
遮光对西藏林芝市草莓脱毒原种苗光合与叶绿素荧光特性的影响
2023-09-13 12:14敬志豪郭亚轩边巴吉拉桑木顿
黑龙江科学 2023年16期
敬志豪,郭亚轩,伍 海,边巴吉拉,拉 姆,桑木顿
(西藏农牧学院 植物科学学院,西藏 林芝 860000)
光合作用对作物产量具有重要的影响[1]。光照强度是影响光合作用的主要因素之一,每种植物适宜的光照强度不同,光照过强或不够时均会影响光合作用的进行,进而影响有机物积累[2]。长时间的强光照射对植物光合作用有明显的抑制作用[3],通过人工控光,将光照强度调低,不但会影响植物的生长,还会影响其发育。本实验在高原强光照的自然环境下采用遮光处理,研究宁玉草莓脱毒原种苗光合与叶绿素的荧光特性,为草莓苗在高原地区生长的光照调控提供参考。
1 材料与方法
1.1 实验设计
试验于2021年7—8月在西藏农牧学院农场实习地日光温室外空地进行。使用遮阳网、纱网等遮光材料,将光照强度调整为开放环境中的75%、50%、25%和对照,分别记为T1、T2、T3、CK。试验期间,日平均温度为19.06 ℃,供试材料为宁玉草莓的组培脱毒原种苗。
1.2 测定指标与方法
1.2.1 光合特性指标测定
在遮光处理后第3 d选择草莓中上部的功能叶,利用Li-6400XT光合仪(LI-COR公司生产,美国)在晴天9:00-11:00时测量净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)及胞间CO2浓度(Ci)4项指标,重复3次。按公式计算:水分利用效率(WUE)=Pn/Tr。
1.2.2 叶绿素荧光参数测定
分别在遮光处理后第1 d、2 d、3 d选取草莓中上部的功能叶,采用OS5p+叶绿素荧光仪(北京澳作生态仪器有限公司生产,中国)测量叶绿素荧光的各项指标。在Fv/Fm测量模式下,用暗适应叶夹对准选取的叶片进行暗适应约30 min,以充分进行暗适应。测定荧光参数Ft、Fo、Fm、Fv/Fm、Fv/Fo等。在Yield模式下获取各种荧光参数Ft、F′、Fm′、ETR及Yield等。其中Ft代表终端荧光值,Fo为暗适应后的最小荧光参数,Fm为暗适应后的最大荧光参数,Fv/Fm是暗适应后PSⅡ的最大量子产量,Fv/Fo代表PSⅡ的潜在活性,Yield是利用饱和脉冲法进行荧光淬灭分析的根本,此时PSⅡ的有效量子产量最接近光合作用的实际量子产量[7]。Fv/Fm及Yield的计算公式如下:
1.3 数据处理
采用Microsoft Excel 2010进行数据统计并作图,使用IBM SPSS statistics 21软件进行分析。
2 结果与分析
2.1 遮光率对草莓叶片光合参数的影响
由表1可得,Pn随着遮光率的增大而呈下降的变化趋势,Gs、Tr随着遮光率的增大呈先上升后下降的变化趋势,与CK差异显著(P<0.05)。其中,CK与50%遮光处理相比,Pn、Gs、Ci、Tr显著差异,WUE无显著差异。50%遮光处理与25%遮光处理相比,Gs、Tr、WUE无显著差异,Pn、Ci显著差异。与CK相比,Pn、Gs、Tr均下降,Pn依次下降了15.45%、29.74%、40.16%,Gs依次下降了40.91%、35.45%、58.18%,Tr依次下降了41.45%、36.01%、59.84%。Ci则随着遮光率的减小呈现持续下降趋势,在50%遮光处理时达到最大值,与CK相比,Ci相对CK升高了24.22%。WUE随着遮光率的减小呈先上升再下降后上升的变化趋势,均大于CK,在50%胁迫遮光率时达到波峰,与CK相比,WUE相对CK升高了44.22%。

表1 遮光率对草莓叶片光合参数的影响
2.2 遮光率对草莓叶片叶绿素荧光参数的影响
2.2.1 遮光率对草莓叶片初始荧光的影响
Fo的减少表明天线色素的热耗散增加[4],Fo的增加则表明PSⅡ反应中心破坏或可逆失活[5]。由表2可以看出,75%、50%、0%遮光率的初始荧光在1 d、2 d、3 d均呈现先下降后升高的趋势,25%遮光率的初始荧光呈持续升高趋势,75%、50%、25%、0%遮光率在3 d所测量的Fo均高于1 d,但浮动幅度不大,表明遮光率与初始荧光Fo无关。

表2 遮光率对草莓叶片叶绿素荧光暗适应参数的影响
2.2.2 遮光率对草莓叶片实际光量子产量的影响
Yield能够反映PSⅡ反应中心在部分关闭情况下的实际原初光能捕获效率,可作为植物叶片光合电子传递速率快慢的相对指标[6]。由表3可以看出,随着时间的增长,75%遮光率呈现逐渐上升趋势,50%、25%、0%遮光率呈现先上升后下降的趋势。50%遮光率在2 d上升幅度较大,比1 d高5.47%。25%遮光率在3 d下降幅度较大,比2 d低6.45%。这说明一定遮光率可以提高草莓叶片实际光量子产量。
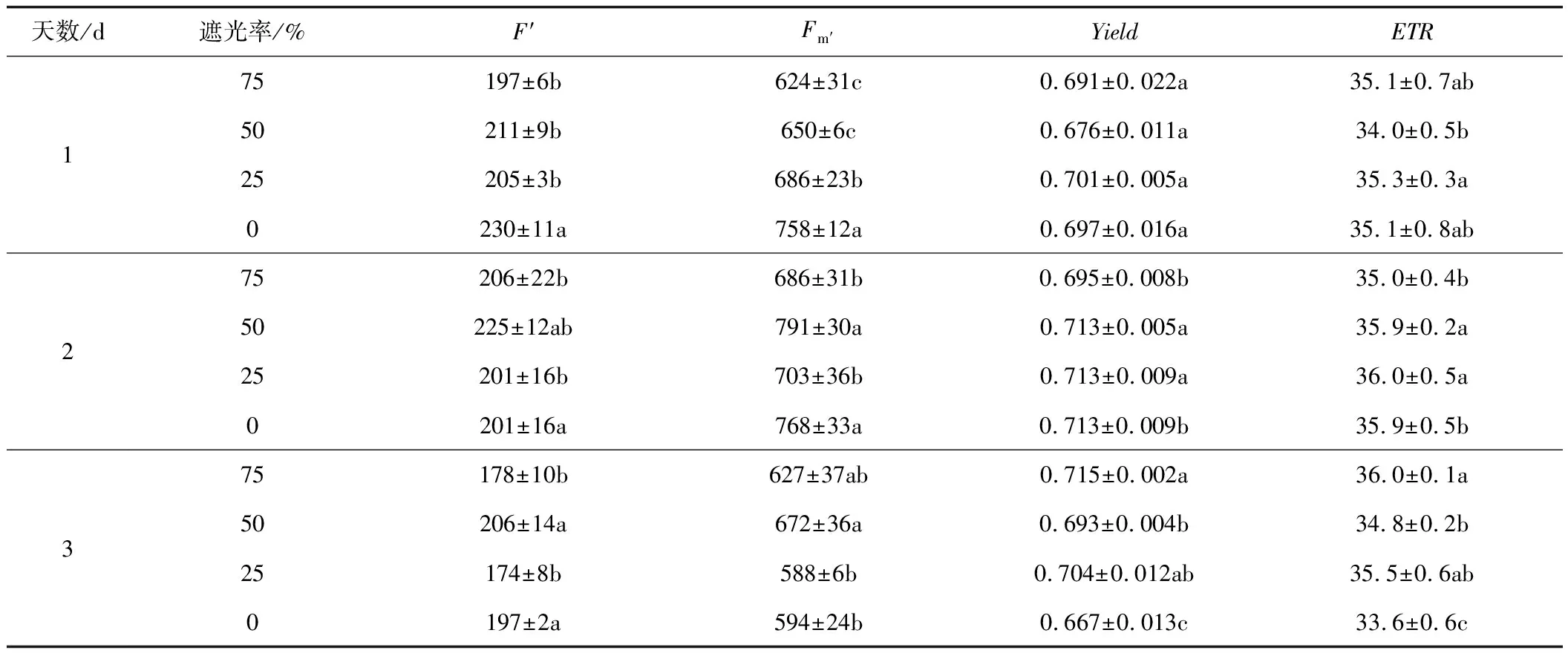
表3 遮光率对草莓叶片叶绿素荧光光适应参数的影响
2.2.3 遮光率对草莓叶片Fv/Fm、Fv/Fo、ETR的影响
PSⅡ反应中心可以将天线色素吸收的过量光能以热耗散的形式消耗,从而避免对光合机构造成失活或破坏[7]。ETR在一定程度上可以反映出PSⅡ反应中心的电子传递速率。由表2、表3可以看出,随着时间的增长,遮光率处理的Fv/Fm、Fv/Fo均呈逐渐下降的变化趋势,ETR呈现先上升后下降的变化趋势。在Fv/Fm中,3 d比1 d分别下降2.05%、2.91%、2.83%。随着遮光率的增大,Fv/Fm、Fv/Fo均呈现先下降再升高后下降的变化趋势,均在在50%遮光率时达到波谷,25%遮光率时达到波峰。随着遮光率的增大,ETR在1 d和3 d时均呈现先下降再升高后下降的变化趋势,在50%遮光率时达到波谷,25%遮光率时达到波峰,但2 d时,ETR呈现先上升后下降的变化趋势,且在25%遮光率时达到波峰。各处理组的ETR在2 d达到最大值,说明遮光率的增大提高了草莓光合作用光系统Ⅱ反应中心的电子传递速率。
3 结论与讨论
叶绿体是光合作用的场所,叶绿素含量的高低直接关系到光合速率的大小。研究发现,净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)均显著高于CK,说明此时植株的光合作用受到了抑制。大量研究表明,影响植株Pn降低的因素包括气孔因素与非气孔因素[8]。本研究中,随着遮光率的增大,50%、0%遮光率处理草莓叶片Pn下降的同时,其Gs、胞间CO2浓度(Ci)相应下降,而在25%遮光处理草莓叶片Pn下降的同时,其Gs下降而Ci升高,说明50%、0%遮光处理与25%遮光处理影响Ci变化的原因不同,50%、0%遮光处理Ci降低的主要原因是由于气孔因素造成的,而非气孔因素是25%遮光处理Ci升高的主要原因,这与张志刚等[9]对辣椒幼苗低温、低温弱光处理的研究结果一致,因为非气孔因素阻碍了CO2的利用[10],造成细胞间隙CO2的积累[11]。
叶绿素荧光参数可直接反映胁迫对光合作用的内在影响,Fo、Fv/Fm的变化可推断反应中心的变化情况及可能的光保护机制。初始荧光Fo升高表明其光合机构遭到破坏,而Fo降低则表明其非光化学能量耗散的增加。经遮光率处理后,75%、50%、0%遮光处理Fo均有下降的过程,后又上升,说明本试验草莓在暗适应过程中的首要原因是非光化学能量的耗散。随着遮光率的降低,均能够提高草莓Yield、Fv/Fm、Fv/Fo、ETR叶绿素荧光参数,但不做遮光处理则表现出一定的抑制。3 d时,草莓Yield、Fv/Fm、Fv/Fo、ETR叶绿素荧光参数均出现了下降变化趋势,造成这一结果的因为是植物为了更好地适应逆境条件,体内的一些蛋白发生变性、渗透调节物质含量变化或一些酶的活性受到了显著的抑制,进而抑制了草莓叶绿素的合成,造成叶绿素荧光参数的下降。50%遮光处理对草莓生理特性影响显著,表明草莓不适合西藏的强光环境,需要适当遮光,最佳遮光率为25%~50%。
猜你喜欢
科学(2022年4期)2022-10-25
干旱地区农业研究(2022年1期)2022-01-28
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
阅读与作文(小学高年级版)(2017年10期)2017-10-11
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
中国铸造装备与技术(2015年5期)2015-12-10
少儿科学周刊·儿童版(2015年10期)2015-11-07
爆笑show(2015年4期)2015-06-24
