
高寒草甸植物群落功能多样性对不同生长期干旱的响应机制
2023-09-11 07:26刘慧慧胡国铮干珠扎布冀国旭王子欣高清竹王庆锁旦久罗布
生态学报 2023年16期
刘慧慧, 胡国铮, 干珠扎布, 冀国旭, 王子欣, 高清竹, 王庆锁,严 俊, 旦久罗布
1 中国农业科学院农业环境与可持续发展研究所, 北京 100081 2 农业农村部农业农村生态环境重点实验室,北京 100081 3 国家农业环境数据中心,北京 100081 4 西藏那曲市草原站,那曲 852000
在干旱胁迫下植物形态结构、生理功能及生物化学过程受到抑制作用,如叶面积和叶厚度、叶片气孔导度、叶片净光合速率[1]受限。在植物个体水平上表达为植物功能性状的差异[2],植物通过采取不同的资源分配策略,以达到最高的生存能力[3],进而对生态系统关键过程产生不同影响。植物物种间差异体现在物种对不同生境的适应,即植物的功能性状[4]。与关键生态过程相关的性状被称为植物生活史的核心,包括植物个体大小、种子大小和叶组织结构(叶面积、干物质含量等),并对资源利用、植物再生和扩散密切相关并对干扰有显著响应[5—6]。功能多样性作为生物多样性的一个重要方面,是指可以影响生态系统功能的物种性状值和分布范围[7]。基于植物功能性状和群落功能多样性的研究,能更好地探知不同生境中植物面临的限制和机遇[8],预测植物群落组成[9—10],衡量功能多样性对生态系统功能影响的作用[6,8,11—12]。越来越多的研究表明,生态系统的功能如群落生产力、物质循环、能量流动、对干扰或入侵的恢复力[13—14]更依赖于群落功能多样性[2,15—18]。全面深入地了解植物功能多样性与生态系统过程或功能之间的关系,有助于更好地应对全球气候变化和人类活动干扰下生物多样性丧失的生态学后果。
被称为“第三极”的青藏高原作为地球上最高的生物地理单元,具有独特的水热条件和复杂特殊的地理环境,气候敏感并且生态环境脆弱[19—21],极易受到气候变化和人类活动的干扰。在全球变化背景下,降水格局发生季节性变化,青藏高原出现极端干旱事件的频率增加、强度加剧[22—23]。在草地生态系统中,干旱通常导致功能多样性降低[24],不同植物的适应干旱的策略不同,优势种多表现出忍耐干旱的策略,而稀有种多表现出躲避干旱的策略[24]。然而降水季节性的改变对植物功能多样性影响的研究较少,青藏高原高寒草甸植物功能多样性对不同生长季干旱的响应机制尚且不明。
通过模拟不同时期干旱事件,观测藏北高寒草甸群落优势种的盖度、选取并测定植物功能性状,基于功能性状和物种盖度计算群落功能多样性指数,并分析群落功能性状和功能多样性对不同时期干旱的响应情况,以阐明不同生长时期干旱条件下高寒草甸植物群落功能多样性对干旱胁迫的响应机制。为预测未来季节性干旱事件对青藏高原高寒草甸植物群落功能性状、群落特征和功能多样性的影响提供科学依据。
1 材料与方法
1.1 研究区概况
研究区地处念青唐古拉山以北的高寒草甸,位于西藏那曲市“国家农业环境那曲野外科学试验站”内,地理坐标为92°02′E,31°44′N(图1),平均海拔4500 m以上。根据国家气象科学数据中心[25],试验区近十年平均温0.3℃,生长季(5—9月)月平均温度高于0℃。年均降水量为450—500 mm,90%以上集中在生长季,平均土壤体积含水量约为22.86%,土壤质地为高山草甸土。研究区草地类型为寒冷湿润冻原、高山草甸类[26],主要以高山嵩草(Kobresiapygmaea(C. B. Clarke) C. B. Clarke)为建群种,常见种有藏豆(Hedysarumtibeticum(Bentham) B. H. Choi &H. Ohashi)、钉柱委陵菜(PotentillasaundersianaRoyle)、多裂委陵菜(PotentillamultifidaL.)、二裂委陵菜(PotentillabifurcaL.)、弱小火绒草(Leontopodiumpusillum(Beauv.) Hand.-Mazz.)等,植物返青期多在5月,植物枯黄期多在8—9月。
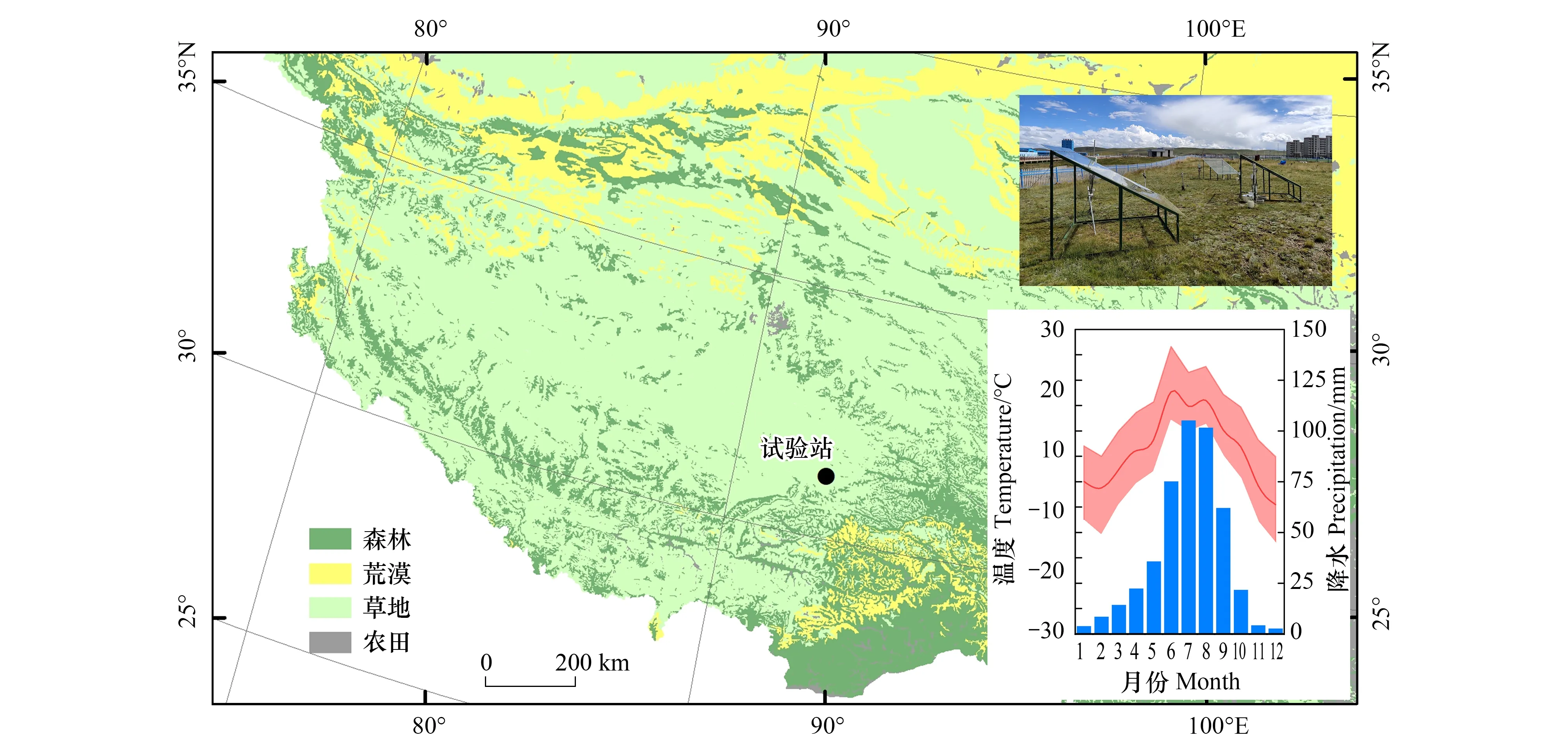
图1 研究区域图Fig.1 Experimental plot●为试验站地理位置,试验样地(右上),试验样地多年月平均降水量和气温情况(右下)。
1.2 试验设计
本试验自2016年起,通过设置截雨棚装置模拟高寒草甸在不同生长阶段的干旱事件[27],截雨棚为透光率90%的PC板,地面覆盖范围2 m×2 m的可移动装置(图1)。设置生长季前期(5—6月)干旱(Early growing season drought,ED)、生长季中期(7—8月)干旱(Middle growing season drought,MD)、非生长季(9月—次年4月)干旱(Non-growing season drought,ND)和对照未截雨(Control,CK)4个处理,4次重复,共16个子小区。每个处理小区边界土壤中垂直埋入地下深度为30 cm的隔水板,并在处理小区间设置2 m缓冲区。每个处理小区分成4个0.5 m×0.5 m的子小区,其一作为观测区,其三为破坏性采样区,并在小区中埋设有EM50自动记录器(Decagon公司,美国)监测土壤温湿度,干旱处理小区的土壤体积含水量在截雨期间显著降低[28]。
1.3 功能性状的测定
于2021年8月中旬生长季高峰期进行群落调查和样品采样,根据“质量比假说”选取优势种作为功能性状的观测对象[29—30],共选取高山嵩草、藏豆、钉柱委陵菜、多裂委陵菜、二裂委陵菜及弱小火绒草6个优势物种,各小区中优势种的分盖度总和平均占小区群落总盖度86.47%,最高达到95.11%。在各试验小区的采样子小区中根据“五点取样法”选取5—10株个体,沿叶柄基部剪下最新生长且完全展叶的10—20个叶片,由于高山嵩草叶片狭小,采样时选取20个叶片,其余5种植物采取10个叶片。取样后即刻装入密封袋中,保持叶片含水量。为研究不同时期干旱下高寒草甸植物的功能性状的差异,参考LEDA Traitbase数据库(www.leda-traitbase.org.)[2,8]对植物功能性状的描述和选定标准,测定了8项叶片功能性状:叶面积(LA:mm2)、比叶面积(叶面积与叶干重之比,SLA:mm2/mg)、叶干物质含量(叶干重与叶鲜重之比,LDMC: mg/mg)、叶碳含量(LCC:mg/mg)、叶氮含量(LNC:mg/mg)、叶磷含量(LPC:mg/mg)、叶碳氮含量比(C∶N:mg/mg)以及叶氮磷含量比(N∶P:mg/mg)。称取叶片鲜重,置于60℃烘箱中烘干48 h至恒重,称得干重,测量精确到0.01 g。叶面积通过对叶片扫描后,采用ImageJ(https://imagej.nih.gov/ij)软件测量。LCC和LNC采用有机元素分析仪(Flash EA1112,Thermo Scientific,USA)测定,LPC采用钒钼黄吸光光度法测定。
以优势种的相对盖度为权重,计算了每个功能性状的群落加权平均值(CWM),即LACWM、SLACWM、LDMCCWM、LCCCWM、LNCCWM、LPCCWM、C∶NCWM和N∶PCWM,以表征群落的功能性状。计算公式[31]:
式中,Pi为第i种盖度占总盖度的比例,n为物种数目,Xi是物种的特征值。
1.4 群落功能多样性
群落功能多样性由功能均匀度、功能分异度和功能分散度等指数描述,每个指数描述群落特征分布的不同方面[32],且对干旱的敏感程度及对生态过程影响存在差异。采用R-4.0.5软件的FD程序包中dbFD函数[33]计算了功能均匀度指数(FEve)、功能分异度(FDiv)、功能分散度(FDis)和Rao二次熵(RaoQ)指数[34—35],各指标计算公式(表1)。数据均通过Bartlett方差齐性检验符合方差齐性,同时计算各功能多样性指数的变异度。

表1 功能多样性指数各指标及其计算公式和生态学涵义
并根据以下公式计算各功能多样性指数的变异度:
ΔFDdp=FDdp-FDCK
式中,FD(functional diversity)代表功能多样性指数,包括FEve,FDiv,FDis和RaoQ指数,dp(drought periods)为干旱时期,包括ED、MD和ND。ΔFD(即ΔFEve、ΔFDiv、ΔFDis和ΔRaoQ)表示不同干旱时期下功能多样性指数与对照组CK相比的变异程度。
1.5 数据分析
采用R-4.0.5软件进行统计学分析及绘图,调用“FD”程序包中的dbFD函数计算功能多样性指数。数据采用单因素方差分析(ANOVA)和Duncan多重比较分析群落总盖度、优势物种相对盖度、优势种的功能性状、群落功能性状在各处理间的差异,采用变异度分析群落功能多样性对不同时期干旱处理的响应。并采用Pearson相关系数分析群落功能性状和功能多样性的相关性,探讨群落功能性状与功能多样性对干旱响应间的关系。
2 结果与分析
2.1 优势种盖度对不同时期干旱的响应
优势种的相对盖度对不同时期干旱的响应存在差异。在ED处理下各优势种相对盖度均显著降低(P<0.05),其中二裂委陵菜相对盖度最低,为(2.3±1.0)cm,与CK相比降低了66.8%。MD处理下仅多裂委陵菜与二裂委陵菜的相对盖度显著降低(P<0.05),分别为(5.7±1.8)cm和(3.8±0.4)cm,比CK降低了 49.0%和44.5%。ND处理对各优势种的相对盖度均没有显著影响(图2)。ED和MD处理导致植物群落总盖度显著降低(P<0.05)(图3),分别为(59.2±1.7)cm和(67.0±9.8)cm。ED处理对优势种相对盖度的影最大,使其下降了37.6%,MD处理降低了优势种相对盖度的29.5%。
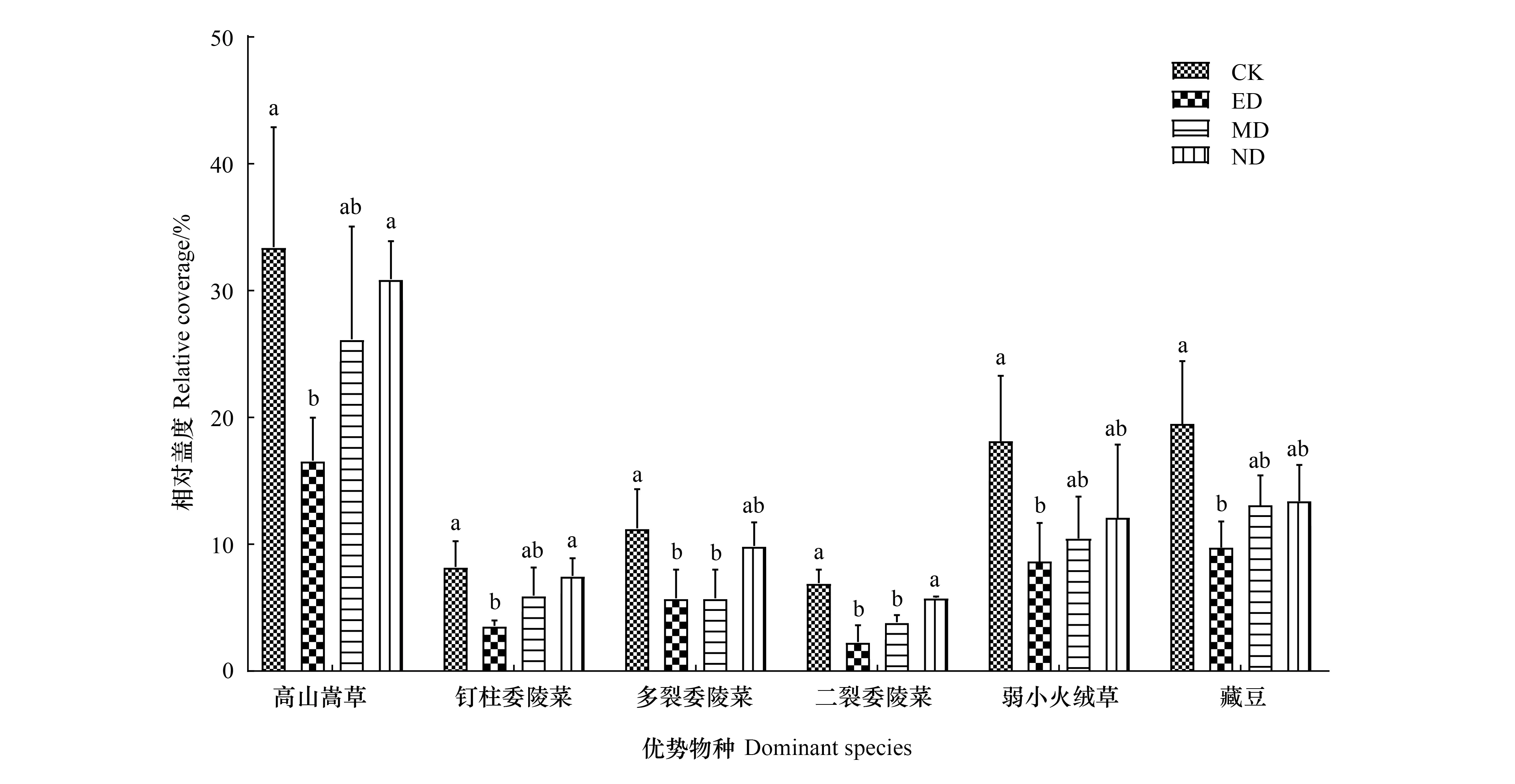
图2 不同时期干旱处理下6个优势种的相对盖度Fig.2 Relative coverage of 6 dominant species under drought treatment in different timing不同小写字母代表组间差异显著,显著性水平P<0.05;图中数据为平均值±标准差;CK对照;ED生长前期干旱;MD生长中期干旱;ND非生长时期干旱
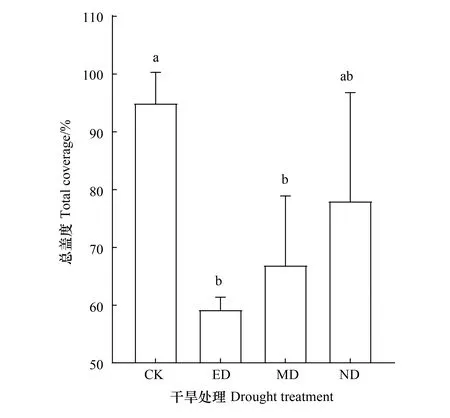
图3 不同时期干旱处理下群落总盖度 Fig.3 Total coverage under drought treatment in different timing 不同小写字母代表组间差异显著,显著性水平P<0.05;图中数据为平均值±标准差
2.2 优势种功能性状对不同时期干旱的响应
植物各功能性状对干旱胁迫产生差异化表达,ED和MD处理对优势种功能性状的影响较ND更显著。ED和MD处理显著降低了藏豆、多裂委陵菜和弱小火绒草的LA,而钉柱委陵菜的LA仅在ED处理下显著降低(表2)。ED、MD和ND处理下高山嵩草SLA显著降低,ED和MD处理下藏豆SLA显著降低,而弱小火绒草的SLA仅在MD处理下显著降低(P<0.05)。ED处理下高寒草甸、多裂委陵菜和二裂委陵菜的LDMC显著升高,而弱小委陵菜在ED和MD处理下均显著增加;MD处理下藏豆、钉柱委陵菜和多裂委陵菜的LCC显著增加,而二裂委陵菜在ED和MD处理下显著升高(P<0.05)。藏豆和二裂委陵菜的LNC在ED、MD和ND处理下均显著降低,高山嵩草、多裂委陵菜在ED和MD处理下显著降低(P<0.05)。在MD处理下,藏豆、钉柱委陵菜、二裂委陵菜和弱小火绒草的LPC均显著降低,而高山嵩草在ED和MD处理下均显著降低(P<0.05)。对于优势种的C∶N,在ED和ND处理下高山嵩草显著降低,ED和MD处理下多裂委陵菜显著升高,而二裂委陵菜在ED、MD和ND 处理下均显著升高(P<0.05)。在ED、MD和ND处理下,高寒草甸的N∶P显著升高,而ED和MD处理下多裂委陵菜的N∶P显著降低,ED和ND处理下二裂委陵菜N∶P显著降低,而藏豆仅在MD处理下显著升高(P<0.05)。
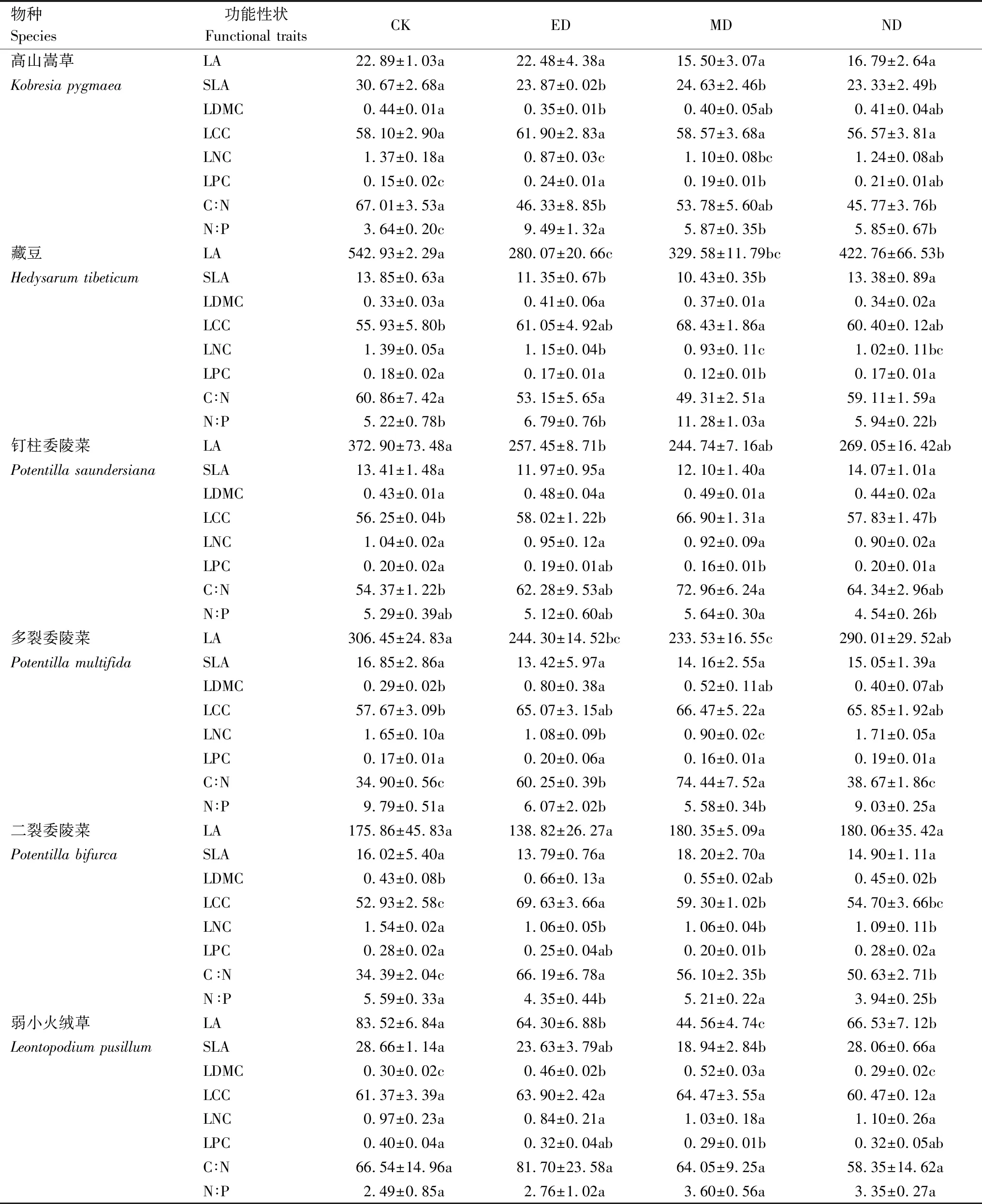
表2 不同生长时期干旱处理下植物功能性状的多重比较结果
2.3 群落功能性状对不同时期干旱的响应
各群落功能性状多受干旱胁迫抑制,仅群落叶干物质含量和叶碳氮比表现出了正向响应(图4)。在ED和MD干旱处理下LACWM、SLACWM、LNCCWM、LPCCWM和N∶PCWM均显著低于CK(P<0.05),且ED处理对其的影响更强。在ND处理下仅LACWM和LPCCWM显著低于对照(P<0.05),但仍高于ED和MD处理。同时干旱处理显著增加了群落的LDMCCWM(P<0.05),且ED处理下LDMCCWM高于MD和ND处理(图4)。各时期干旱处理下群落LCCCWM均与CK无显著差异(图4)。群落C∶NCWM仅在ED处理下显著升高(图4)。
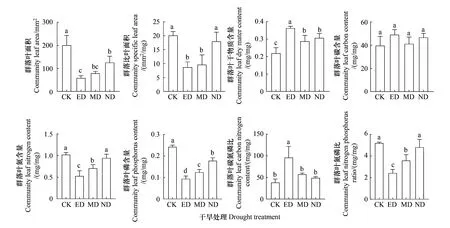
图4 群落功能性状对不同时期干旱的响应Fig.4 Responses of community functional traits to drought in different timing不同小写字母代表组间差异显著,显著性水平P<0.05;图中数据为平均值±标准差
2.4 群落功能多样性对不同时期干旱的响应
不同时期干旱处理显著改变了群落的功能多样性。对照处理下各功能多样性指数分别为,FEve指数为0.77±0.04,FDiv指数为0.88±0.01,FDis指数为2.44±0.10以及RaoQ指数为6.40±0.36(图5)。FEve指数在MD处理下显著低于CK(P<0.05),与CK的差值最低值达到-0.12(图5)。ED处理下FDis、FDiv指数均有显著低于CK(P<0.05),与CK的差值最低分别达到-0.33和-0.12(图5)。ED处理下RaoQ指数显著高于CK(P<0.05),与CK的差值最高达到1.64(图5)。ND处理下各群落功能多样性均与CK无显著差异。
2.5 群落功能性状与功能多样性对干旱响应的相关性
各群落功能性状的变化对FDiv、FDis以及RaoQ指数改变的相关性较为一致,而与ΔFEve的相关性存在差异(图6)。ΔFEve与ΔLACWM(r=0.81,P<0.01)和ΔLPCCWM(r=0.75,P<0.05)呈显著正相关关系。而ΔFDiv、ΔFDis以及ΔRaoQ与ΔLDMCCWM、ΔLCCCWM和ΔC∶NCWM的响应呈正相关关系,其中ΔLDMCCWM与ΔFDiv的影响呈显著正相关(P<0.05),ΔLCCCWM和ΔC∶NCWM与ΔFDiv、ΔFDis以及ΔRaoQ之间均显著正相关。而ΔFDiv、ΔFDis和ΔRaoQ与ΔLNCCWM、ΔLPCCWM、ΔN∶PCWM呈负相关关系,其中ΔFDiv与ΔLNCCWM(r=-0.83,P<0.01)、ΔLPCCWM(r=-0.73,P<0.05)以及ΔN∶PCWM(r=-0.69,P<0.05)呈显著负相关,ΔFDis和ΔRaoQ与ΔN∶PCWM呈显著负相关(r=-0.73和r=-0.71,P<0.05)。
3 讨论
3.1 高寒草甸功能性状对不同时期干旱的响应
藏北高寒草甸各植物功能性状对干旱胁迫存在差异化响应。中高寒草甸优势种的SLA、LA在生长季前期和中期干旱处理下显著降低,其原因是干旱胁迫破坏了植物水分吸收和蒸腾作用间的动态平衡,导致植物叶片气孔关闭、叶绿素含量降低,蒸腾速率和光合作用活力降低,CO2同化过程受到抑制,干物质积累受限[30],生产力下降的同时引起SLA、LA的降低。主要是土壤水分不足以维持植物蒸腾需要时,叶片萎蔫甚至凋落,以减少水分散失[36],更多的同化物分配到根系生长中。在个体表型上,植物叶片趋向于变小变厚以适应干旱胁迫[30]。而叶片薄而富含氮的植物具有高转化率和高生物量同化率,即SLA、LA、LNC和LPC等功能性状高时,植物具有高生产力和高竞争力[30]。干旱处理下,多裂委陵菜、二裂委陵菜和弱小火绒草的LDMC显著增加,同时藏豆、多裂委陵菜和钉柱委陵菜的LCC显著增加。LDMC和LCC表征植物叶片组织的密度,反映植物竞争资源的能力[37],植物叶片LDMC和LCC高时,叶片厚且寿命长,植物具有较低的生物量同化速率,即低生产力和低竞争力[30]。干旱胁迫使得高寒草甸优势种叶片形态趋向小而厚的同时叶片干物质含量升高,表明高寒草甸植物通过降低生物量同化速率提升对干旱胁迫的抵御能力[37—38]。
叶片中N和P元素作为光合作用酶和生物能量分子的必需元素,其浓度与植物净光合速率密切相关。高寒草甸群落功能性状LNC和LPC在生长季前期和中期干旱处理下显著降低,Wright提出的“叶片经济波谱”中认为植物LNC、LPC和SLA与光合速率彼此之间呈显著正相关,低SLA的植物具有低光捕获率和净光合速率[39]。干旱胁迫限制了高寒草甸植物的净光合速率,高寒草甸群落植物通过调节叶片氮磷含量使其处于较低的相对生长速率以维持在干旱逆境中的生长。同时LNC和LPC共同反映植物体在生态系统中受N限制或P限制的养分供应情况,对照处理下群落N∶P为5.8±2.2,N∶P<10表征植物生长更大程度上受到N元素的限制[39],且在生长季前期和中期干旱处理下N∶P显著降低,指示着高寒草甸群落在干旱胁迫下N限制加剧。植物碳氮比演化机制研究中提出的“适应性生长假说”指出在强烈N限制环境中,维持高C∶N有助于植物提高养分利用效率从而优先保证植物生存[40], 高寒草甸群落C∶N在生长季前期干旱处理下显著升高,故高寒草甸植物在干旱胁迫下通过提高养分利用效率以维持生存。
3.2 高寒草甸功能多样性对不同时期干旱的响应
高寒草甸植物群落功能多样性对干旱事件的响应可能存在多种调控机制,即植物群落采取不同的干旱策略。以往研究认为,采取耐旱策略的优势种在群落中占主导地位时,往往使功能多样性在响应干旱时下降[18—19,34]。这与高寒草甸植物群落对生长季前期干旱的响应相一致,FDis、FDiv显著降低,即群落中物种间功能离散程度和物种特征值差异性降低,群落物种生态位的分化和资源利用程度降低;而RaoQ指数呈显著升高,群落物种间的差异度加大。表明高寒草甸植物群落在响应生长季前期干旱时可能更倾向于耐旱策略。此外,干旱胁迫下生态位的分化和物种共存机制却能促使功能多样性增加[41],群落应对干旱的策略倾向于避旱型[34]。生长季中期干旱处理下群落FEve多样性显著降低,即群落功能特征在空间中丰度分布的均匀性降低[34],高寒草甸植物群落在响应生长季中期干旱时表现出避旱型策略。
3.3 群落功能性状与功能多样性对干旱的响应
高寒草甸植物群落应对干旱的策略与不同功能性状对干旱的响应密不可分。前期研究发现地中海生态系统中群落功能性状通过影响功能多样性进而作用于生态系统的多功能性[42],旱地群落内的功能多样性改善了生态系统的多功能性。此外,植物功能多样性驱动了黄土高原植物群落的碳存储功能[43]。FDiv、FDis和RaoQ指数与群落植物的LDMC、LCC、LNC和LPC相关。可见群落功能性状如群落植物叶片的干物质积累情况、叶碳氮磷元素计量,通过影响功能多样性作用于生态过程。研究表明群落中具有低SLA和(或)LNC的植物多采取保守型生存策略即耐旱型,以增加在资源有限如干旱胁迫[11,44]的环境中持续存在的能力;而高SLA和LNC的植物则多采取避旱生存策略[30]。 高寒草甸群落应对生长季前期干旱的耐旱策略与植物群落的物质积累和养分循环有密切联系,植物通过提高养分利用效率忍耐干旱[41]。而功能均匀度指数主要与群落植物的叶面积和叶氮含量相关,高寒草甸群落对生长季中期干旱的避旱策略与植物形态性状和P元素利用有关。
4 结论
在干旱胁迫下高寒草甸群落植物叶片功能性状表现为叶片小而厚且寿命长,同化速率降低,并且受氮元素限制加剧。结果中,ED干旱胁迫对群落功能性状的抑制最强;ED和MD处理显著改变了群落的功能多样性,ED处理下群落物种间功能离散程度和物种特征值差异性降低,MD处理下群落功能特征在空间中丰度分布的均匀性降低。功能性状与群落功能多样性对ED和MD干旱存在响应差异,表明不同时期干旱可能采取不同的生存策略,即群落对生长季前期干旱采用耐旱策略、对生长季中期干旱采用避旱策略。探讨了高寒草甸植物群落功能多样性对不同生长时期干旱胁迫的响应情况,为预测未来季节性干旱事件对青藏高原高寒草甸植物群落功能性状、群落特征和功能多样性的影响提供科学依据。
猜你喜欢
疯狂英语·新读写(2023年4期)2023-05-10
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
家庭医药(2021年12期)2021-12-09
家庭医药(2021年23期)2021-10-21
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
活力(2019年21期)2019-04-01
防护林科技(2015年5期)2015-06-10
海南热带海洋学院学报(2015年5期)2015-03-14
