
不同毛色羊驼皮肤中Agouti和PMEL基因表达与毛纤维中黑色素含量的相关性研究
2023-10-08 00:43:24刘学玮黄昀巍迟志端张俊珍
家畜生态学报 2023年9期
刘学玮,黄昀巍,迟志端,张俊珍*
(1. 山西农业大学 动物科学学院,山西 太谷 030801;2. 山西农业大学 动物医学学院,山西 太谷 030801)
羊驼在传统动物学分类上属于哺乳纲、偶蹄目、骆驼科,是毛用型经济动物,羊驼毛有22种天然色。哺乳动物黑色素(真黑素和褐黑素)是由黑色素细胞产生的,真黑素使毛色表现为黑色或褐色,而褐黑素使毛色表现为黄色或红色。毛色主要受遗传因素控制[1],Agouti基因和PMEL基因都是较为重要的毛色调控基因。
研究表明,小鼠的Agouti基因位于2号染色体,结构复杂,其表达可促进褐黑素的产生并抑制真黑素的产生[2]。真黑素和褐黑素之间的转换通常发生于黑色素细胞,Agouti基因编码的ASIP蛋白(Agouti Signaling Protein)为旁分泌信号因子,随着人、鼠、狐狸、猪、绵羊等的Agouti基因被发现,人们了解到不同物种的Agouti基因及ASIP蛋白具有较高的同源性[3]。
小鼠PMEL基因被定位在第10号染色体上[4],由11个外显子组成并拥有神秘的重复结构域(RPT),该结构域由数量可变的不完全串联重复组成,通过辅助控制淀粉样蛋白的形成影响黑色素的合成和沉积[5]。PMEL基因对哺乳动物黑色素的合成与沉积有重要影响[6],可直接启动黑色小体内纤维的形成,促进黑素小体合成[7]。前黑素小体蛋白(premelanosome protein-17,PMEL-17)直接参与黑色素的合成,在小鼠的PMEL17基因敲除试验中,毛纤维中的黑色素含量减少了40%~50%[8],这表明PMEL对于表皮色素沉积起着重要作用。本研究以羊驼毛纤维为研究对象,分析羊驼皮肤中Agouti与PMEL的表达与毛纤维中黑色素含量的相关性,为毛色的形成机制提供理论依据。
1 材料与方法
1.1 主要材料
白色、棕色和黑色的健康青年Huacaya型羊驼各3只(山西农业大学羊驼养殖基地),用剪毛剪从毛纤维根部取10 mg,并用取皮器取直径4 mm的皮肤组织。
TaKaRa LA TaqTM 聚合酶:DRR20AM 宝生物工程(大连)有限公司;dNTP:D4030A,宝生物工程(大连)有限公司;TaKaRa、AxyPreP DNA凝胶回收试剂盒:康宁生命科学(吴江)有限公司;琼脂糖:Agarose I D0014-50G(Lot: GI-241100),上海生工/SABC(Lot 061180);Tris 碱(三羟甲基氨基甲烷):上海Sangon;Amresco 分装、EDTA (Na2EDTA,乙二铵四乙酸钠盐):华美公司;Sigma 分装、EB(溴化乙锭):美国Fluck;乌贼墨黑色素、Easy Pure Genomic DNA Kit、2×Es Taq Master Mix、DNA Marker、Trizol:Chanmq-mix。
ABI-2720 pcr 仪:美国 Applied Biosystems 公司;天平:上海精密科学仪器有限公司 ;Thermo EC250-90 型电泳仪:美国 Thermo 公司;紫外照相分析系统:天能科技(上海)有限公司;ABI 3730XL 测序仪:美国 Applied Biosystems 公司;Tenbroeck组织研磨机:美国新泽西州米尔维尔惠顿;索尔瓦尔超速离心机:DuPont Instruments,Wilmington,DE,USA。
1.2 试验方法
1.2.1 转录组测序与基因差异表达分析 用Trizol法提取羊驼皮肤总RNA,经反转录后,利用Illumina HiSeq 4000平台对不同毛色羊驼皮肤cDNA文库进行测序。测序长度为150 bp,过滤低质量片段后,获得目标片段。用RESM软件包(版本v1.2.12)计算基因表达RPKM值,基于NOIseq对黑色、褐色、白色羊驼皮肤差异表达基因进行分析(以VicugnapacosVicPac3.2为参考),将差异总数(fold change)≥ 2和可能性(probablity)≥ 0.8的基因定义为差异表达基因。
1.2.2 黑色素含量的测定 以乌贼墨黑色素混悬液为标准,用分光光度法测定毛纤维中真黑素的含量。各30 mg毛纤维样品清洗后,分别加入3 mL 1M NaOH(85 ℃反应4 h)、3 mL 30%的HI(80 ℃反应2 h)、3 mL 0.1 mol/L PBS(pH=10.5,室温剧烈混合10 min)以分别进行碱溶性总黑色素(ASM)、真黑素(EM)和褐黑素(PM)的测定。取3 mg乌贼墨于3 mL双蒸水中,在超声波中溶解5 min,紫外分光光度计读取其吸光值以做对照。用紫外分光光度分别以A350/mg、A400/mg、A400/mg分别读取毛纤维中真黑素、褐黑素和碱溶性总黑素的吸光值,分别与乌贼墨的吸光值做对比后,获取毛纤维中各种黑色素的含量。
1.2.3Agouti和PMEL基因表达分析 取1.2.1中获得的羊驼皮肤总RNA 1 mg,用Chanmq-mix反转录为cDNA。采用SYBR Green检测系统和比较阈值周期(CT)法对羊驼皮肤中Agouti和PMEL的mRNA丰度进行qRT-PCR分析。所有反应在Stratagene Mx3005P实时荧光定量PCR系统上进行。以18S rRNA为内参,采用2-ΔΔCT法计算基因的相对表达量,即Agouti和PMEL mRNA的丰度与18S rRNA的丰度进行归一化。引物序列见表1。

表1 qRT-PCR引物序列Table 1 qRT-PCR primer sequence
1.2.4 统计分析 实时荧光定量PCR和黑色素含量用“平均值±标准差(Mean±SD)“表示,用SPSS v.21. 0 软件(IBM, Armonk, NY, USA)分析数据,采用ANOVA方差 和T检验进行多重比较。不同毛色羊驼毛纤维中黑色素含量与皮肤中Agouti和PMEL的基因表达量之间的相关性用Person皮尔逊相关性分析方法进行分析。
2 结果与分析
2.1 不同毛色羊驼皮肤毛色基因表达谱
以已知的毛色基因为参考,通过对白色和黑色毛色羊驼皮肤的转录组中的差异基因进行筛选,得到23个已知差异表达的毛色相关基因,其中,PMEL、TYR、TYRP、ASIP以及TYRP2(DCT)均为差异较大的毛色调控基因(表2)。

表2 不同毛色羊驼皮肤中毛色相关的差异表达基因Table 2 Differentially expressed hair color related genes in alpaca skins
2.2 不同毛色羊驼毛纤维黑色素含量
通过对不同毛色的羊驼毛纤维中碱溶性总黑色素(ASM)、真黑素(EM)和褐黑素(PM)的含量测定,结果显示随着毛纤维毛色加深,毛纤维中的ASM、EM和PM的含量升高;与白色毛纤维中的黑色素相比,棕色和黑色ASM和EM的含量增加,并呈差异极显著(P<0.01;P<0.001)(图1A,B),而棕色和黑色毛纤维中的PM含量没有显著性差异(图1C)。

图1 不同毛色羊驼毛纤维中总黑色素(A)、真黑素(B)、褐黑素(C)含量注:*P<0.01,**P<0.05,***P<0.001,****P<0.0001,下同。Fig1. Alkali-soluble melanin content (A), eumelanin content (B) and pheomelanin content (C)in wool fibers of alpacas with different hair colorsNote: * means P<0.01,** means P<0.05, *** means P<0.001,**** means P<0.0001,the same below.
2.3 Agouti基因表达及其与黑色素的相关性
由图2可知,Agouti(ASIP)在白色羊驼皮肤中的基因表达高于棕色皮肤和黑色皮肤,尤其是黑色皮肤,呈差异性极显著(P<0.001)。由表3可知,不同毛色羊驼皮肤中的Agouti基因表达量与毛纤维中ASM和EM含量显著相关(P<0.05),而与PM含量没有显著相关性。

图2 不同毛色羊驼皮肤中Agouti mRNA的差异表达Fig. 2 Differential expression of Agouti mRNA in Alpaca skins with different hair colors

表3 不同毛色羊驼皮肤中Agouti的基因表达量与黑色素的相关性Table 3 Correlation of expression of Agouti mRNA in skins with different hair colors and melanin contents in hair of alpacas
2.4 PMEL基因表达及其与黑色素的相关性
由图3可知,随着羊驼毛纤维毛色加深,PMEL在皮肤中的基因表达量明显增加,与白色羊驼皮肤中的基因表达量相比,PMEL在羊驼棕色皮肤和黑色皮肤的基因表达量呈差异性显著 (P<0.05,P<0.001)。

图3 PMEL mRNA在不同毛色羊驼皮肤中的差异表达Fig. 3 Differential expression of PMEL mRNA in Alpaca skins with different hair colors
不同毛色羊驼皮肤中PMEL的基因表达量与黑色素的相关性见表4。由表4可知,不同毛色羊驼皮肤中的PMEL基因表达量与毛纤维中ASM和EM含量显著相关(P<0.05),而与PM含量没有显著相关性。
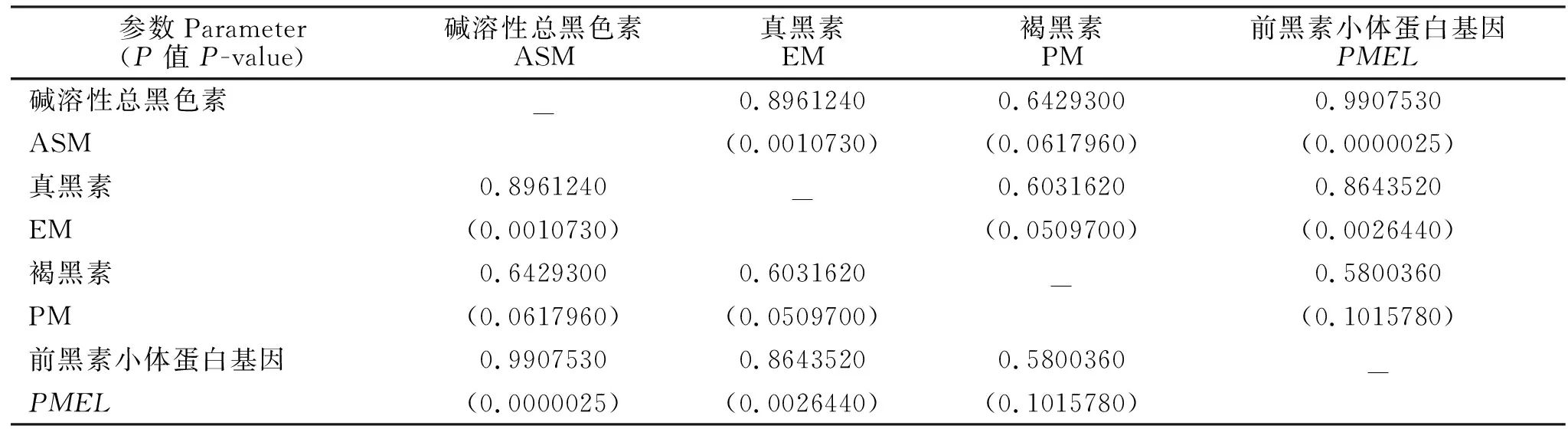
表4 不同毛色羊驼皮肤中PMEL的基因表达量与黑色素的相关性Table 4 Correlation of expression of PMEL mRNA in skins with different hair colors and melanin contents in hair of alpacas
3 讨 论
哺乳动物皮肤中黑色素细胞产生的黑色素分为两类,即真黑素(EM)和褐黑素(PM),黑色素的含量、分布和比例不仅决定哺乳动物的毛发、皮肤和眼睛的颜色[9-10],而且为它们的生存提供了一定的保护,其形成有着复杂的调控机制。从黑色素细胞的分化成熟,到黑色素合成及转运到角化细胞的过程中,有许多基因、miRNAs和lncRNAs等参与[10],并影响哺乳动物的毛色[11]。常作为动物毛色调控或研究相对比较深入的候选基因主要有TYR、TYRP1、MCIR、PMEL、KITLG、Agouti等。
毛色基因Agouti、Pomc和Extension基因位点可调节真黑素(EM)和褐黑素(PM)的比例大小,它们通过黑素皮质素受体1(MC1R)分别编码Agouti信号蛋白(ASIP)、黑色素细胞刺激激素(MSH)和其受体[12]。ASIP使毛囊黑色素细胞合成褐黑素,且与另一种Agouti基因相关蛋白(AgRP)共同调节毛色,同时与脊椎动物皮肤毛囊中黑色素细胞刺激激素(a-MSH)竞争性地结合黑素皮质激素受体1,导致环腺苷酸磷酸水平下降,进一步抑制TYR的活性,最终合成更多黑色素,进而产生浅色被毛[12]。即ASIP基因表达量越低,动物毛纤维颜色越深。本研究结果也证实了Agouti基因表达量与毛纤维毛色的关系,且与毛纤维中ASM和EM含量显著相关,而与PM含量无显著关系,说明Agouti主要调控了毛纤维中的ASM和EM含量。
PMEL基因编码前黑素体蛋白PMEL17,是一种跨膜蛋白,通过蛋白水解在黑素小体(合成和储存黑色素的酸性细胞器)中产生腔内纤维[13]而直接参与黑色素的合成,在小鼠的Pmel17基因敲除突变中显示,毛纤维的黑色素含量减少了40%~50%;在PMEL17成熟过程中,先以前体P1的形式嵌在内质网的膜中,再运送到高尔基体中,通过低聚糖转化到成熟的P2的形式,再投入黑素小体中形成纤维性基质,真黑素的含量上升[7]。同时PMEL基因在a-MSH的调控下表达量增加,进而提高了PMEL17的蛋白表达水平[14]。PMEL基因型和MCIR基因型共同影响着黑色素和褐黑色素的色素沉积。与灰色山羊皮肤中PMEL的表达高于白色山羊皮肤一样,羊驼在黑色毛色的皮肤中的表达也高于白色和棕色毛色的皮肤,表明PMEL在红色或黄色显型中起重要作用[15]。在北极狐皮肤中,Agouti和PMEL的mRNA表达量显著高于心、肝、脾、肾、肌肉组织,且在北极蓝狐和北极白狐中表达量存在差异[17]。也有研究表明,Agouti基因的第一个编码外显子的缺失导致标准银黑狐的毛色表型[18]。从羊驼选择信号的分析来看,ASIP和PMEL分别是羊驼和美洲驼驯化信号基因,亦是正向选择作用的候选基因[19],本试验发现PMEL与ASM和EM的相关性显著,揭示羊驼ASM和EM是深色被毛色形成的主要物质基础。
此外,在毛色形成过程中,角蛋白相关蛋白27-1、KCNG1、GREB1、APOE和CNNM1也是羊驼毛色形成的重要候选基因,可是,其调控哺乳动物毛色形成过程中的功能鲜见报道,有待进一步研究。
4 结 论
在白色和黑色羊驼皮肤中存在23个已知的差异表达基因,Agouti mRNA在白色羊驼皮肤中表达较高,而PMEL mRNA在黑色羊驼皮肤中表达较高;不同毛色羊驼皮肤中Agouti与PMEL的基因表达量均与毛纤维中ASM和EM含量显著相关,揭示了Agouti与PMEL调控羊驼毛色形成的分子机制,为分子育种提供重要的理论依据。
猜你喜欢
学苑创造·A版(2024年5期)2024-06-10 21:55:57
养猪(2021年4期)2021-08-26 10:57:46
环球时报(2020-06-12)2020-06-12 04:14:42
中华皮肤科杂志(2020年8期)2020-01-11 21:33:34
中华皮肤科杂志(2020年4期)2020-01-11 07:00:43
畜牧兽医科学(2018年14期)2018-02-14 01:44:16
作文评点报·低幼版(2018年39期)2018-01-08 07:51:46
中国真菌学杂志(2017年3期)2017-08-31 12:43:04
畜牧兽医学报(2015年3期)2015-07-05 08:22:51
科技知识动漫(2015年6期)2015-06-18 11:39:47
