
黄土高原旱塬区3种堇菜属植物光合特性分析
2023-09-03 08:12:32江龙龙黄文迪吕小旭田树鑫王冲
山西农业科学 2023年4期
江龙龙 黄文迪 吕小旭 田树鑫 王冲

关键词:堇菜属;紫花地丁;光合特性;适应性;黄土高原旱塬区
堇菜属(Viola)隶属于堇菜科(Violaceae),该属植物为1年生或多年生草本植物,在我国约有111余种[1],分布广泛、资源丰富,普遍具有清热解毒、凉血消痈的功效,迄今明确功效与主治记载的有20余种[2],该属植物也常作为早春园林绿化植物开发利用[3]。该属植物多生于林缘、山坡、草地、荒地、田边、村庄路边等处,生境不同其主要分布的种类也有差异。黄土高原旱塬区常分布紫花地丁(Violaphilippica)、早开堇菜(Violaprionantha)、斑叶堇菜(Violavariegata)等该属植物,同一生境下不同堇菜属植物的生长不同,不同生境下同种堇菜种群大小也有差异[4]。黄土高原是较易受到干旱影响的地区,该地区的植物对于干旱具有较高的敏感脆弱性[5],同时,随着退耕还林和农田撂荒后群落结构发生改变,堇菜属植物作为地被植物易受光照的改变而影响种群繁殖和生态适应[6],因而,推测水分和光照是限制黄土高原分布的堇菜属植物生态适应性的关键因子。
光合作用是植物重要的生理生化过程,是植物生长发育和产量形成的基础[7]。植物的光合特性在一定程度上能够体现其对生境的响应情况,光合特征参数的变化在一定程度上能反映植物对环境的生长适应性,通过对植物净光合速率、蒸腾速率、气孔导度、光能利用效率及水分利用效率等光合参数的研究,可以了解光合作用中植物利用光强的能力及其对生境的适应情况[8],反映植物在环境中必备的生存条件,掌握植物生态适应性[9]。王匡[10]通过比较4种铁线莲植物光合特性,推测甘青铁线莲能够适应较为干旱的环境;刘旻霞等[11]通过植物光合特性研究3种绿化树种光合生理机制,分析生理生态因子相关性,为兰州北山人工造林树种优选及生态环境恢复提供参考依据;和红晓等[12]研究表明,红花巨紫荆和粉白花巨紫荆的光合特性存在差异,红花巨紫荆的光合速率及水分利用率均较高,说明其在有机物合成、抗逆性等方面表现较突出,在引种栽培过程中可以合理开发和应用。目前,堇菜属植物光合特性的研究主要集中在宝山堇菜[13]、紫花地丁[13]、维西堇菜[14]、斑叶堇菜[15]等物种的单独研究,未见通过比较堇菜属植物光合特性的差异来研究堇菜属植物光合特征如何响应生境差异。
为了解堇菜属植物分布及种群特征与生境的关系,本试验通过对黄土高原旱塬区同一生境下不同堇菜属植物的光合特性参数进行对比分析,分析不同生境下紫花地丁光合特性差异、光合特性与生态因子相关性,掌握黄土高原旱塬区堇菜属植物生态适应性的特点,预测生态环境变化对堇菜属植物分布的影响,为干旱区堇菜属植物资源开发利用提供基础资料。
1 材料和方法
1.1 研究区概况
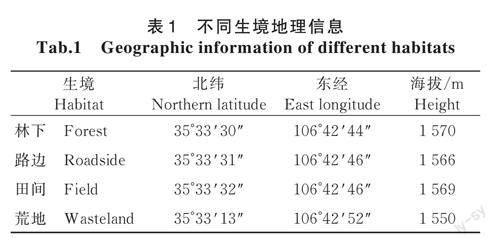
研究区域位于甘肃省平涼市北山山顶自然生境,属黄土高原旱塬区,为干旱、半湿润季风型大陆性气候,四季分明,土壤类型主要为壤土;年平均气温8.9℃,春旱和伏旱十分严重,水资源不足,降雨分布不均,主要集中在7—9月,年平均降水量503.7mm,年无霜期133~286d,年均日照时数2424.3h。4种生境地理信息如表1所示。
1.2 试验材料
选取相对均匀分布于同一生境路边下的紫花地丁、斑叶堇菜、早开堇菜为试验材料,选取路边、田间、林下、荒地4种生境下的紫花地丁为试验材料。要求每个待测植株生长发育良好、无病虫害、长势基本一致。
1.3 试验方法
随机选取试验点的待测植株完整第3枚叶,且大小相似的叶片各3个,采用北京雅欣理仪科技有限公司生产的YaXin-1241型叶面积仪对叶片的叶面积进行测量(cm2)。
使用北京雅欣理仪科技有限公司生产的Yaxin-1005便携式光合荧光仪,于2021年4月(花期)、5月(果期)、6月(叶期),分别选取晴朗天气,在不同生境下测定每株堇菜第3或第4枚成熟叶片的光合特性参数。测定时间为8:00—18:00,每间隔2h测定一次,每片叶片重复3次。测定前将气孔流速设定为0.5L/min,叶面积设定以叶面积仪所测为准,同时测定光合有效辐射(PAR)、叶片温度(Tleaf)、净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)。
光能利用效率(LUE)=Pn/PAR(1)
水分利用效率(WUE)=Pn/Tr(2)
在不同生境分别采取5个土样(土壤深度10cm),置于铝盒中,利用烘干法测定土壤含水率(SMC)。
1.4 数据分析
试验利用SPSS22.0和Origin2020软件对数据进行统计分析。
2 结果与分析
2.1 3种堇菜属植物光合特性比较
2.1.1 3种堇菜属植物光合速率变化 3种堇菜属植物Pn日变化显示(图1),斑叶堇菜与紫花地丁均呈双峰曲线变化,分别出现在10:00(主高峰)和16:00(次高峰),斑叶堇菜与紫花地丁主高峰值分别为20.64、25.37μmol/(m2·s),均在14:00出现明显的“光合午休”。早开堇菜在12:00达到最高峰值(16.86μmol/(m2·s)),无明显“光合午休”。综合Pn日变化情况,紫花地丁8:00—16:00,其净光合速率均显著高于斑叶堇菜和早开堇菜(P<0.05)。
3种堇菜属植物不同生长期Pn日均值变化显示(图1),斑叶堇菜、紫花地丁及早开堇菜随着生长周期延续,从花期至果期至叶期均呈下降趋势。纵观整个生长周期的净光合速率大小依次为紫花地丁>斑叶堇菜>早开堇菜。
综合3种堇菜属植物Pn日变化及各生长期日均变化情况,紫花地丁在整个生长过程中的净光合速率均高于斑叶堇菜及早开堇菜。
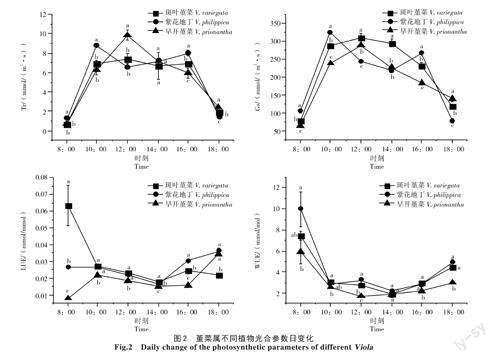
2.1.2 3种堇菜属植物光合参数变化 3种堇菜属植物Tr比较显示(图2),斑叶堇菜与紫花地丁均呈双峰曲线变化。斑叶堇菜在12:00达到主高峰值(7.36mmol/(m2·s)),紫花地丁在10:00达到主高峰值(8.78mmol/(m2·s)),早开堇菜在12:00达到最高峰值(9.84mmol/(m2·s))。Tr日均值大小依次为紫花地丁>早开堇菜>斑叶堇菜,其中,紫花地丁Tr与Pn变化趋势一致。
3种堇菜属植物Gs比较显示(图2),紫花地丁呈双峰曲线变化。紫花地丁在10:00达到主高峰值(324.6mmol/(m2·s)),斑叶堇菜在12:00达到最高峰值(310.7mmol/(m2·s)),早开堇菜在12:00达到最高峰值(288.93mmol/(m2·s))。Gs日均值大小依次为斑叶堇菜>紫花地丁>早开堇菜,其中,紫花地丁Gs与Pn变化趋势一致。
种堇菜属植物LUE比较显示(图2),斑叶堇菜在8:00达到最高峰值(0.064mmol/mmol),紫花地丁与早开堇菜均在18:00达到最高峰值,分别为0.036、0.034mmol/mmol。3种堇菜LUE均从10:00开始降低,至14:00开始升高。LUE日均值大小依次为斑叶堇菜>紫花地丁>早开堇菜。
3种堇菜属植物WUE比较显示(图2),斑叶堇菜、紫花地丁及早开堇菜均在8:00达到最高峰值,在12:00及14:00达到最低值。斑叶堇菜、紫花地丁及早开堇菜的WUE最高峰值分别为7.46、9.98、5.92mmol/mol。WUE日均值大小依次为紫花地丁>斑叶堇菜>早开堇菜。
综合3种堇菜属植物光合参数变化情况,紫花地丁Tr及WUE均为最高,在10:00—18:00太阳有效辐射最强的时间段其LUE也处于较高水平,同时紫花地丁光合参数日变化趋势表明,在中午强光照和高温环境条件下,存在明显的“光合午休”现象;斑叶堇菜Gs及LUE均为最高,存在“光合午休”现象;早开堇菜各项光合参数均最低,无“光合午休”现象。
2.2 紫花地丁不同生境下光合特性比较
2.2.1 不同生境下光合速率变化 4种生境下紫花地丁Pn日变化显示(图3),田间、路边生境呈双峰曲线变化,分别出现在10:00(主高峰)和16:00(次高峰),田间与路边主高峰值分别为24.42、25.37μmol/(m2·s),分别在12:00和14:00出现“光合午休”。林下生境在12:00达到最高峰值(17.77μmol/(m2·s)),荒地生境在14:00达到最高峰值(35.66μmol/(m2·s)),均无“光合午休”。综合Pn日变化情况,田间及路边生境下紫花地丁净光合速率较林下和荒地高,但荒地生境在光照强度最强的14:00达到最高峰值。
4种生境下,紫花地丁不同生长期Pn日均值变化显示(图3),其净光合速率大小依次为田间>路边>荒地>林下。
综合紫花地丁在4种生境条件下的Pn日变化及各生长期日均变化情况,田间的紫花地丁整个生长过程中净光合速率均高于其他生境条件。
2.2.2 紫花地丁光合参数与生态因子相关性分析
综合4种生境条件下紫花地丁生理生态因子的相关性,生态因子光合有效辐射(PAR)、叶面温度(Tleaf)及土壤含水量(SMC)间呈极显著正相关(P<0.01),光合因子中Pn与Tr呈显著正相关(P<0.05);光合参数Pn、Tr与生态因子PAR、Tleaf、SMC呈正相关,光合参数Gs、LUE、WUE与生态因子PAR、Tleaf、SMC呈负相关;光合参数Pn、Tr与Gs呈正相关,与LUE、WUE呈负相关(表2)。
3 结论与讨论
光合作用是植物同化环境碳的重要生理过程,相关参数表征着植物对生境因子的适应能力[16],如Pn是表示光合作用強弱变化的核心指标之一,Pnmax反映了植物的遗传特性和对环境的适应能力,Tr大小直接决定植物在单位时间通过蒸腾作用散失的水量多少,气孔是植物叶片与外界进行水气更新的“通道口”,资源利用效率是反映植物利用水分、光能等资源完成生长发育的重要生理指标[11]。
本试验通过对分布于黄土高原旱塬区路边3种常见堇菜属药用植物光合特性研究显示,紫花地丁Pn日变化指数及最高峰值均显著高于斑叶堇菜和早开堇菜,且紫花地丁在花期、果期、叶期整个生长期的Pn日均指数也高于其他堇菜,表明紫花地丁相较于其他堇菜有更大的光合作用潜力和能力;紫花地丁Tr及WUE显著高于其他堇菜,表明其在生理活动过程中消耗水形成有机质的基本效率也最高;紫花地丁在10:00—18:00太阳有效辐射最强的时间段,其LUE处于较高水平,表明紫花地丁通过光合作用将能量转化为有机干物质的效率较高;紫花地丁Pn、Tr、Gs及LUE日变化趋势高度一致,表明其在应对路边无遮挡干旱的强光照和高温环境条件方面,采取了“光合午休”的生理对策,较其他堇菜表现出更强适应能力及抗逆性,也是紫花地丁在黄土高原旱塬区能够分布较广的原因之一。
植物叶片光合参数的日变化可以反映光合作用与外界环境变化的关系,Pn的大小不仅受到植物生物学特征影响,还受多种生态因子的影响[17]。本研究通过对比不同生境条件下紫花地丁在整个生长周期中Pn变化情况,依次为田间>路边>荒地>林下,表明紫花地丁在田间环境下表现出较高的光合作用能力;4种生境条件下紫花地丁生理生态因子相关性分析表明,生态因子PAR、Tleaf及SMC间呈极显著正相关,光合因子中Pn与Tr呈显著正相关,生态因子PAR、Tleaf、SMC与光合参数Pn、Tr呈正相关,与Gs、LUE、WUE呈负相关。分析认为,作为植物光合作用动力的PAR影响紫花地丁叶片温度、促进水分蒸发从而加速光合效率,随着PAR进一步增强,造成紫花地丁气孔关闭、降低对资源利用效率从而出现“光合午休”,这与紫花地丁在光照较强的环境中可提高其体内能量代谢水平[18]的结论一致,即适当强度的PAR是紫花地丁生物质积累和能量代谢等生理活动的基础;生态因子SMC变化与紫花地丁LUE、WUE呈负相关,表明在土壤含水率降低即出现干旱情况下,紫花地丁通过提高对光能及水分资源的利用效率来适应环境,这与韩阳瑞等[19]研究干旱区沙柳的水分利用效率高于柠条和油蒿,从而证明沙柳对干旱环境的适应能力优于其他2种灌木的结论吻合,即水分利用效率越高,植物对干旱环境的适应性也越强。
此外,紫花地丁在荒地生境中14:00的净光合速率达到最高峰值(35.66μmol/(m2·s)),显著高于4种生境Pn日变化任一时刻,对比发现荒地生境此刻PAR参数(752.33μmol/(m2·s))与田间生境PAR日均值(769.44μmol/(m2·s))接近,分析认为荒地生境下高大的杂草及亚灌木植物适度遮挡中午的强光,从而打破紫花地丁“光合午休”,提高了该时段的净光合速率,表明紫花地丁在适度遮阴条件下能够提高其光合作用效率。
光合参数的动态变化体现了植物对环境因子的适应机制,是反映植物对光能利用能力和效率的重要指标[20]。黄土高原旱塬区堇菜属3种植物都有应对干旱和强光照的光合特性,3种堇菜属植物光合特性符合其在黄土高原旱塬区的分布特点,路边干旱无遮挡区域下紫花地丁比早开堇菜和斑叶堇菜更适宜,但其最适宜的生境为田间,因而,紫花地丁在黄土高原旱塬区分布最广,各种生境均有分布,紫花地丁能更加适应黄土高原旱塬区的生态环境;而早开堇菜多分布在荒地、斑叶堇菜多分布在林下等适度遮阴的生境中。堇菜属植物光合特性表明,其通过提高光合作用中水分利用效率(WUE)和关闭气孔降低蒸腾速率(Tr)来应对干旱条件,印证了前人通过研究其他方面的特性来解释堇菜属植物具有较好的抗旱能力[3,21-23];光照强度可能是限制黄土高原旱塬区堇菜属植物分布的主要因素。紫花地丁在中午光照强度最高时的适度遮阴可打破植物“光合午休”,表现出更大的光合作用潜力和能力,在有机质形成和干物质积累方面更具优势,且有更强适应能力及抗逆性,提升植株对资源的有效利用,表现出了优于早开堇菜和斑叶堇菜的生态适应性。
猜你喜欢
故事作文·低年级(2024年2期)2024-03-06 10:52:07
保健与生活(2023年15期)2023-08-04 11:02:30
农业科技通讯(2023年1期)2023-02-12 07:07:54
中国外汇(2019年23期)2019-05-25 07:06:20
小学生导刊(2018年20期)2018-11-30 03:05:12
山东农业科学(2016年12期)2017-01-21 14:41:54
山东农业科学(2016年12期)2017-01-21 14:39:43
山东农业科学(2016年11期)2016-12-17 21:16:16
吉林农业·下半月(2016年10期)2016-12-16 23:45:53
金色少年(奇趣科普)(2016年5期)2016-12-01 03:24:36
