
人工接种东北酸菜发酵过程中氨基酸类物质的代谢变化
2023-09-09 07:19李潇吴兴壮韩艳秋张晓黎高雅王琛
现代食品科技 2023年8期
李潇,吴兴壮,韩艳秋,张晓黎,高雅,王琛
(辽宁省农业科学院食品与加工研究所,辽宁沈阳 110161)
东北酸菜是以白菜或甘蓝为原料制得的盐渍菜,因其独有的风味、口感与营养保健功能,目前已成为我国东北地区最具代表性食品,深受消费者喜爱[1]。而随着现代食品加工技术的不断进步,人工接种乳酸菌发酵酸菜的模式逐渐兴起。这种模式可在一定程度下缩短酸菜的发酵周期,提高酸菜的营养及食用安全性,因此具有广泛的应用前景[2]。
发酵过程中,微生物的协同代谢以及酶和代谢物的进一步反应,能够赋予东北酸菜独有的风味品质[3]。研究表明接种发酵酸菜的风味成分包括酯、醛、酮、醇、酸、氨基酸等[4],更有相关研究表明食品发酵过程中形成的高浓度氨基酸会对发酵产品的风味起关键作用。例如,食醋中含有的较高浓度的谷氨酸和丙氨酸对其鲜味及甜味具有积极影响[5];张建萍等[6]认为腐乳中的咸鲜味主要是由谷氨酸和天冬氨酸所赋予。而Jin等[7]认为中国米酒的甜、鲜、酸和苦味分别与丙氨酸、天冬氨酸和谷氨酸、酪氨酸以及除精氨酸外的其他苦味氨基酸有关;同样,彭杰等[8]则表示呈鲜味的天冬氨酸、天冬酰胺、谷氨酸,呈甜味的丙氨酸与呈苦味的精氨酸一同造就了郫县豆瓣口味的复杂性与厚重感。
近年来,组学技术不断地在发酵酸菜微生物的多样性以及主要风味代谢物的组成方面有所应用[9],然而关于人工接种发酵的东北酸菜发酵过程中风味物质代谢途径变化规律的研究仍有所欠缺。
本研究以人工接种乳酸菌发酵的东北酸菜为研究对象,借助非靶向代谢组学技术分析东北酸菜在人工接种发酵过程中代谢物组成以及氨基酸类物质的代谢变化规律。研究结果可为后续调控东北酸菜风味物质形成,提升东北酸菜品质提供理论依据。
1 材料与方法
1.1 材料与试剂
植物乳杆菌(L.plantarumLNJ005)来源于辽宁省农业科学院食品与加工研究所;干酪乳杆菌(L.caseiBNCC134415)和柠檬明串珠菌(Leu.citreumBNCC194779)均采购自北纳创联生物技术有限公司。东北大白菜(杠菜)、食用盐(中国盐业集团有限公司)和白酒(北京顺鑫农业股份有限公司牛栏山酒厂)均采购自农贸市场。
1.2 仪器与设备
SW-CJ-1FD型单人单面工作净化台,上海沪净医疗器械有限公司;CJ50-3叠加式培养箱,上海程捷仪器设备有限公司;YXQ-LS立式压力蒸汽灭菌器,上海博迅实业有限公司医疗设备厂;TGL-16G-C高速台式冷冻离心机,上海安亭科学仪器厂;LGJ-18C真空冷冻干燥机,北京四环起航科技有限公司;UHPLC-Q Exactive HF-X串联质谱仪,美国Thermo公司。
1.3 实验方法
1.3.1 东北酸菜制备与样品收集
通过生理盐水将植物乳杆菌、干酪乳杆菌和柠檬明串珠菌的菌悬液稀释到浓度为菌体数量级为107。将3种乳酸菌菌悬液按体积比为1:1:1混合均匀,制成液体混合菌发酵剂。
市售大白菜,清洗切丝后,添加2%的食盐(m/m),搅拌均匀。随后将白菜丝装入350 mL塑料泡菜瓶中并压实。将液体混合菌发酵剂加入到泡菜瓶中,20 ℃密封发酵30 d。
根据酸菜的酸度在发酵前期变化较大,酸菜发酵液样品选择在5个采样时间点(第0天、第2天、第10天、第18天和第30天)进行采集,分别在每个泡菜瓶的上、中、下三层采集发酵液后混合均匀(约10 mL)[10],进行代谢物分析。
1.3.2 样品处理与LC-MS/MS分析
取发酵液样品,4 ℃下10 000g离心10 min,分离后取上清在4 ℃下20 000g离心15 min,收集上清。收集的上清液在-80 ℃冷冻后进行真空冷冻干燥,冷冻干燥后的样品用含L-2-氯苯丙氨酸(0.02 mg/mL)作为内标的提取液:甲醇:水=4:1(V/V)溶解。研磨6 min后,低温超声提取30 min。在4 ℃下13 000g离心15 min,收集上清进行UHPLC-Q Exactive HF/MS分析。将每个时间点采集的样品作为一组,每组样品测定5个平行。并将发酵不同时间的样品组作为差异分析对比组。每个处理样品中取20 μL上清液混合作为质量控制样品(QC)。
色谱条件:采用ACQUITY UPLC HSS T3色谱柱(100 mm×2.1 mm,1.8 µm),流动相A为包含0.1%(V/V)甲酸的三聚氰酸水溶液H2O/ACN(95:5);流动相B为包含0.1%(V/V)甲酸的三聚氰酸-乙腈溶液ACN/IPA/H2O(47.5:47.5:5)。进样量为2 µL,柱温为40 ℃。梯度洗脱条件为0 min,100:0(V/V),流速0.4 mL/min;3.5 min,75.5:24.5(V/V),流速0.4 mL/min;5 min,35:65(V/V),流速0.4 mL/min;5.5 min,0:100(V/V),流速0.4 mL/min;7.4 min,0:100(V/V),流速0.6 mL/min;7.6 min,48.5:51.5(V/V),流量0.6 mL/min;7.8 min,0:100(V/V),流速0.6 mL/min;9 min,0:100(V/V),流速0.4 mL/min;10 min,0:100(V/V),流速0.4 mL/min。
质谱条件:采用正负电离扫描模式,扫描范围为70~1 050m/z,加热器温度为425 ℃;毛细管温度为325 ℃;鞘气流速50 arb;辅助气流速13 arb;离子喷雾电压,正电离模式为3.5 kV,负电离模式为-3.5 kV。整个分析过程中,QC样品每隔5~15个进样一次,用于监控稳定性。
1.3.3 数据分析
将原始数据导入Progenesis QI(Waters Corporation,Milford,USA),得到滞留时间、质荷比、峰强度等参数信息。利用串联质谱(MS/MS)将这些特征值与MS片段在网上公共数据库和自建数据库(http://www.hmdb.ca/,https://metlin.scripps.edu/)上进行比对。MS质量误差小于10×10-6,是基于二级质谱匹配分数确定代谢物种类。数据分析在Majorbio Cloud Platform在线平台(www.majorbio.com)上进行。
采用Relative-betweeness Centrality方法进行拓扑学分析,采用Benjamini-Hochberg方法(BH法)进行多重检验校正,对P值进行校正,当经过校正的P<0.05时,认为此通路存在显著富集情况。
2 结果与分析
2.1 接种发酵酸菜代谢物及代谢功能通路分析
人工接种发酵东北酸菜(以下简称酸菜)不同阶段代谢物的共有数目见图1。在酸菜接种发酵过程中共检测出638种代谢物。从发酵开始到结束共有619种代谢物同时存在,且众多代谢物含量在发酵过程中均存在不同程度的改变。此外,发酵前18 d共有的4种化合物中甘油磷脂类化合物存在差异下调,而发酵第2天到第30天共有的13种化合物中脂类、酚类和氨基酸类化合物存在差异上调。在已被确定的代谢物中,以氨基酸,多肽及其类似物为主,占总代谢物的25.43%,随后是脂肪酸及其共轭物和碳水化合物,占总代谢物的5.31%和4.36%。而萜类、嘌呤类、羰基化合物、以及其他类化合物的种类较少。

图1 接种发酵东北酸菜代谢物数目Fig.1 Number of metabolites of Inoculated fermented Dongbeisuancai
通过将代谢途径涉及的基因与京都基因和基因组百科全书(KEGG)数据库进行匹配,可以更加直观地了解酸菜发酵过程中的生物转化途径,详细见图2a。接种发酵酸菜代谢途径主要与氨基酸代谢、脂类代谢、其他次生代谢物生物合成、碳水化合物代谢、核苷酸代谢、外源生物降解与代谢、辅酶因子和维生素代谢、萜类和多酮类代谢、能量代谢以及聚糖生物合成与代谢有关。

图2 接种发酵东北酸菜代谢途径分类与化合物分类Fig.2 Metabolic pathways and compounds classification of inoculated fermented Dongbeisuancai
不同的代谢途径对酸菜香气、滋味等品质的形成具有重要意义。脂类代谢,包括“不饱和脂肪酸的生物合成”、“脂肪酸代谢及生物合成”、“丙酮酸代谢”等均涉及挥发性风味酯的形成。氨基酸代谢、核苷酸代谢、萜类和多酮类代谢也同样涉及挥发性风味物质的形成。而碳水化合物代谢,包括“淀粉和蔗糖代谢”以及“果糖和甘露糖代谢”,也可能是酸菜发酵过程中产生的香气和滋味化合物的重要来源。此外,葡萄糖代谢是对酸菜酸味具有重要贡献的乳酸和醋酸的生产途径。虽然碳水化合物代谢是酸菜发酵中的基础代谢过程,它与其他代谢途径的协同运作能够反映微生物利用碳水化合物、蛋白质和其他基质生长的生物学必要性[11]。但氨基酸代谢同样是接种发酵酸菜的主要代谢途径,并且对酸菜风味与滋味的形成具有重要意义[12]。在与生物合成代谢相关的化合物中,氨基酸的种类最多,这也说明氨基酸代谢在酸菜发酵过程中的重要作用(图2b)。
2.2 接种发酵酸菜中氨基酸分析
通过UHPLC-Q Exactive系统的检测以及与代谢物数据库的匹配,得到酸菜接种发酵过程中氨基酸的种类与变化,详细见图3。在混菌接种发酵的酸菜中鉴定出12种氨基酸,分别是L-精氨酸、L-脯氨酸、L-甲硫氨酸、焦谷氨酸、反-4-L-羟基谷氨酸、L-半胱氨酸、L-丝氨酸、L-天冬酰胺、β-丙氨酸、L-天冬氨酸、L-谷氨酰胺、L-谷氨酸,并且在发酵过程中,L-精氨酸和L-脯氨酸的丰度始终较高。但相比之下,自然发酵的酸菜中含有更多的游离氨基酸种类,这可能是由于混菌的接种使发酵过程具有主导乳酸菌,而自然发酵酸菜中微生物的种类更加丰富,也因此产生更多的代谢物质[13]。氨基酸的呈味效果与其手型结构有相关性,L-氨基酸可呈现出酸、甜、苦、咸、鲜5种味道,但L-氨基酸呈甜味阈值均较高,因此其呈甜味不明显[14]。接种发酵前两天,除L-丝氨酸外,其他氨基酸的丰度均呈现上升趋势。尤其是L-半胱氨酸,上升程度明显较高。与之相反,L-丝氨酸的丰度呈现急剧下降趋势。这可能是由于在发酵前两天存在L-丝氨酸与L-半胱氨酸的转化。发酵第2天开始L-谷氨酸与反-4-L-羟基谷氨酸的丰度基本维持稳定,而其他氨基酸的丰度缓慢上升到发酵第10天基本稳定。这说明前10天的代谢过程涉及大部分氨基酸的合成与转化,也表明前10天是接种发酵酸菜风味与滋味形成的重要时期。值得注意的是,L-半胱氨酸的丰度在发酵前10天始终显著上升,这表明与L-半胱氨酸生物合成的相关代谢较为活跃。此外,呈酸味的半胱氨酸大量积累,说明发酵前10天是酸味形成的重要时期。同时与Xiao等[15]的研究结果相似,东北酸菜的谷氨酸与天冬氨酸具有较高的丰度,结合两者较低的酸味与鲜味阈值,导致酸菜整体酸感较强。
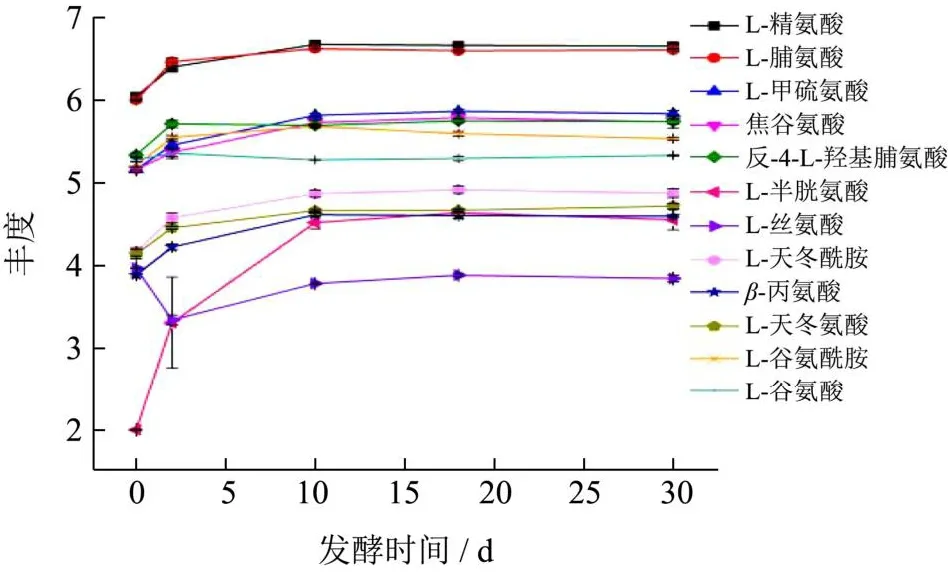
图3 接种发酵东北酸菜发酵过程氨基酸变化Fig.3 Changes of amino acid during fermentation of Dongbeisuancai
2.3 接种发酵酸菜中氨基酸代谢通路富集分析
对酸菜发酵过程中的代谢物进行富集分析,使用超几何分布算法获得代谢物显著富集的通路详见图4所示。

图4 接种发酵东北酸菜氨基酸代谢通路富集Fig.4 Amino acid metabolism enrichment pathway of inoculated fermented Dongbeisuancai
在人工接种发酵前两天,多种代谢物在相关氨基酸代谢通路中显著富集并显示出重要性。而对应的多组氨基酸代谢通路同时显示出重要作用。显著富集的代谢物对应的重要氨基酸代谢包括精氨酸和脯氨酸代谢、半胱氨酸和甲硫氨酸代谢、甘氨酸、丝氨酸和苏氨酸代谢、谷胱甘肽代谢以及赖氨酸降解。在这些氨基酸相关的代谢途径中,精氨酸和脯氨酸代谢富集更多的代谢物,同时在代谢中具有更重要的作用。其次是半胱氨酸和甲硫氨酸代谢,虽然该代谢通路富集的代谢物较少,但其在代谢中仍具有较高的相对影响值。谷胱甘肽代谢通路富集了较多的代谢物,而其影响值介于甘氨酸、丝氨酸和苏氨酸代谢与赖氨酸降解之间。发酵第2天到第10天,具有重要影响的氨基酸相关代谢路径显著减少。在这一发酵阶段,谷胱甘肽代谢表现出较高的重要性。而半胱氨酸和甲硫氨酸代谢以及赖氨酸降解虽然仍具有重要性,但两者的代谢物富集相对较少。发酵第10天到第18天,组氨酸代谢路径上出现了代谢物富集,并且其与甘氨酸、丝氨酸和苏氨酸代谢均具有重要作用,而半胱氨酸和甲硫氨酸代谢的重要性降低。值得注意的是,甘氨酸、丝氨酸和苏氨酸代谢路径在发酵第2天到发酵第10天无代谢物富集,这也表明甘氨酸、丝氨酸和苏氨酸代谢的路径在发酵前两天和第10天到第18天不同。纵观整体发酵期间,精氨酸和脯氨酸代谢路径仅在发酵前两天具有重要性,因此发酵前两天以精氨酸和脯氨酸代谢为主。而谷胱甘肽代谢在发酵第2天到第10天,重要性显著提高,随后通路代谢物无富集。发酵第10天到第18天,代谢路径以甘氨酸、丝氨酸和苏氨酸代谢以及组氨酸代谢为主。此外,半胱氨酸和甲硫氨酸代谢在发酵前18天,赖氨酸降解在发酵前10天始终具有重要性,但伴随发酵进行,重要性逐渐降低。随后,发酵第18天到第30天,已无具有明显重要性的氨基酸代谢路径出现。
2.4 接种发酵酸菜中氨基酸代谢途径分析
根据在酸菜发酵过程中富集的氨基酸,采用KEGG对人工接种酸菜发酵过程中涉及的氨基酸代谢途径进行了分析。参与不同氨基酸代谢途径的代谢物和关键酶详见图5和表1。针对发酵前两天具有重要作用的精氨酸和脯氨酸代谢途径,根据富集的代谢物L-精氨酸、N2-琥珀酰-L-鸟氨酸、L-谷氨酸、5-半醛-L-谷氨酸、L-脯氨酸、γ-谷酰-γ-氨基-丁酸、4-氨基丁酸和亚精胺,预测该代谢途径主要包括:(1)琥珀酰基转移酶途径,涉及5种酶。通过琥珀酰基相关的转移酶、水解酶和氧化还原酶将L-精氨酸转化成L-谷氨酸。(2)脯氨酸生物合成途径,涉及3种酶。通过激酶、半醛脱氢酶和羧酸还原酶将L-谷氨酸转化成L-脯氨酸。(3)γ-氨基丁酸(GABA)生物合成途径,涉及6种酶。Yang等[16]认为GABA是由谷氨酸经谷氨酸脱羧酶的作用转化而来。而本研究由于检测到γ-谷酰-γ-氨基丁酸,因此预测GABA合成途径是由L-精氨酸开始,通过不同的裂解酶、水解酶、氧化还原酶的作用,最终转化成4-氨基丁酸。(4)多胺生物合成途径,涉及1种酶。通过亚精胺合成酶将丁二胺转化成亚精胺。详细转化途径见图5精氨酸和脯氨酸代谢。
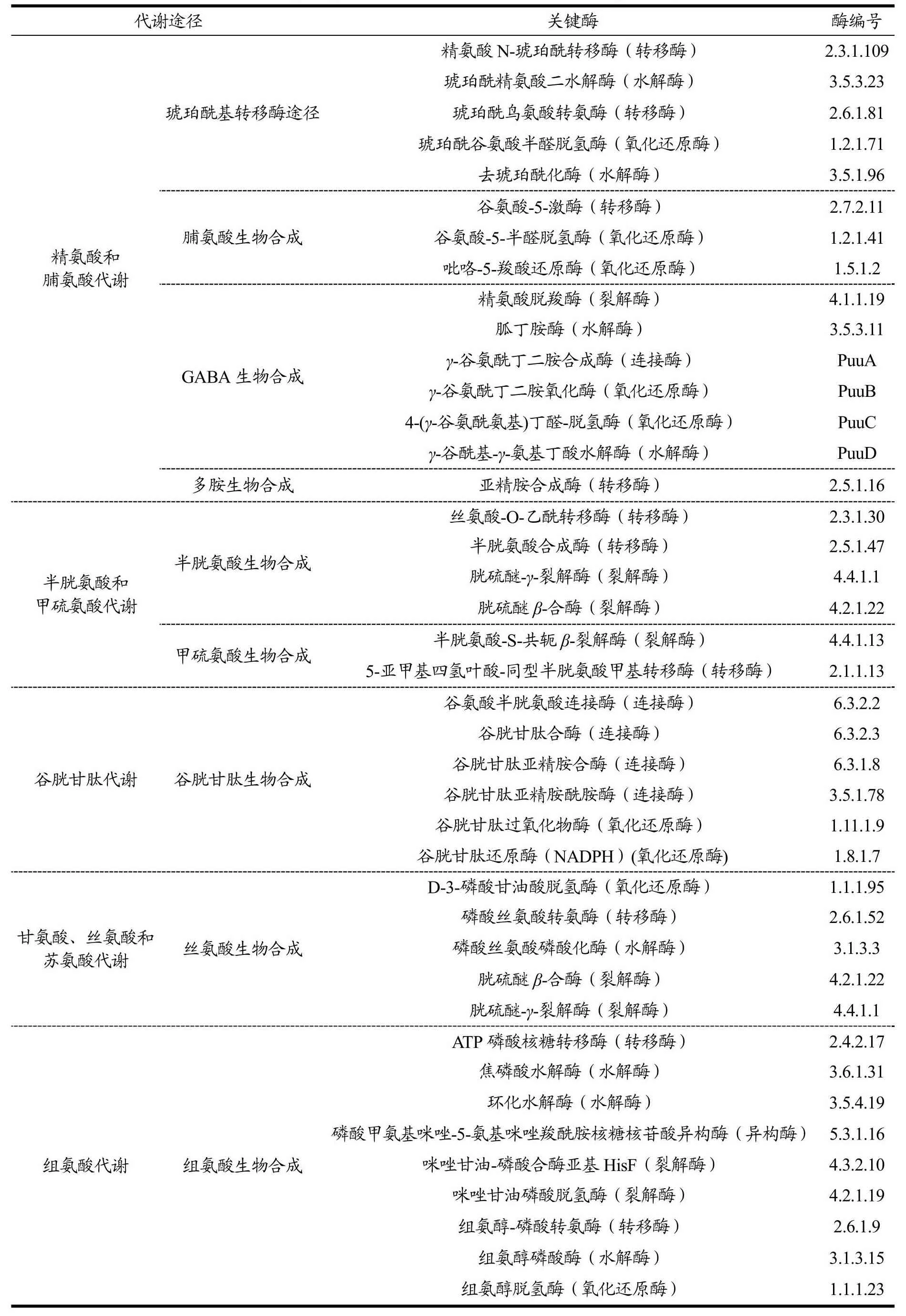
表1 不同氨基酸代谢途径的关键酶Table 1 Key enzymes of different amino acid metabolic pathways

图5 接种发酵酸菜中氨基酸代谢途径分析Fig.5 Analysis of amino acid metabolism pathway in inoculated fermented Dongbeisuancai
针对半胱氨酸和甲硫氨酸代谢途径,预测该代谢途径主要包括:(1)半胱氨酸生物合成途径,涉及4种酶。通过2种转移酶将L-丝氨酸转化成L-半胱氨酸以及在2种裂解酶的作用下将同型半胱氨酸化成L-半胱氨酸。其中L-丝氨酸转化成L-半胱氨酸也可以通过这两种氨基酸的含量变化得到印证(图3)。(2)甲硫氨酸生物合成途径,涉及2种酶。通过裂解酶和转移酶将胱硫醚转化成L-甲硫氨酸。此外,半胱氨酸也可以通过转胺反应转化为甲硫氨酸[17]。详细转化途径见图5甲硫氨酸和半胱氨酸代谢。
谷胱甘肽是由谷氨酸、半胱氨酸和甘氨酸作为前体组成的含γ-酰胺键和巯基的三肽物质[18]。针对谷胱甘肽代谢途径,富集的代谢物包括L-谷氨酸、L半胱氨酸、L-γ-谷氨酸半胱氨酸、还原型谷胱甘肽(GSH)、氧化型谷胱甘肽(GSSG)和亚精胺。预测该代谢途径涉及谷胱甘肽生物合成途径,涉及6种酶。与张子洁等的研究结果相似[19],通过连接酶将L-谷氨酸和L-半胱氨酸转化成GSH。或者亚精胺经连接酶转化成GSH。同时还存在GSH和GSSG的相互转化。详细转化途径见图5谷胱甘肽代谢。
丝氨酸代谢以丝氨酸生物合成为主,代谢途径涉及5种酶,由糖酵解途径(EMP)中合成的3磷酸甘油酸(3PG)经多种酶转化成L-丝氨酸,同时L-半胱氨酸也可经由胱硫醚途径转化成L-丝氨酸。详细转化途径见图5丝氨酸代谢。
组氨酸代谢以组氨酸生物合成途径为主。该途径是由磷酸戊糖途径生成的5-磷酸核糖-1-焦磷酸(PRPP)经多种酶逐步转化成L-组氨醛后经脱氢酶转化成L-组氨酸。详细转化途径见图5组氨酸代谢。
赖氨酸降解途径富集的代谢物包括N6-乙酰-L-赖氨酸、L-β-赖氨酸、3,5-二氨基己酸、5-羟基赖氨酸和S-2-羟戊二酸。因此预测赖氨酸降解途径主要由L-赖氨酸(1)在乙酰转移酶作用下转化成N6-乙酰-L-赖氨酸,(2)在2种异构酶的作用下先转化成L-β-赖氨酸后转化成3,5-二氨基己酸,(3)在多种酶的作用下转化成5-羟基赖氨酸,(4)在多种酶作用下转化成S-2-羟戊二酸后又经氧化还原酶转化成琥珀酸,参与三羧酸循环(TCA循环)。详细转化途径见图5赖氨酸降解。
3 结论
本研究鉴定了人工接种发酵东北酸菜发酵过程中的代谢产物,其中以氨基酸、多肽及其类似物占比较大。同时,鉴定出的氨基酸对酸菜酸味的形成具有重要作用,并且发酵前10天是接种酸菜风味与滋味形成的重要时期。酸菜发酵过程中主要代谢途径涉及氨基酸的生物合成和代谢,根据代谢物的富集发现发酵前两天以精氨酸和脯氨酸代谢为主,第2天到第10天以谷胱甘肽代谢为主,第10天到第18天以丝氨酸和组氨酸代谢为主,此外赖氨酸降解,半胱氨酸和甲硫氨酸代谢分别在发酵前10天和前18天始终存在,但伴随发酵进行,逐渐减弱。发酵18天后,基本无氨基酸代谢路径出现。本研究揭示了氨基酸代谢调控接种发酵东北酸菜风味的机制,但还需要进一步基于微生物在基因和酶水平上的表达研究来完善机制研究。本研究从氨基酸代谢途径的角度切入,完善了东北酸菜发酵机理,同时为提高东北酸菜风味和口感提供了理论基础。
猜你喜欢
沈阳医学院学报(2022年6期)2022-12-03
南昌大学学报(医学版)(2021年1期)2021-11-29
动物营养学报(2021年6期)2021-07-12
广东饲料(2016年3期)2016-12-01
中外医疗(2015年11期)2016-01-04
动物营养学报(2015年10期)2015-12-01
中国当代医药(2015年30期)2015-03-01
应用化工(2014年10期)2014-08-16
中国中医药现代远程教育(2012年18期)2012-09-18
河南医学研究(2012年3期)2012-08-15
