
竹叶花椒-飞龙掌血嫁接树表型及生理性状的抗旱响应
2023-10-10 08:52:06郝佳波郭永清饶绍松辛培尧
干旱地区农业研究 2023年5期
王 飞,郝佳波,胡 青,郭永清,饶绍松,陆 斌,辛培尧
(1. 西南林业大学,国家林业和草原局西南风景园林工程技术研究中心,云南 昆明 650224;2.云南省林业和草原科学院,云南 昆明 650201;3.鲁甸县林业和草原局,云南 昭通 657199)
竹叶花椒(Zanthoxylumarmatum)又称青花椒,隶属芸香科(Rutaceae)花椒属(Zanthoxylum),是一种兼具食用和药用价值的木本油料植物,广泛分布于云南、四川、重庆等地[1]。早在《诗经》中就有文字记载其可用作调味料及中药材[2]。多年来,国内外研究人员致力于花椒栽培管理[3-4]、遗传育种[5-6]、主成分分析与利用[7-8]以及产品贮藏加工[9-10]等多方面的研究,且取得了显著成效。竹叶花椒是云南省传统主栽花椒种之一,主要分布在金沙江、牛栏江流域的干热河谷区以及丽江、楚雄等部分地区[11]。随着花椒产业的发展和良种推广,花椒现已成为云南主要经济树种之一[6]。但由于干热河谷地区特殊的环境和立地条件,干旱少雨、土壤贫瘠等问题日益突出,这严重影响了花椒的推广种植。因此,针对云南竹叶花椒种植区的生态条件,开展提高竹叶花椒相关抗旱性研究,对扩大花椒产区、促进花椒产业的发展至关重要。
嫁接苗是经济林木栽培最常用的苗木类型,而优良砧木的选育对后期嫁接苗木的长势、抗逆性、果实的产量和品质等至关重要[12]。应用砧木良种嫁接可增强栽培品种的抗逆性,如抗旱[13-14]、抗寒[15]、抗盐碱[16-17]以及抗病虫害[18]等。目前,国内外在苹果(Maluspumila)[19]、柑橘(Citrusreticulata)[20-21]、葡萄(Vitisvinifera)[22-23]等果树中有关砧木选育嫁接的研究报道较多,且大部分是通过研究植株各组织表型、解剖结构以及各生理指标的变化来反映砧木嫁接对接穗的影响。但有关花椒砧木选育及利用砧木嫁接来提高花椒抗逆性等方面的研究鲜见报道。龚霞等[24]选用大红袍作砧木,通过嫁接不同接穗品种,为四川南部花椒产区筛选出了少刺且具有一定抗病虫害特性的砧穗组合。蔡麟阁等[25]通过观测3种砧木材料嫁接无刺花椒后接口愈合情况、成活率、生长结果状况及抗病性等指标,发现豆椒作为无刺花椒砧木效果最好。在花椒抗旱砧木选育研究中,龚霞等[26]设置了不同干旱胁迫梯度,测定了6种花椒砧木的膜透性、叶片含水量、生长量等,利用隶属函数法对其抗旱性强弱进行了排序;而有关花椒的解剖结构、内在生理生化抗旱响应机制等却鲜见报道。
飞龙掌血(Toddaliaasiatica)是芸香科飞龙掌血属(Toddalia)多年生木质藤本植物,自然分布于云南、贵州等干热河谷区的路旁、山坡疏林中,其主根深扎,侧根及毛细根密集,对干旱、贫脊的土壤适应性较强[27]。另外,在系统发育研究中,Li等[28]基于16个物种的叶绿体基因组构建最大似然系统发育树,发现飞龙掌血属与花椒属的亲缘关系最为密切。根据前期嫁接和初步栽培观测,发现以飞龙掌血作砧木嫁接竹叶花椒后其嫁接亲和性较好,无“大小脚”现象,且嫁接苗在干旱条件下生长表现良好,是选育抗旱花椒砧木的优良材料。目前有关飞龙掌血嫁接竹叶花椒后的主要性状表现及抗旱响应机制尚未清楚。因此,本研究在观测竹叶花椒和飞龙掌血1年生实生苗根系特征的基础上,对比分析了飞龙掌血做砧木嫁接竹叶花椒后叶片的解剖结构、色素含量及主要生理指标的差异,并测算了果实产量,以期揭示飞龙掌血作为竹叶花椒抗旱砧木的特性和栽培价值,为竹叶花椒抗旱砧木的选育及该嫁接组合的应用推广提供理论基础。
1 材料与方法
1.1 试验材料
材料采集在鲁甸明德花椒庄园进行。明德花椒庄园位于云南省昭通市鲁甸县小寨镇,属低纬度山地季风气候;年平均气温14.7℃,年均无霜期289 d,年均降水量仅74.1 mm。从庄园育苗基地随机各选5株竹叶花椒和飞龙掌血1年生实生苗作为根系观测材料。同时,以2年生飞龙掌血实生树为砧木,接穗在4年生竹叶花椒上采集,用枝接法嫁接。待嫁接树接口愈合后,在园内选取长势基本一致的2年生飞龙掌血、4年生竹叶花椒及其嫁接树各5株作为试验样木,在每株样木树冠外围向阳面采集1年生枝条4~6叶位上完好的健康成熟功能叶,装入采样管,标记后置于液氮中带回实验室备用。所有材料于2022年4—5月采集,保证采样前2个月无降雨及人工灌溉,且此时正值云南旱季末期,植株受干旱的影响较大,嫩叶略卷曲,采集的样品更具代表性。
1.2 实生苗根系观测
清水浸泡并冲洗干净的完整苗木根系,利用万深LA-S植物根系分析仪(杭州万深检测科技有限公司)测量并统计竹叶花椒和飞龙掌血1年生实生苗的根系总根长、体积、平均直径、根尖数以及分叉数等指标。
1.3 叶片解剖结构观察
参考李亚男等[29]提出的石蜡切片法,分别制作飞龙掌血、竹叶花椒及其嫁接树叶子最宽处横切面切片(厚度10~12μm),利用徕卡Leica显微镜观察并拍照。根据图像中标注的比例尺大小,用Image J软件测量叶片组织厚度,取值为多视野下的平均值,并计算栅海比、叶肉组织紧密度及疏松度。
1.4 叶片光合色素含量测定
参考李萍[30]采用的光合色素含量测定方法,利用相关公式分别对应计算飞龙掌血、竹叶花椒及其嫁接树叶片中叶绿素a、叶绿素b、类胡萝卜素的含量。
1.5 叶片生理指标测定
分别测定飞龙掌血、竹叶花椒及其嫁接树叶片中超氧化物歧化酶(SOD)、过氧化物酶(POD)活性和丙二醛(MDA)含量3种主要生理指标。超氧化物歧化酶、过氧化物酶活性和丙二醛含量的测定方法分别为比色法、愈创木酚法和硫代巴比妥酸法[31]。
1.6 果实产量测算
选取冠幅基本一致的竹叶花椒和嫁接树各5株,在果实成熟期测量各单株枝条数、枝条长度、枝条果穗数、鲜果产量及单位面积产量,并计算各指标平均值。
1.7 数据统计分析
用Excel 2003、SPSS 20.0等软件进行数据整理和统计学分析。
2 结果与分析
2.1 竹叶花椒和飞龙掌血的根系特征
结合根系扫描结果(图1)和扫描数据方差分析(表1)可以看出,竹叶花椒和飞龙掌血1年生苗木根系之间的各项扫描参数均呈现显著差异,其中飞龙掌血的总根长、根体积和平均直径分别是竹叶花椒的1.3、1.8、1.2倍。从根系各指标的描述统计可以看出飞龙掌血苗木根系更为密集、发达,说明其对干旱的适应能力更强。

表1 飞龙掌血和竹叶花椒苗木根系特征比较Table 1 Comparison of root characteristics of seedlings in T. asiatica and Z. armatum
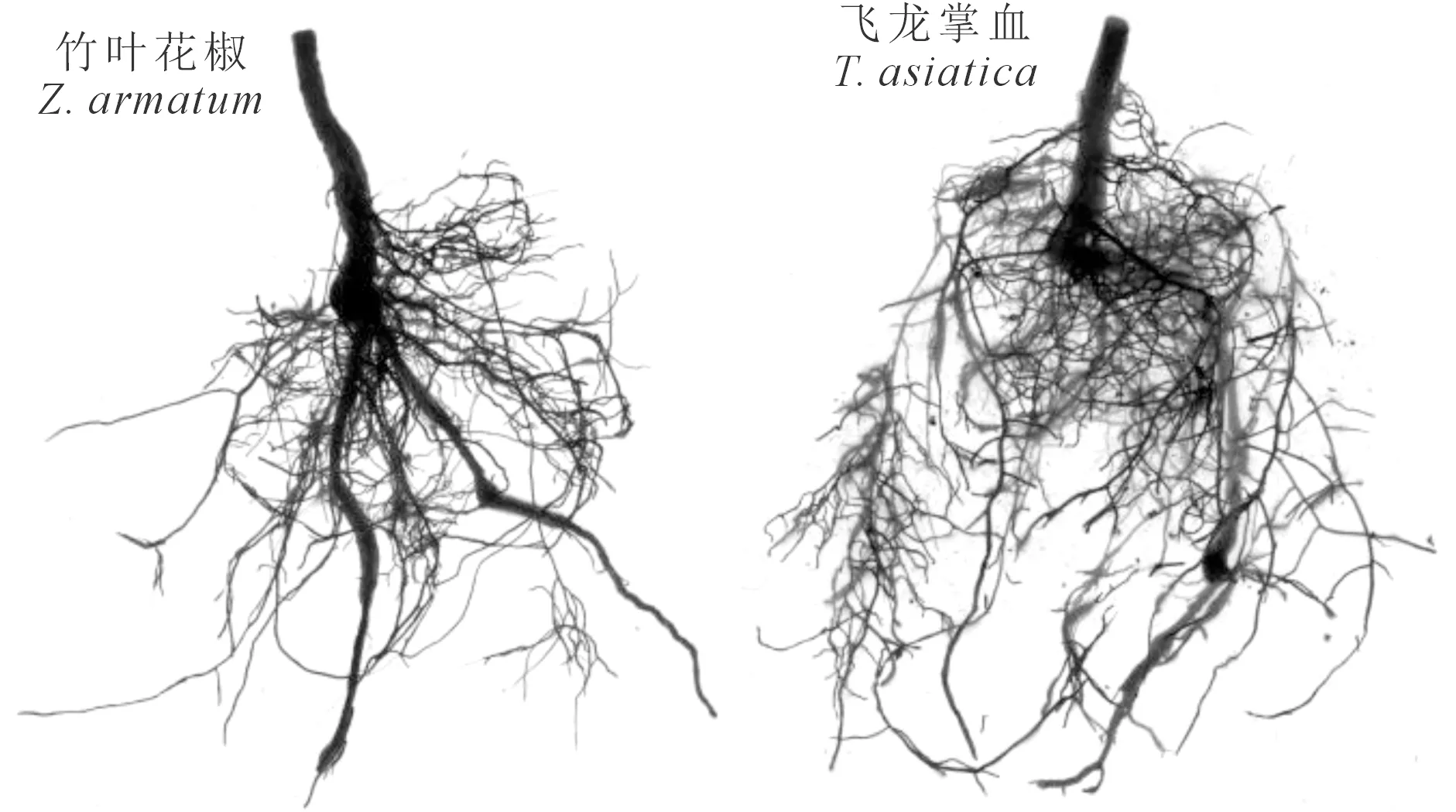
图1 竹叶花椒和飞龙掌血苗木根系扫描Fig.1 Root scanning of seedlings in Z. armatum and T. asiatica
2.2 砧木嫁接对竹叶花椒叶片解剖结构的影响
飞龙掌血、竹叶花椒及其嫁接树的叶片解剖结构和各微观形态指标统计分析结果如图2、表2所示。由图2可以直观地看出,飞龙掌血叶片的栅栏组织非常发达,厚度占到叶片总厚度的50%以上;经计算,发现嫁接树叶片的栅栏组织、海绵组织厚度与竹叶花椒相比分别显著提高了46.57%和35.38%。但相比飞龙掌血,嫁接树叶片的栅栏组织显著降低了45.81%,而海绵组织却显著增加了50.61%。从叶片栅海比(PSR)来看,嫁接树的栅海比与竹叶花椒相比虽提高了9.24%,但两者差异不显著。竹叶花椒和嫁接树叶片的叶肉组织紧密度和疏密度在P<0.05水平上差异显著。相比竹叶花椒,嫁接树叶片的叶肉组织紧密度增加了21.82%(表2)。
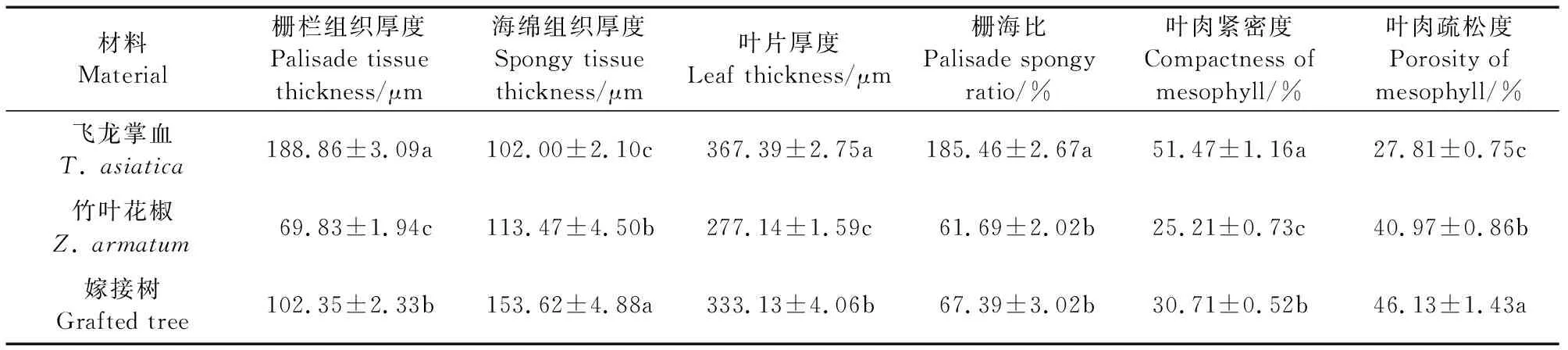
表2 飞龙掌血、竹叶花椒及其嫁接树叶片解剖结构特征比较Table 2 Comparison of anatomical features of T. asiatica, Z. armatum and their grafted tree

注:STT:海绵组织厚度;PTT:栅栏组织厚度。Note: STT: Spongy tissue thickness; PTT: Palisade tissue thickness.图2 飞龙掌血、竹叶花椒及其嫁接树叶片解剖结构横切面Fig.2 Cross-section of leaf anatomy of T. asiatica, Z. armatum and their grafted tree
2.3 砧木嫁接对竹叶花椒叶片光合色素含量的影响
嫁接树叶片的3种光合色素含量均介于飞龙掌血和竹叶花椒之间。嫁接树叶片叶绿素a、叶绿素b和类胡萝卜素含量相比竹叶花椒对应显著增加了13.5%、87.3%、33.5 %;但与飞龙掌血相比,嫁接树对应的3种光合色素含量则显著降低了13.4%、9.9%、29.2%(图3)。嫁接组合的3个物种之间叶片各光合色素含量均达显著差异水平,说明砧木嫁接显著提高了竹叶花椒的光合色素含量,促进了光合作用。

注:图中不同小写字母表示不同材料间差异显著(P<0.05)。Note: Different lowercase letters indicate significant differences among materials (P <0.05).图3 飞龙掌血、竹叶花椒及其嫁接树叶片光合色素含量比较Fig.3 Comparison of photosynthetic pigment content in leaves of T. asiatica, Z. armatum, and their grafted tree
2.4 砧木嫁接对竹叶花椒抗旱生理指标的影响
从飞龙掌血、竹叶花椒及其嫁接树叶片的SOD、POD活性和MDA含量的测定分析结果可以看出(表3),与竹叶花椒相比,嫁接树叶片的SOD和POD活性分别提高了22.1%和6.4%,但MDA含量减少了4.6%;与飞龙掌血相比,嫁接树叶片的SOD和POD活性分别减少了6.2%和15.7%,而MDA含量却提高了84.1%。这3项生理指标在嫁接组合的3个物种间均达显著差异水平。
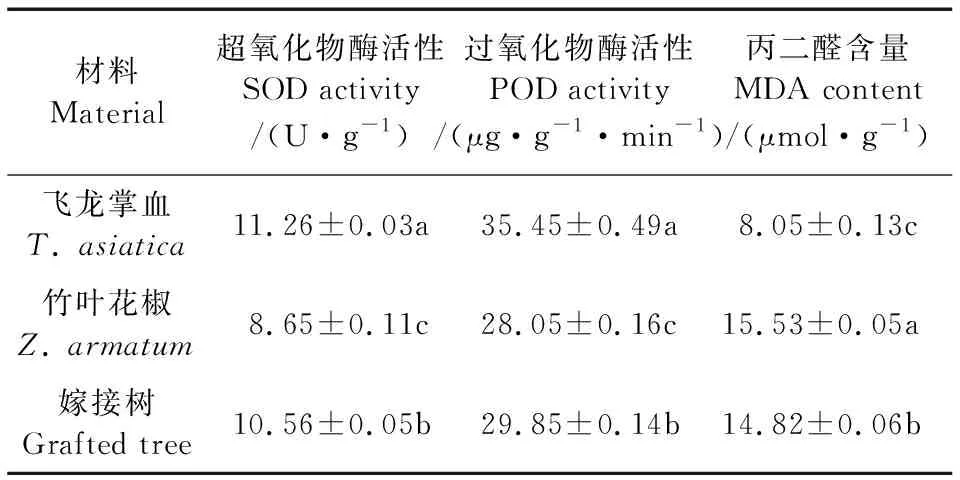
表3 飞龙掌血、竹叶花椒及其嫁接树主要生理指标分析Table 3 Analysis of main physiological indexes of T. asiatica,Z. armatum, and their grafted tree
2.5 叶片解剖结构、光合色素含量和抗旱生理指标的相关关系
表4中3种光合色素含量之间呈极显著正相关,Chl.a与Chl.b、Caro.的相关系数r分别为0.89和0.97;栅海比与栅栏组织厚度(PTT)、3种光合色素含量、SOD和POD活性均呈极显著正相关关系,但与海绵组织厚度(STT)和MDA含量呈极显著负相关关系,而丙二醛(MDA)含量除了与海绵组织厚度极显著正相关之外,与其余各指标(PTT、PSR、Chl.a、Chl.b、Caro.、SOD和POD)均显著或极显著负相关,其中与栅海比的相关系数最大(r=-0.98)。而值得注意的是,海绵组织厚度与3种光合色素含量之间均不相关。

表4 叶片解剖结构、光合色素含量及抗旱生理指标间的相关性分析Table 4 Analysis of leaf anatomy, photosynthetic pigment content and correlation between main physiological indicators
2.6 竹叶花椒和嫁接树产量比较分析
竹叶花椒和嫁接树的树体特征及果实产量统计结果见表5。方差分析显示,竹叶花椒和嫁接树的平均枝条长度、枝条果穗及单位面积产量呈显著差异。其中,相比竹叶花椒,嫁接树的平均枝条果穗和单位投影面积产量分别显著提高了57.1%和34.1%,而平均枝条长度显著降低了31.6%。

表5 竹叶花椒和嫁接树果实产量分析Table 5 Analysis of fruit yield of Z. armatum and its grafted tree
3 讨 论
抗逆性是植物在长期抵抗和适应极端环境的过程中,经自然选择形成的稳定的、可遗传的特性,这种特性可在植物体内部各组织结构及生理生化反应等方面得以体现[32]。竹叶花椒受干热河谷地区特殊气候的影响,干旱成为限制其生长、分布的主要因素[33]。因此,通过优良砧木的选择来提高竹叶花椒抗旱性在生产上有着重要的实践意义。相关研究表明,通过优良砧木嫁接可在一定程度上改变植物内部原有的组织结构及生理生化过程,增强树体抗旱、抗寒等能力,以达到经济林果提质增产的目的[34]。
植物根系是评判植物抗旱性强弱的重要指标,扎根越深、根系越发达就越能适应干旱和贫瘠的环境[35]。通过对竹叶花椒和飞龙掌血1年生实生苗根系扫描数据的分析,发现两者根系的各项扫描指标间均呈显著差异,其中飞龙掌血的总根长、体积和平均直径分别是竹叶花椒的1.3、1.8倍和1.2倍;从数据描述统计结果可以看出飞龙掌血苗木根系更为密集、发达,说明其对干旱的适应能力更强。
叶子是绝大多数植物进行光合作用的器官,对外界环境变化敏感,其叶形和内部解剖结构均能反映植物对周边环境的适应性[36]。如植物比叶面积小,具角质层或蜡质层,栅栏组织发达,上表皮细胞排列紧密等特征都是对干旱生境的响应[37-38]。李鸿雁等[39]对6种牧草叶片的9项与抗旱相关的解剖结构参数进行了测定,并利用叶片各项抗旱相关参数对抗旱能力进行了综合评价,发现不同牧草的叶片厚度、栅栏组织和海绵组织的厚度之间差异显著,且指出栅栏组织越紧密其抗旱能力越强。而本研究通过对飞龙掌血、竹叶花椒及其嫁接树叶片解剖结构分析发现,利用抗旱砧木飞龙掌血嫁接,可显著增加竹叶花椒栅栏组织、海绵组织及叶片总厚度,提高栅海比和叶肉紧密度,这与上述研究结论相符,说明砧木嫁接在一定程度上增强了竹叶花椒的干旱适应性。
植物光合色素含量与其光合生产力紧密正相关,尤其是叶绿素。长期的干旱环境会阻碍植物光合色素的积累,使光合生产力下降[40]。而通过砧木嫁接显著增加了竹叶花椒叶片中3种光合色素的含量,提高了其在干旱环境下的光合生产力,这为植株有机物的积累和正常生长奠定了基础。植物体内部的组织结构与其生理生化过程密切相关[41],如叶片栅栏组织的厚度、紧密度与光合作用相关;气孔的密度、大小与呼吸作用相关。本研究通过对叶片解剖结构、光合色素含量及主要生理指标的相关性分析,发现3种光合色素含量与叶片栅栏组织厚度显著相关,但与海绵组织厚度却不相关,这是因为叶绿体主要存在于叶片栅栏组织中,而光合作用的主要场所就是叶绿体。孙旺旺等[42]在研究连翘叶片解剖结构和色素含量时指出,叶绿体中含有叶绿素、类胡萝卜素等多种光合色素,而叶绿体则主要位于叶片栅栏组织的细胞内。
植物为了适应干旱环境,其抗氧化能力、渗透调节作用以及膜脂过氧化程度等均会随之改变。相关研究表明,在干旱胁迫过程中,各蛋白酶(POD、SOD、CAT等)活性、渗透调节物的含量如脯氨酸(Pro)以及膜脂过氧化反应产物(MDA)含量等均会变化以响应干旱胁迫[43],这对判断植物抗旱性强弱有着重要的指示作用。其中,抗氧化酶POD、SOD等主要参与清除细胞内产生的活性氧自由基(ROS),进而帮助植物体抵御和忍耐干旱、高温等非生物逆境造成的伤害[44]。石晓英等[19]研究了干旱胁迫下3种中间砧对苹果苗生理特征的影响,发现从胁迫开始到结束,POD和SOD活性呈先升后降的趋势,且各砧穗组合的变化幅度存在差异,指出干旱胁迫初期植物体会通过提高抗氧化酶活性来适应干旱环境,这与本研究结果基本一致。通过砧木嫁接,发现嫁接树的POD、SOD活性相比竹叶花椒有了显著的提高,这说明在干旱条件下嫁接树能更多地同时利用POD、SOD清除细胞内的ROS,避免细胞膜损伤以应对干旱环境。另外,干旱胁迫下会加快膜脂过氧化反应,致使MDA过多积累。有研究表明MDA含量和植物抗旱性呈负相关[45]。本研究经测定发现,干旱环境下竹叶花椒的MDA含量最高,而抗旱性较强的飞龙掌血则最低,嫁接树介于飞龙掌血和竹叶花椒之间,在一定程度上表明花椒抗旱性与MDA含量呈反比关系,这与前人研究结果相一致。经相关性分析,发现除了海绵组织厚度之外,MDA含量与其他各指标均呈显著负相关关系,而现有的研究已表明植物抗旱性与海绵组织呈反比关系,与栅海比、光合色素含量及抗氧化物活性呈一定的正相关关系[46],这进一步说明竹叶花椒MDA含量与其抗旱性为负相关。相比竹叶花椒,嫁接树的MDA含量显著降低,这间接反映出嫁接树在干旱环境下细胞膜脂损伤程度较低,说明其干旱适应性更强。
4 结 论
飞龙掌血作砧木嫁接竹叶花椒后,可显著增加竹叶花椒叶片的栅栏组织厚度和叶肉紧密度,促进叶片中光合色素含量的积累,增强光合作用能力,调节主要生理指标的含量,进而提高竹叶花椒的抗旱性。研究结果可为竹叶花椒抗旱砧木选育和该嫁接组合的应用推广奠定理论基础。
猜你喜欢
课堂内外·小学版(低年级)(2024年2期)2024-04-29 00:44:03
Asian Herpetological Research(2023年4期)2024-01-02 06:49:24
西部交通科技(2022年2期)2022-04-27 23:12:50
基层中医药(2021年8期)2021-11-02 06:25:06
今日农业(2020年15期)2020-12-15 10:16:11
少儿美术(2020年9期)2020-11-05 09:11:54
东坡赤壁诗词(2018年1期)2018-03-31 09:10:10
中成药(2018年1期)2018-02-02 07:19:44
天天爱科学(2017年12期)2018-01-31 02:11:36
作文通讯·高中版(2017年11期)2017-12-20 08:09:43
