年1期)2018-02-02 07:21:22
中成药(2018年1期)2018-02-02 07:19:57
中国调味品(2017年2期)2017-03-20 16:18:21
现代检验医学杂志(2016年3期)2016-11-15 01:59:48
中国塑料(2016年7期)2016-04-16 05:25:52
山西果树(2014年6期)2015-03-12 20:40:56
