
橡胶树未授粉胚珠愈伤组织乳管细胞的鉴定及分化研究
2023-09-25 07:56:56谭德冠彭晶付莉莉孙雪飘韩冰莹张家明
热带农业科学 2023年8期
谭德冠 彭晶 付莉莉 孙雪飘 韩冰莹 张家明
(中国热带农业科学院热带生物技术研究所/中国热带农业科学院海南热带农业资源研究院/农业农村部热带作物生物学与遗传资源利用重点实验室,海南海口 571101)
橡胶树(Hevea brasiliensisMuell.Arg.)属大戟科橡胶属,是重要的热带经济林木。其树皮被定期开割,树皮中被割断的乳管排出牛奶状胶乳(细胞质),经收集和加工后成为重要的工业原料——天然橡胶[1]。全球商品天然橡胶的99%来源于橡胶树[2]。
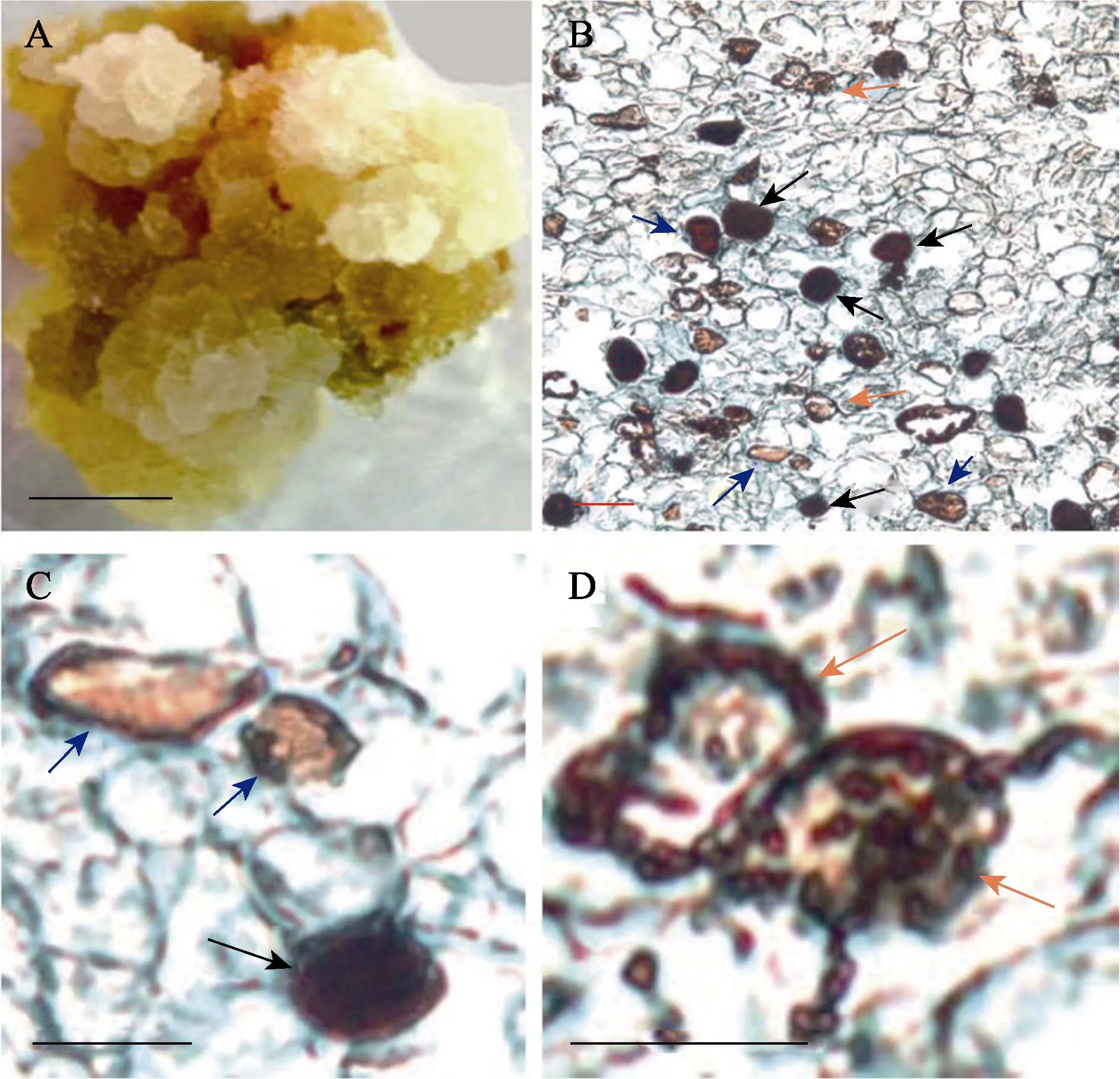
图1 组织化学染色法鉴定未授粉胚珠来源的愈伤组织中乳管细胞
植物乳管是指含有胶乳的高度特化细胞,是橡胶树合成和贮存胶乳的场所[3];橡胶树茎干树皮乳管数量与其产量呈正相关关系[4-5]。因此橡胶树乳管分化机制的研究成为科技人员的重要课题之一。目前大部分科技人员以橡胶树树皮作为模型开展乳管分化机制的研究并取得一定进展。发现外源茉莉酸(JA)能显著促进树皮乳管细胞分化[6];从橡胶树中克隆了HbSKP1、HbJA1、HbCOI1、HbLAP1、HblMYC1、HblMYC2、HbEREBP1等[7-10]与JA信号途径相关的基因。一些学者以橡胶树的花药愈伤组织为模型开展乳管分化研究,发现JA、茉莉酸甲酯(MeJA)也能促进愈伤乳管细胞分化[11-12]。
橡胶树的未授粉胚珠是其组织培养外植体来源之一[13-14]。与花药等其他外植体相比,未授粉胚珠具有容易消毒、能完全避免污染的优势,此外未授粉胚珠与花药的愈伤组织诱导率相近[15]。因此,以未授粉胚珠来代替花药等其他外植体诱导愈伤组织,并以该来源的愈伤组织为模型研究乳管分化机制,可能具有工作量少、效率高等优势。目前尚不清楚橡胶树未授粉胚珠来源的愈伤组织是否存在乳管细胞及其分化情况。本研究通过组织化学法、免疫组织化学法、分子生物学法证实未授粉胚珠愈伤组织中存在乳管细胞,并发现JA能促进未授粉胚珠愈伤乳管细胞分化,其分化依赖于橡胶树基因型,为研究乳管分化机制的模型提供实验依据。
1 材料与方法
1.1 材料
于中国热带农业科学院试验场三队采集橡胶树品种热研7-33-97、热研88-13、热研8-79的花枝,立即置于4℃冰箱中贮存24 h。
1.2 方法
1.2.1 试材制备用镊子收集未开放的雌花,75%(V/V)酒精消毒1 min,无菌水清洗1遍,0.1%升汞消毒10 min,无菌水清洗5次。在无菌条件下用解剖针剖开雌花挑出胚珠,接种于诱导愈伤组织培养基中。培养条件为25℃暗培养。诱导愈伤组织培养基的组分是以MS培养基为基本培养基,添加2 mg/L 2,4-D,0.5 mg/L 6-BA,1 mg/L NAA,7%(m/V)蔗糖,植物凝胶2.2 g/L,调整pH至5.8。
1.2.2 组织化学法鉴定愈伤组织乳管细胞参照田维敏等[16]报道的组织化学法鉴定未授粉胚珠愈伤组织中的乳管细胞。取培养至70 d的热研7-33-97未授粉胚珠愈伤组织于80%酒精中室温固定2 d,酒精系列脱水,60℃下溴-碘溶液染色48 h,冰醋酸清洗2次,再进行渗蜡、包埋,滑走切片机(Leica,德国)切片,贴片,二甲苯脱蜡,固绿染色,中性树胶封片。将制备好的切片于光学显微镜下观察拍照。
1.2.3 免疫组织化学法鉴定愈伤组织乳管细胞参照Tan等[17]报道的免疫组织化学方法鉴定未授粉胚珠愈伤组织中的乳管细胞。取上述固定好的热研7-33-97未授粉胚珠愈伤组织,酒精系列脱水后进行渗蜡、包埋、切片、贴片,二甲苯脱蜡。将样品置于95℃的0.01 mol/L柠檬酸钠缓冲溶液中处理15 min,PBS缓冲液清洗3次,2%牛血清蛋白(BSA)封闭30 min。将样品置于保湿盒中,加入一抗[小橡胶粒子蛋白(Small Rubber Particle Protein,SRPP)兔抗],按1:100稀释于含1% BSA的PBS缓冲液)后4℃处理过夜,PBS缓冲液清洗3次后,加入二抗(羊抗兔,按1:50稀释于含1% BSA的PBS缓冲液),37℃下处理1 h,PBS缓冲液清洗3次;样品用DBT/BCIP碱性磷酸酶显色液浸泡5 min,酒精系列脱水,中性树胶封片,光学显微镜下观察拍照。
1.2.4 橡胶合成相关基因片段的扩增取培养70 d的热研7-33-97未授粉胚珠愈伤组织,采用北京TIANGEN公司的RNAprep Pure Plant Kit试剂盒提取其总RNA,再用Thermo Scientific公司的RevertAid First Strand cDNA Synthesis Kit试剂盒将总RNA反转录为第一链cDNA。根据从NCBI网站上查找到与橡胶合成相关的基因SRPP基因序列(AJ223388)、橡胶延伸因子(Rubber Elongation Factor,REF)基因序列(X56535.1),采用Primer 5.0软件设计引物,其中SRPP基因引物对为SRPP-F(5'-GAAGAGGTGGAGGAAGAG-3')、SRPP-R(5'-CAAAGGCAAATAA GAGGA-3'),REF基因引物对为REF-F(5'-GGCTGAAGACGAAGACAA-3')、REF-R(5'-GCAAAGAAG AAGCCAGAG-3)。PCR反应条件为94℃预变性2 min,94℃变性30 s,56℃退火30 s,72℃延伸30 s,反应进行35个循环,最后72℃补平10 min。对PCR产物进行琼脂糖凝胶电泳检测及测序。
1.2.5 JA对未授粉胚珠愈伤乳管细胞分化的影响在诱导愈伤组织培养基上分别加入0、1、2、3、4 mg/L的JA,将热研7-33-97的未授粉胚珠接种于含不同浓度JA的培养基中,25℃下暗培养。80%酒精固定培养至70 d的各处理愈伤组织,经酒精系列脱水,按1.2.2方法进行组织化学染色和制片。光学显微镜下观察拍照,统计不同浓度JA处理的愈伤乳管细胞发生频率。乳管细胞发生频率=(乳管细胞数量/总的细胞数量)×100%。
1.2.6 不同橡胶树基因型对未授粉胚珠愈伤组织乳管细胞分化的影响分别取在相同培养基及培养条件下培养70 d的热研7-33-97、热研88-13、热研8-79未授粉胚珠愈伤组织固定于80%酒精中,酒精系列脱水,按1.2.2方法进行组织化学染色和制片。光学显微镜下观察不同基因型的未授粉胚珠愈伤组织乳管细胞分化情况。
2 结果与分析
2.1 组织化学法鉴定愈伤组织乳管细胞
组织化学法是一种被广泛应用、经典的鉴定植物组织中乳管细胞的方法[18]。当培养至70 d时,未授粉胚珠分化出的愈伤组织呈浅黄色,表面结构紧密(图1-A)。经溴-碘染色液处理后发现,愈伤组织存在棕色的细胞(图1-B、1-C),这些细胞为乳管细胞,其内含物橡胶被溴-碘染色液处理后变成不溶于二甲苯的、棕色的变性橡胶。愈伤组织中还存在黑褐色的细胞,这些细胞为单宁细胞(图1-C),其单宁物质被染色液染成黑褐色。此外,愈伤组织中还存在淀粉细胞(图1-D),该类细胞中充满颗粒状淀粉。
2.2 免疫组织化学法鉴定愈伤组织乳管细胞
为进一步验证未授粉胚珠来源的愈伤组织是否存在乳管细胞,对该愈伤组织进行了免疫组织化学研究。对愈伤组织的切片进行一抗为SRPP兔抗的处理,该抗体SRPP蛋白能与切片中乳管细胞的SRPP底物特异结合,经羊抗兔的二抗处理,再经DBT/BCIP碱性磷酸酶显色液显色发现,愈伤组织切片中存在蓝黑色的细胞(图2),这些细胞为乳管细胞。免疫组织化学法证实了未授粉胚珠来源的愈伤组织存在乳管细胞。
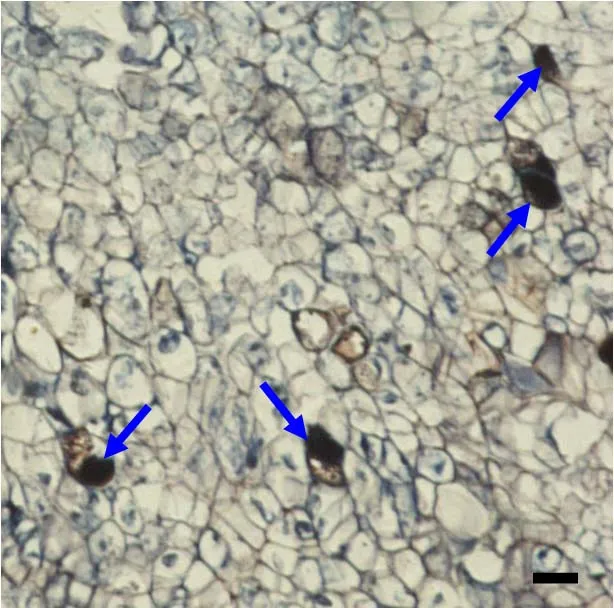
图2 免疫组织化学法鉴定未授粉胚珠来源的愈伤组织中乳管细胞
2.3 橡胶合成相关基因片段的扩增
SRPP、REF是附于小橡胶粒子膜的蛋白质,它们是参与橡胶生物合成的重要酶和调控因子,编码SRPP、REF基因在橡胶树胶乳细胞中特异表达[19-20]。在本研究中,RT-PCR产物电泳结果显示,从橡胶树未授粉胚珠愈伤组织的cDNA中均能扩增出清晰的SRPP、REF基因片段目的条带(图3)。经测序分析,SRPP、REF基因片段的目的条带分别为571、355 bp,经Blast比对发现,它们的序列与已知的橡胶树胶乳表达的基因序列相一致。这从分子水平上进一步证实未授粉胚珠愈伤组织存在乳管细胞,它们的功能与树皮乳管细胞相类似。
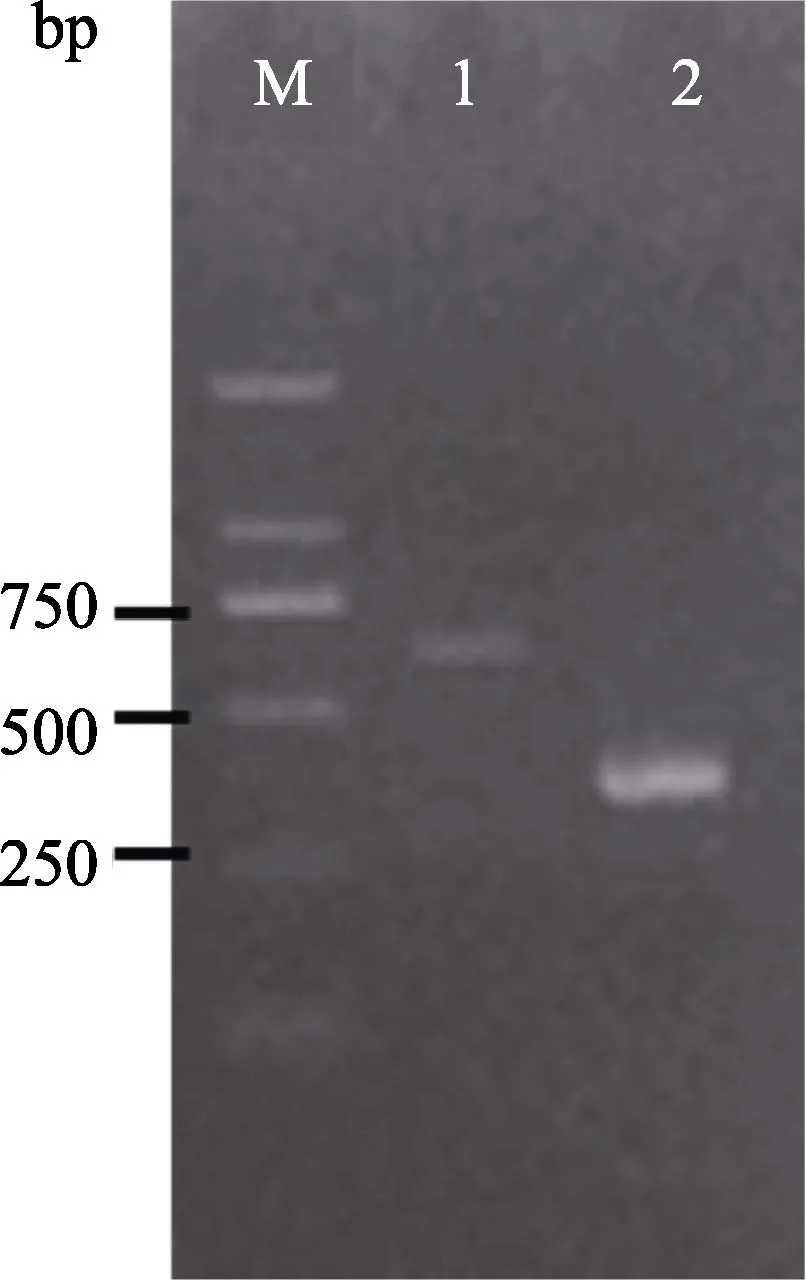
图3 RT-PCR扩增胶乳合成相关基因片段电泳图
2.4 JA对橡胶树未授粉胚珠愈伤乳管细胞分化的影响
施加外源JA已被证实具有促进橡胶树树皮乳管分化的功能[6],但目前尚不清楚是否对未授粉胚珠愈伤组织乳管细胞具有效果。在本研究中,笔者将不同浓度的JA添加于诱导愈伤的组织培养基中,结果发现,JA会显著影响未授粉胚珠愈伤组织乳管细胞分化:当培养基中JA浓度为1 mg/L时,愈伤乳管细胞发生频率增加至8.8%,显著高于其他处理(图4)。当培养基中JA浓度为2、3 mg/L时,愈伤乳管细胞发生频率分别下降至5.8%、5.4%,略低于对照(6.2%),但三者在统计学上差异不显著。当培养基中JA浓度为4 mg/L时,愈伤乳管细胞发生频率下降至2.3%,显著低于其他处理(图4)。在JA浓度1~4 mg/L时,随着JA浓度升高,愈伤乳管细胞发生频率呈逐步下降趋势(图4)。这些结果表明,低浓度的JA能促进未授粉胚珠愈伤组织乳管细胞分化,而高浓度的JA会抑制未授粉胚珠愈伤组织乳管细胞分化,其中以JA浓度为1 mg/L效果最佳。

图4 JA对未授粉胚珠来源的愈伤组织乳管细胞分化的影响
2.5 不同橡胶树基因型对未授粉胚珠愈伤组织乳管细胞分化的影响
在培养条件一致的条件下对3个橡胶树品种的未授粉胚珠的愈伤乳管细胞发生频率进行统计分析发现,不同橡胶树基因型其未授粉胚珠愈伤乳管细胞发生频率存在差异。热研7-33-97与热研8-79的未授粉胚珠愈伤乳管细胞发生频率较接近,分别为6.2%与5.9%,在统计学上没有显著差异,但均显著高于热研88-13(3.2%)(图5)。热研88-13的未授粉胚珠愈伤乳管细胞发生频率最低,比热研7-33-97、热研8-79分别低94%、84%(图5)。上述结果表明,未授粉胚珠愈伤组织乳管细胞分化依赖于橡胶树基因型。
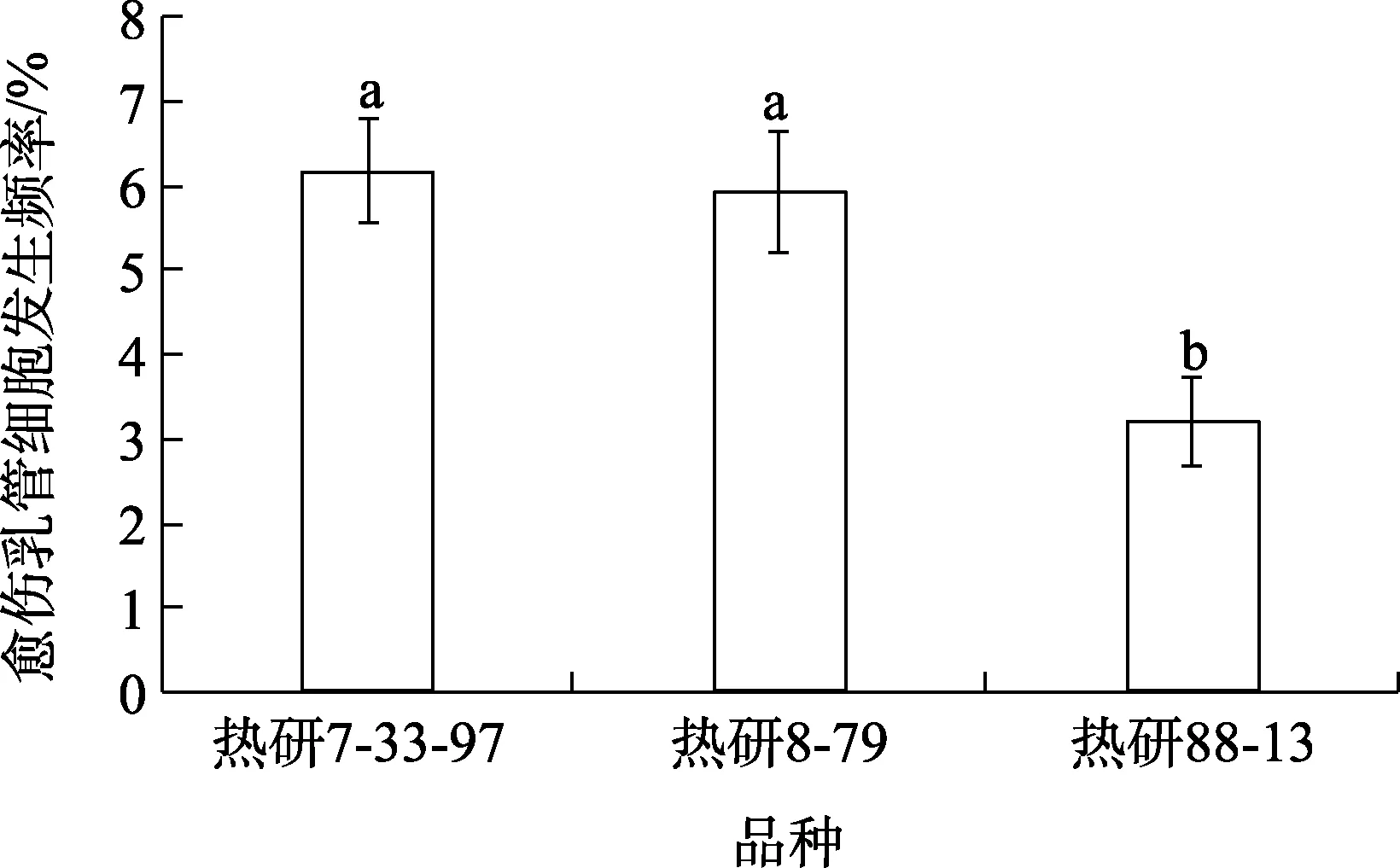
图5 不同橡胶树基因型的未授粉胚珠来源的愈伤组织乳管细胞分化频率的比较
3 讨论与结论
3.1 讨论
本研究通过组织化学法、免疫组织化学法和分子生物学法证实未授粉胚珠愈伤组织中存在乳管细胞;RT-PCR结果显示,SRPP、REF基因均能从未授粉胚珠愈伤组织cDNA中扩增出来,经Blast比对发现,它们的序列与已知的橡胶树胶乳表达的基因序列相一致,表明未授粉胚珠愈伤乳管细胞,能表达与树皮乳管细胞相一致的、与胶乳合成相关的特异基因;同时发现,未授粉胚珠愈伤乳管细胞与树皮乳管细胞一样[6,21],它们的分化受外源JA调控。这些结果为橡胶树未授粉胚珠愈伤组织可作为研究乳管分化机制的模型提供充分的实验依据。
目前已报道的用于研究橡胶树乳管分化的愈伤组织模型来源于橡胶树花药[17]、茎段[5]、叶柄[22]等外植体。橡胶树长期生长在高温潮湿的环境下,该环境富含微生物[23],其茎段和叶柄均长期暴露于外界环境中,环境中的微生物极易滋生在它们的皮层表面,并可能进一步侵染进它们的皮层细胞间/内;橡胶树的花药在发育早期被花被紧密包裹,但在发育至单核期时,花被包裹花药并不紧密,外界环境的微生物会通过雨水等媒介侵染至花药中,使花药组织不再处于无菌环境。因此以橡胶树花药、茎段、叶柄等作为外植体来诱导愈伤组织,存在污染率高的缺点。未授粉胚珠被橡胶树的子房紧密包裹住,属无菌环境,因此以其为外植体来诱导愈伤组织,能完全避免污染现象发生。同时未授粉胚珠接种效率远高于花药[15]。因此,以未授粉胚珠来源的愈伤组织为模型,研究橡胶树乳管细胞分化机制,可能具有工作量少、效率高等优势。
外源JA已被证实具有促进橡胶树树皮乳管细胞分化,从而增加乳管细胞数量的功能,但报道中只具有正调控橡胶树乳管分化的效果[6,21]。本研究发现,低浓度JA具有正调控橡胶树未授粉胚珠愈伤组织乳管分化效果,而高浓度JA具有负调控橡胶树未授粉胚珠愈伤组织乳管分化效果,因此,如果以未授粉胚珠愈伤组织为模型研究乳管分化可能会获取更多的生物学信息。
本研究发现培养70 d的橡胶树未授粉胚珠愈伤组织中存在乳管细胞,但该乳管细胞来源尚不清楚。该乳管细胞可能由胚珠组织中的乳管细胞分化产生,也可能由愈伤组织中薄壁细胞分化产生,因此需要对未授粉胚珠愈伤组织乳管细胞的整个分化过程进行系统研究。
自然界中存在12 500种含胶乳植物,它们分属22个科900个属[24],但目前尚未有其他植物未授粉胚珠愈伤组织中存在乳管细胞的报道。含胶植物乳管细胞的主要功能是起防御作用,除合成和贮存橡胶外,还会分泌一些有毒代谢物来保护植物体免受外来侵害[25]。目前尚不清楚橡胶树未受粉胚珠愈伤组织的乳管细胞是否也存在防御功能,需要对它们的代谢产物成分作深入研究。
3.2 结论
本研究采用组织化学法、免疫组织化学法、分子生物学法证实未授粉胚珠愈伤组织中存在乳管细胞。RT-PCR结果显示,小橡胶粒子蛋白(SRPP)、橡胶延伸因子(REF)基因均能从未授粉胚珠愈伤组织cDNA中扩增出,Blast比对发现,它们的序列与已知的橡胶树树皮胶乳表达的基因序列相一致,表明其功能与树皮乳管细胞相似。培养基中添加低浓度的茉莉酸(JA)能促进未授粉胚珠愈伤组织乳管细胞分化,而高浓度的JA会抑制其分化,JA浓度以为1 mg/L效果最佳。橡胶树未授粉胚珠愈伤组织乳管细胞分化依赖于基因型。本研究的结果为橡胶树未授粉胚珠愈伤组织可作为研究乳管分化机制的模型提供实验依据,对促进乳管分化机制研究具有一定意义。
猜你喜欢
生物技术通报(2023年2期)2023-03-07 12:54:40
热带作物学报(2022年11期)2022-12-16 02:56:10
蔬菜(2022年7期)2022-07-29 15:47:37
山东农业大学学报(自然科学版)(2020年2期)2020-05-21 00:18:06
林业科学(2020年3期)2020-04-28 07:39:18
热带作物学报(2019年1期)2019-06-11 09:40:06
广西植物(2016年10期)2016-11-11 06:51:39
西南农业学报(2016年4期)2016-05-17 05:41:45
江苏农业学报(2015年6期)2015-03-26 10:57:58
中国蔬菜(2013年8期)2013-01-28 04:52:48
