
基于生长测量仪监测的亚热带地区马尾松多时间尺度径向变化及其与环境因子的关系
2023-09-11 07:26:14曾林辉迟永刚
生态学报 2023年16期
曾林辉,周 蕾,*,寇 亮,迟永刚
1 浙江师范大学地理与环境科学学院,金华 321004 2 中国科学院地理科学与资源研究所,生态系统网络观测与模拟重点实验室, 北京 100101
森林能够吸收温室气体和防风固沙,为人类提供木材以及为动物提供栖息地[1]。树干径向变化是林业研究的重要内容,对人类的生产生活具有极其重要的意义[2]。全球气候变化背景下,气温的逐年上升和降水模式的改变对树干径向变化的影响也在增强[2—5]。例如,适当的温度升高对树干的径向变化具有促进作用,然而温度过高会对树木产生干旱胁迫[6];适当的降水会促进树干的径向变化,然而过多的降水会抑制树木生长,甚至造成树木死亡[7]。因此,研究树干径向变化及其对气候变化的响应,有助于准确预测森林生态系统生产力如何应对气候变化,有利于量化评估未来森林生态系统的动态变化。
目前国内外研究树干径向变化的方法很多[8—9]。例如树木年轮法主要适用于年际尺度研究,缺乏树木生长的生理基础[9—12]。结合组织解剖学的微树芯法和针刺创伤法,能够从植物内部(细胞层面)揭示较短时间尺度上的树干木质部生长发育变化及其对环境因子的响应[13—16]。上述这些方法都会在不同程度上损伤树体,并且缺乏连续高时间分辨率的树干径向变化,无法准确分析树干径向变化的生理机制和气候驱动力[9—10,17]。高分辨率树干径向变化记录仪(Dendrometer)能够完整连续地监测树干径向变化,提供了树木径向从日尺度到年尺度的变化,准确地描述环境因子对树干径向变化促进或抑制作用[9, 17—19]。Dendrometer测得的树干径向变化信息既包含了树干的生长信息,也包含了树干因水分的消耗和补充而发生的变化情况,所以监测短时间尺度树干径向变化也能够在一定程度上补充和解释长时间尺度关于树干径向生长研究结果[20—21]。
目前已有很多研究利用Dendrometer在全球热带,温带,寒带等地区开展树木日尺度和季节尺度的径向变化研究。在日尺度研究中,杨艳刚等(2009)在我国山西芦芽山(温带地区)研究白杄树干的日径向变化过程,发现日径向变化包括树干真正的加粗生长和木质部含水量变化导致的膨胀和收缩变化,并将一日内的变化分为收缩、恢复、增长三个阶段[22];Rezaei等(2022)在德国马夸特(温带地区)研究苹果树日径向变化时发现在日收缩阶段,收缩持续时间随着饱和水汽压亏缺(VPD)的增加而增加,日收缩量与 VPD的关系在采收后强于采收前[23]。在季节尺度研究中,Biondi等(2010)在墨西哥热带地区研究山松年际生长时发现太阳辐射最大的5月份是山松生长快速期,并且土壤温度升高可以促进山松径向生长[24];Miller等(2022)在德国西南部四个海拔梯度上研究温带地区道格拉斯冷杉(Douglasfirs)、挪威云杉(Norwayspruces)和银冷杉(Silverfirs)径向生长动态时发现干旱会影响树干生长开始时间,干旱年份生长开始时间明显早于正常年份[25]。但是,目前亚热带地区的树干日尺度、季节尺度下的径向动态变化及其对环境的响应的研究则相对较少。
我国南方亚热带森林生态系统在全球碳水循环过程中发挥着至关重要的作用[26—27],有研究数据证实我国东亚季风区亚热带森林净生态系统生产力总量约为每年7.2亿吨碳,约占全球的8%[28—29]。马尾松(PinusMassoniana)是我国亚热带地区广泛分布的主要造林和当地本土树种[30—31],因其在营养贫乏的土壤中的具有高生存能力和快速生长能力[26, 31],在植被和土壤恢复中发挥了关键作用。高时间分辨率的马尾松树干径向变化以及对环境因子的响应的研究,对于气候变化下的植树造林设计和可持续森林管理至关重要[31]。本研究以马尾松作为研究树种,利用高分辨率树干径向变化记录仪监测马尾松树干径向变化动态及其对气象要素的响应。主要研究目标是:1)揭示亚热带地区马尾松树干径向昼夜变化特征;2)确定马尾松年内季节尺度下树干径向变化周期和生长动态;3) 明确影响日尺度和季节尺度上马尾松树干径向变化的主要环境因子。
1 材料与方法
1.1 研究区概况
研究区位于江西省泰和县中国生态系统研究网络千烟洲森林站内(26°44′N,115°44′E,102 m)。该站地处江西省中部吉泰盆地,属于典型的亚热带季风气候。年均温度为17.9℃,年均降水量为1485.1 mm,年总太阳辐射为4661 MJ/m2,年均相对湿度为84%,年无霜期323 d[32]。千烟洲属于典型的红壤丘陵地貌,成土母质多为红色。实验站内森林覆盖率高达90%,优势树种为马尾松、杉木和湿地松等[33]。
1.2 数据获取
随机选取研究区通量塔附近4棵胸径、树高和生长状况等方面特征相似的马尾松(Pinusmassoniana)树作为实验对象,分别在每棵马尾松胸径1.3 m处安装树干径向变化记录仪(DC3, Ecomatik, 德国)。为减少树皮膨胀和收缩对探头压力的影响,在安装前要先去除安装部位的树皮。记录仪由传感器(DC3)和数据采集器(UX120-006M)两部分组成。工作原理是树干对探头的压力在膨胀与收缩期间发生连续变化,压力的变化会引发传感器电阻的变化;数据采集器记录电阻的变化信息,并将其转化为周长长度的变化。传感器设置数据采集时间间隔为30分钟。数据采集时间范围为2018年9月至2019年9月,起始时间从2018年9月1日0时开始。导出的电压值经过计算得到树干周长值,文中的径向变化均指周长变化。
气象数据来源于千烟洲试验站综合气象观测场,选择的环境因子包括空气温度(Air Temperature,AT)、相对湿度(Relative humidity,RH)、降水量 (Precipitation,Prec) 。土壤温度(Soil Temperature,ST) 由林内小型自动气象站记录,观测深度为40 cm。气象数据均为每日每小时的观测值。根据空气温度和相对湿度计算得出VPD[34]:
VPD = (1-RH) × 0.6108 × e(17.27AT/(AT+273.3))
(1)
1.3 数据分析
在马尾松树干径向日动态的研究中,计算马尾松日径向变化(即昼夜变化)来表示每个月径向变化在白天和夜晚的差异。具体计算方法是把每月每日30 min间隔的径向变化值平均得到月平均径向昼夜变化,所得到的图形类似于正弦波[35—36]。根据每条正弦波,获得每月平均日变化的最大值和最小值,以及它们的发生时间段。气象数据则将每个月每小时的气象数据平均后得到月平均每小时气象数据。
在马尾松径向季节动态研究中,计算马尾松径向日变化值来表示树干径向变化每天的差异。计算方法采用Achim等提出的日最大值法[37],即从每日的48个数据中提取出最大值,做连续两日的最大值之差以作为径向日变化值。径向日变化值的累积加和得到2018年9月到2019年9月的13个月的径向日累积变化值(首日即2018年9月1日变化值为0),4棵马尾松径向日累积变化值的平均值得到平均径向日累积变化值。本研究选取Gompertz方程[24, 38—39]拟合平均径向日累积变化值得到平均累积变化曲线,模拟出该时期马尾松径向生长过程。因Gompertz方程具有灵活性和不对称形状,被认为是可用于描述整个生命周期的树木生长模式以及季节性生长的各种 sigmoidal 模型中是最合适的模型[18, 24, 39—40]。Gompertz方程公式如下:
Y=Y0+Aexp(-eβ-kt)
(2)
式中,Y表示日径向累积增长量;Y0是下渐近线;A为上渐近线;β为x轴的截距参数;K为变化速率参数,其大小会影响生长季的长短;t是以天为单位的时间。拟合方程进行求导得出马尾松该时期的生长速率函数(mm/d)。当生长速率超过或低于0.02 mm/d的时间即为生长开始和停止的时间[41—43]。在确定生长开始和停止的时间之后,主要生长季节的时间则通过生长期内每个月的增长量确定,平均月增长量大于0的月份即为主要生长季[20, 31]。通过分析马尾松月平均增长量、平均径向日累积变化及日生长速率可将年内径向变化大致分为四个时期,并对主要生长季内的三个时期进行径向变化与环境因子的相关性分析和回归分析。环境因子数据则平均24小时气象数据得到日平均气象值。
为了消除个体的生长趋势差异来分析树木对外部环境的敏感性,在日尺度和季节尺度的树干径向变化与环境因子的关系研究中均使用了最小-最大归一化(0—1归一化)方法,避免不同绝对增长率的影响[31]。运用Pearson相关分析法计算主要生长季节内各生长时期的标准化日径向变化及径向日变化与环境因子之间的相关性,明确日尺度和季节尺度各生长期树干径向变化的主要影响因子。具体来说,在研究日尺度时取标准化后的马尾松生长期内日径向变化为因变量,在研究季节尺度时取标准化后的径向日变化为因变量,同时期内的环境因子为自变量,采用Pearson分析方法做相关性分析。此外,为分析各生长时期环境因子对马尾松径向生长的贡献程度,本研究在季节尺度上以环境因子为自变量,径向变化为因变量进行逐步回归分析。数据统计分析使用SPSS 23,作图使用Origin 2019b。
2 结果
2.1 环境因子的日动态和季节动态
空气温度和VPD在一天内变化呈现波峰曲线,最大值出现在14:00—15:00,其变化幅度从冬季到来年夏季越来越大(图1);相对湿度在一天内变化呈现波谷曲线,最小值出现在15:00—16:00,同样变化幅度从冬季到夏季越来越大。土壤温度一天内的变化无论冬夏季都相对较小,降水量一天内的变化从冬季到夏季无明显规律。

图1 千烟洲地区2018.9—2019.9空气温度、相对湿度、降水、土壤温度和水汽压亏缺因子日动态变化Fig.1 Daily variation of air temperature, relative humidity, precipitation, soil temperature and vapor pressure deficit over the period 2018.9—2019.9 in Qianyanzhou station
研究站点日平均气温为18.8℃,月平均温度从6.3℃(2018年12月)增加到29.6℃(2019年7月),19年6月和7月的日最高温度都超过38℃(图2)。土壤温度与空气温度相比,日振幅较小,日平均土壤温度为21.2℃,最低月均温(2019年1月)为10.9℃,最高月均温(2019年8月)为29.7℃。年降水总量为1773.9 mm,除在11月15日降水量剧增外,降水从3月开始增多,主要集中在3月、6月和7月。相对湿度集中在58%—100%之间,饱和水气压亏缺VPD全年变化在0.6—1.3 kPa之间。
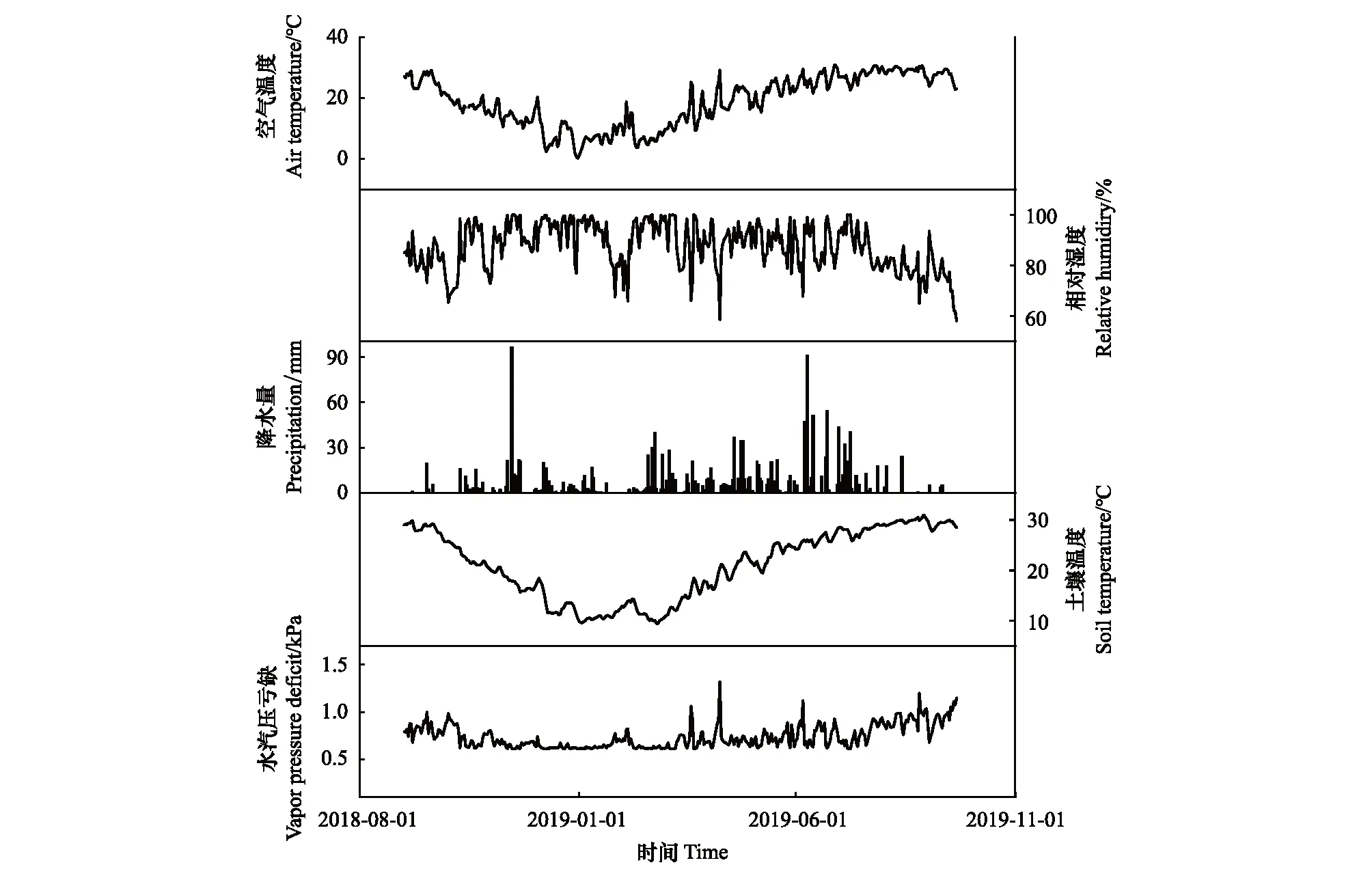
图2 千烟洲地区2018.9—2019.9空气温度、相对湿度、降水、土壤温度和水汽压亏缺因子季节动态变化Fig.2 Seasonal variation of air temperature, relative humidity, precipitation, soil temperature and vapor pressure deficit radiation over the 2018.9—2019.9 in Qianyanzhou,Jiangxi
2.2 马尾松树干径向变化的日动态
马尾松树干径向的昼夜变化特征表现出明显的季节特性(图3)。全年日径向变化幅度在0.01—0.06 mm之间。从3月开始日径向变化变化幅度开始变大,到9月日径向变化变化幅度开始缩小,至12月全天日径向变化趋于0。从全年看,日径向变化幅度在夏季(7月)最大,在冬季(12月)最小。然而,与变化幅度的季节性不同,日径向变化的周期性没有明显的季节性差异,全年的日变化最小值出现在12:00—14:00;日变化最大值出现在夏季的19:00—20:00,在冬季的8:00—9:00。

图3 2018年9月—2019年9月马尾松(Masson pine)月平均日径向昼夜变化模式 Fig.3 The mean daily radial diurnal variation pattern of Masson pine from September 2018 to September 2019
2.3 马尾松树干径向变化的季节动态
虽然不同样本树的年累计变化量存在差异,但径向生长变化模式高度相似(图4)。马尾松年累计生长量范围是8—10 mm,平均年累计生长量在9 mm左右。树干在秋冬时节累计生长曲线呈现先平稳波动后下降的趋势,在12月至来年2月初这个时间段开始下降,平均累计收缩量最大2.5 mm左右;从2月开始上升,特别是进入春季累积生长量迅速增加,在春夏交替时期,累积生长曲线呈现出上陡坡形状;进入夏季后,累积生长量增长变缓,曲线呈现出稳定的缓慢升高态势。
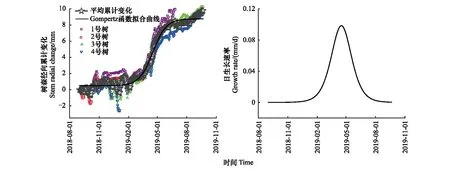
图4 2018年9月—2019年9月Gompertz 函数模拟的马尾松(Masson pine)径向生长变化及生长速率Fig.4 The radial growth change and growth rate of Masson pine from 2018.9 to 2019.9 simulated by Gompertz function
马尾松生长速率超过0.02 mm/d的时间在2月中旬,即为开始生长时间;低于0.02 mm/d的时间在9月初,即为停止生长时间(图4);生长速率在4月末5月初达到最大值(图4)。从图5可知马尾松在生长时期内每个月的平均径向生长增长量,发现3—8月的平均增长量都在0以上,所以马尾松主要生长季节在3—8月;4月的增长量最高,累计增长超过2 mm。

图5 马尾松(Masson pine)生长时期内各月份生长量 Fig.5 The growth of Masson pine in each month of the main growing season
通过分析树干13个月的月平均增长量、平均径向日累积变化以及日生长速率(图4和5),马尾松径向日变化大致分为4个时期:(1)树干开始生长期(2月中旬至3月底):该阶段树干各组织水分恢复,树干开始波动增长;(2)树干快速生长期(4月初至5月底):树干增长速度最快,进入持续稳定快速的生长时期;(3)树干缓慢增长期(6月初至8月底):树干增长速度变缓,树木趋向于稳定的状态,但树木仍处于生长状态;(4)树干收缩期(9月至来年2月):树木生长逐渐停止,树干为抵御冬天严寒开始收缩。
2.4 马尾松树干径向变化与气候特征的关系
表1显示了各个生长阶段马尾松日径向变化与环境因子的关系。研究表明在开始生长期内相对湿度(r=0.803,P<0.01)、降水量(r=0.426,P<0.01)、土壤温度(r=0.316,P<0.01)与马尾松日径向变化呈现正相关;而空气温度(r=-0.315,P<0.01)和VPD(r=-0.750,P<0.01)与马尾松日径向变化呈现负相关。在快速生长期内,马尾松径向昼夜变化与相对湿度(r=0.846,P<0.01)和土壤温度(r=0.283,P<0.05)呈正相关,与VPD(r=-0.845,P<0.01)呈负相关,与空气温度和降水量都不相关(P>0.05)。在缓慢增长期内,相对湿度(r=0.632,P<0.01)和土壤温度(r=0.247,P<0.01)与马尾松日径向变化呈现正相关,空气温度(r=-0.659,P<0.01)和VPD(r=-0.666,P<0.01)与马尾松日径向变化呈现负相关。从整个生长期上看,马尾松径向昼夜变化与相对湿度和土壤温度呈正相关,相对湿度的正相关性强于土壤温度;与 VPD和空气温度呈负相关,VPD的负相关性强于空气温度。

表1 马尾松在主要生长季内各生长时期日径向变化量与环境因子的Pearson相关关系分析
表2显示了各个生长阶段马尾松径向日变化与环境因子的关系。研究表明在开始生长期内主要影响因子是空气温度和土壤温度,并且土壤温度与径向日变化的相关性(r=0.316,P<0.01)强于空气温度(r=0.211,P<0.05)。在快速生长期内,径向日变化与降水量和空气温度呈正相关关系,降水量的相关系数(r=0.251,P<0.05)高于空气温度(r=0.161,P<0.05)。缓慢增长期内相对湿度、空气温度、土壤温度和VPD均影响马尾松径向生长。径向日变化与相对湿度(r=0.315,P<0.01)呈显著正相关,与空气温度(r=-0.260,P<0.01)、土壤温度(r=-0.412,P<0.01)和VPD(r=-0.285,P<0.01)呈负相关,土壤温度的负相关性强于空气温度。
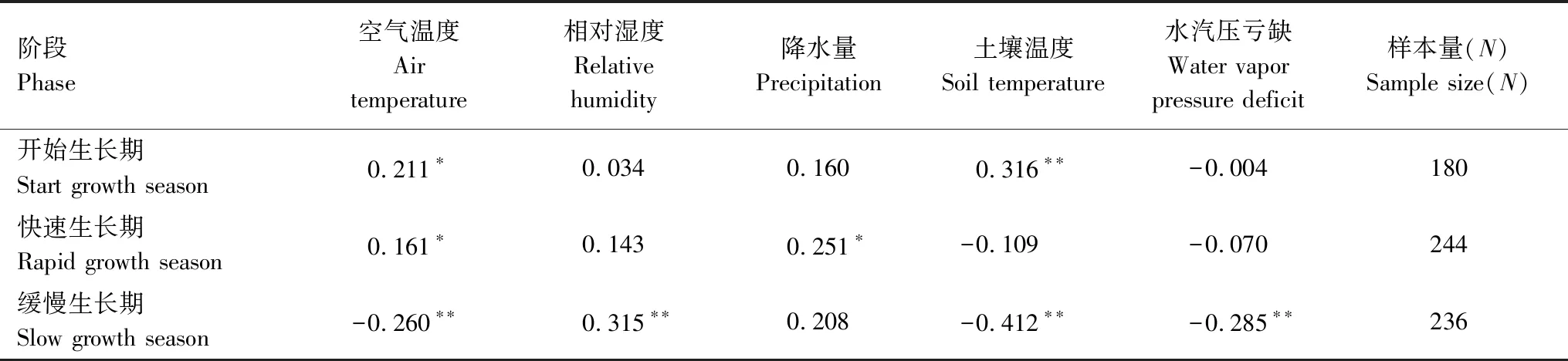
表2 马尾松在主要生长季内各生长时期径向日变化量与环境因子的Pearson相关关系分析
综合整个生长时期,空气温度与径向日变化的相关系数都不高,降水量只对快速生长期的径向日变化有影响,VPD只对缓慢生长期的径向日变化有影响,然而径向日变化对土壤温度和相对湿度的响应与其他环境因子有所不同。土壤温度在三个时期都对马尾松生长有影响,但是影响不同,从开始生长期与径向日变化正相关,到快速生长期与径向日变化不相关,最后在缓慢生长期与径向日变化呈现负相关;而相对湿度则从前两个时期与径向日变化不相关,转变成缓慢生长期内促进树干生长的主要影响因子(图6)。

图6 2018年9月—2019年9月马尾松(Masson pine)径向日变化量与土壤温度和相对湿度的关系Fig.6 Scatter plot of the radial diurnal variation of Masson pine with soil temperature and relative humidity
利用逐步回归分析建立回归模型,分析各生长时期环境因子对马尾松径向生长的贡献程度。结果显示:在开始生长期,土壤温度和空气温度对马尾松径向生长都起到促进作用,土壤温度的促进作用强于空气温度;在快速生长期,空气温度和降水量对马尾松径向生长都起到促进作用,降水量的促进作用强于空气温度;在缓慢增长期内,土壤温度和空气温度对马尾松径向生长都起到抑制作用,土壤温度的抑制作用强于空气温度。
开始生长期:
Y=15.131+0.461×ST+ 0.299×AT(P<0.05)
(3)
快速生长期:Y=14.934+0.286×Prec+0.175×AT(P<0.05)
(4)
缓慢增长期:Y=8.339-0.412×ST-0.300×AT(P<0.01)
(5)
3 讨论
3.1 马尾松树干日径向变化(昼夜变化)特征及其与环境因子的关系
本研究发现马尾松日径向变化在全年都呈白天收缩夜晚膨胀模式,但是秋冬季夜晚膨胀没有春夏季明显(图3),研究结果与我国同在亚热带气候下的江西杉木(Cunninghamialanceolata)和广西红椎(Castanopsishystrix)日生长模式大致相同[36, 44]。树干日径向变化主要由形成层生长和水分变化引起的[45—47]。形成层生长受多因素的影响,温度影响植物体内淀粉和脂质等的转化,调节细胞壁形成,降水影响细胞水势,为细胞的扩大提供动力[45—47];水分变化主要由土壤可利用水、树木蒸腾作用以及降水所引起的水势梯度驱动[47]。
在春夏季,水分变化是影响马尾松树干径向昼夜变化的主要气候因子(表1)。从春季开始,土壤温度上升,降水量增多,相对湿度加大,在夜晚根茎吸收水分充分,形成层细胞快速分裂,树木茎干膨胀。日出后,气温和辐射逐渐增加,湿度降低,树木冠层发生蒸腾作用的损失量小于根茎部的吸水量,因此上午十点前树干周长仍处于增加的状态[48—49]。从中午开始至傍晚这段时间内,蒸腾作用最为强烈,温度的升高和湿度的降低使得土壤水分减少,导致根部吸水量远达不到树冠需水量,茎干出现收缩现象[50—52]。傍晚之后,蒸腾作用减弱,温度降低,湿度回升,根部所吸收的水分增多且几乎都用于茎干组织,导致树干膨胀且膨胀量为全天最大[34, 53]。可见,降水增多和相对湿度加大是导致春夏季树干昼夜变化的主要因子。
很多研究表明进入秋冬时节,树木停止生长进入收缩期,日径向变化主要驱动因素不再是蒸腾作用引起的水分变化,而是温度[20, 35,48—49]。原因在于冬季夜晚温度低于树液的凝固点,细胞内自由水外流,导致树干收缩,而当白天温度升高且高于凝固点,水分回流细胞内,树干膨胀[35, 50—51, 54]。然而本研究中千烟洲冬季温度很少低于零度(图1),所以仍是水分变化驱动树干日径向变化(表1)。这个时期蒸腾作用不及夏季明显,且温度偏低,植物新陈代谢速率随之降低[36, 44]。从全年日径向变化幅度看,夏季振幅远大于冬季,在7月达到最大振幅,此结果与很多研究一致[35, 36, 48]。
3.2 马尾松树干径向变化的季节特征及其与环境因子的关系
本研究发现,马尾松径向日变化呈现出明显的季节性特征,可以分成生长,稳定和收缩3个阶段(图4)。在生长和稳定阶段,马尾松树干径向生长主要受温度(土壤温度和空气温度)与水分(湿度和降水量)的影响,但是其影响程度在各个阶段有所不同(表2)。
在开始生长期里(2—3月),无论是空气温度还是土壤温度都对树干生长有促进作用(表2),这与春季升温是生长季开始主要影响因子这一被广泛认可的结论一致[18, 22, 55]。开始生长期前马尾松因为冬季低温,出现了强烈的脱水收缩现象[56—57]。2—3月份亚热带地区气温回暖,土壤温度达到根系吸收水分的温度条件,根系吸水量增大,茎干水分恢复,树干开始生长[31,58—59]。此时的空气温度达到树木生长有效积温,促进树干生长[31, 58—59]。
在快速生长期(4月初至5月底),千烟洲空气温度上升同时由于梅雨季节降水持续增多(图2),空气温度和降水共同促进树干生长(表2)。4月份开始降水增多,土壤含水量随之增加,在温度的共同作用下根系吸收量比开始生长期更多,树干所需水分充盈,满足生长所需的环境条件,产生大量生长激素,形成层细胞新陈代谢加速,细胞持续膨胀增大和分裂[23, 31,60—61]。树干进入持续稳定快速增长时期,到4月底5月初各方面条件都达到最适宜值,树干径向生长速率达到最大(图4),其中4月树干生长迅速增粗,成为全年累计生长量最大的月份(图5)。这一研究结果与我国其他气候地区的树木生长研究存在差异。例如在暖温带半干旱半湿润地区,江源、杨艳刚等(2009)研究发现白杄和华北落叶松7月份树干径向增长量最大[62—63];季倩雯等(2020)研究发现樟子松树干径向增长量在5月份达到最大[20];在西北干旱区,肖生春等(2012)研究发现6月份是黑河流域下游胡杨树干径向增长量最大的月份[64];在热带湿润地区,吴丽杰等(2020)研究发现苦楝5月份树干径向增长量最大[65]。
在进入缓慢生长期(6月至8月底),空气温度达到全年最高(图2),树木蒸腾作用旺盛,土壤温度也过高,超过了根系吸水的最佳温度,对树干生长造成干旱胁迫[65]。所以此阶段无论是空气温度还是土壤温度都由正相关转为负相关(表2),温度升高抑制树干生长。然而水分方面对树干径向生长的作用与温度截然相反。研究区易受台风影响,虽然进入夏季但湿度和降水量都没有降低(图2),高相对湿度在白天可以抑制树木蒸腾(图2),控制植物体内物质和能量的损失,到晚上有利于树木充分补水[65—66],因此在该时期水分条件,特别是相对湿度对径向生长促进作用更加显著(表2)。至8月底,树干径向大小保持稳定,完成了前期新生细胞的分裂和生长, 进入新生细胞木质化阶段[7, 13, 51],受降水量和空气温度的共同影响[20]。至9月,树木逐渐停止生长。树木停止生长的原因有很多,除受环境因子的影响外更多的是由树木本身复杂的生理特性决定[67],但亚热带地区夏季的持续高温可能会加快木质化过程,是树木提早停止生长时间的重要因素[31, 68]。
4 结论
本次研究采用树干径向生长变化记录仪的监测数据,在多时间尺度上分析亚热带地区的主要树种—马尾松径向昼夜变化特征、季节变化动态以及各个时间尺度下径向变化与环境因子的关系。研究发现:在日尺度上,马尾松日径向变化在全年都呈现白天收缩,夜晚膨胀的模式,在春夏时期收缩和膨胀的程度比秋冬时期更加显著;在季节尺度上,通过分析月平均增长量和日累积量可知,马尾松主要生长月份是3—8月,累计生长量最大的月份是4月,年内生长变化大致可分为4个时期;在马尾松径向变化和环境因子相关性分析中发现,在日尺度上,相对湿度和VPD是控制马尾松径向昼夜变化特征的最主要环境因子;在季节尺度上,土壤温度对马尾松径向生长的影响要比空气温度更为显著,并且在各个时期表现不同,从开始生长期与径向日变化正相关,到快速生长期与径向日变化不相关,最后在缓慢生长期与径向日变化呈现负相关;而水分条件则随时间的推移相关性逐渐增强,降水量在快速生长期成为促进树干生长的主要环境因子,相对湿度在缓慢生长期成为促进树干生长的主要环境因子。因此,在造林设计及管理时应多考虑土壤温度,可采取适当措施调整合适的土壤温度促进马尾松生长,水分条件也相当重要,尤其进入高温时节,应适当增加相对湿度促进生长;同时加强长时间序列的树干径向生长变化的监测研究更加有利于理解树木生长的年际变化及其对气候变化的影响。
猜你喜欢
军事文摘(2021年16期)2021-11-05 08:49:16
绿色科技(2019年5期)2019-11-29 13:17:37
辽宁林业科技(2017年4期)2017-06-22 10:23:36
小猕猴学习画刊(2017年1期)2017-02-17 16:04:14
小猕猴学习画刊(2017年1期)2017-02-17 15:57:35
广东第二课堂·小学(2016年11期)2016-12-06 14:29:33
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:50
广西林业科学(2016年1期)2016-03-20 05:33:00
广西林业科学(2016年1期)2016-03-20 05:32:59
广西林业科学(2016年1期)2016-03-20 05:32:58
