
棉花窄卷苞叶基因fg 的精细定位
2023-08-23 07:42:32王康文王雪平王军梁雨裴小雨任翔王星星张先亮彭云玲臧新山马雄风
棉花学报 2023年3期
王康文,王雪平,王军,梁雨,裴小雨,任翔,3,王星星,张先亮,3*,彭云玲,臧新山,3,4*,马雄风,3,4*
(1.甘肃农业大学农学院/ 甘肃省干旱生境作物学重点实验室/ 甘肃省作物遗传改良与种质创新重点实验室,兰州730070;2.中国农业科学院棉花研究所/ 棉花生物育种与综合利用全国重点实验室/ 农业农村部棉花生物学与遗传育种重点实验室,河南 安阳 455000;3.中国农业科学院西部农业研究中心,新疆 昌吉 831100;4.郑州大学农学院,郑州 450000)
棉花是重要的天然纤维作物,也是重要的油料作物,在我国国民经济中处于战略地位。新疆是我国主要的商品棉生产基地。目前,新疆北疆地区已基本实现棉花机械化采摘,南疆地区机采棉面积占比逐年增加。机采棉技术的推广运用对全国棉花产业发展具有重要的推动作用[1]。棉花在机械收获时混入的杂质较多一直是困扰纺织工业的主要难题。目前,我国的机采棉含杂率普遍较高,一般在12%~16%,最高达到20%[2],导致后续籽棉加工清理工序增多、纤维品质显著下降。
棉花一般有3 片苞叶,呈心脏形,紧贴蕾铃,苞叶与蕾铃发育、光合产物积累、病虫害防治和纺纱品质密切相关[3]。已发现的棉花苞叶变异类型主要包括窄卷苞叶[4-5]、凋萎苞片[6]和狭薄卷苞叶[7]。1940 年,美国阿肯色州棉农Frego 首次发现棉花窄卷苞叶突变株,窄卷苞叶狭长、扭曲翻转生长,与絮铃贴合不紧密;窄卷苞叶突变体在吐絮期苞叶自然下卷,至采收期干枯苞叶与籽棉分离,且处在铃壳之下,因此可以降低机采棉含杂率[7]。
1955 年,Green[5]证明棉花窄卷苞叶的遗传受1 对隐性基因控制。2010 年,Li 等[4]利用陆地棉(Gossypium hirsutum)T582(含fg基因)和海岛棉(G.barbadense)Hai7124 的F2群体,将窄卷苞叶基因fg定位在A03 染色体长臂上简单序列重复(simple sequence repeat,SSR)标记NAU3016 和NAU3172 之间,fg与NAU3016 和NAU3172 的遗传距离分别为0.3 cM (centiMorgan,厘摩)和4.7 cM,对应的物理区间为1.87~3.85 Mb(million base pair,百万碱基对),物理距离为1.98 Mb。2017 年,Zhu 等[8]利用混合分组分析测序法(bulked segregate analysis-sequencing, BSA-Seq)将fg基因定位在A03 染色体上物理位置0.00~4.50 Mb 区间内。
本研究以陆地棉T582 为母本,分别与父本陆地棉TM-1、海岛棉3-79 杂交构建2 个F2分离群体,开发插入缺失(insertion-deletion, Indel)分子标记对窄卷苞叶基因fg进行精细定位,为该基因的图位克隆和育种利用奠定基础。
1 材料与方法
1.1 试验材料
配制杂交组合所用的亲本材料为陆地棉T582、陆地棉遗传标准系TM-1 和海岛棉品种3-79,均由中国农业科学院棉花研究所国家棉花种质中期库提供,本课题进行多年连续自交保存。
亲本苞叶性状稳定,3-79 和TM-1 为正常苞叶,T582 为窄卷苞叶。以陆地棉T582 为母本,分别与父本陆地棉TM-1、海岛棉3-79 杂交构建2个F2分离群体。群体1(T582×TM-1)F2群体包含370 个单株;群体2(T582×3-79)F2群体包含2 667 个单株。2021 年4 月28 日,将亲本、F1及F2种植于中国农业科学院棉花研究所东场试验基地(河南省安阳县)。3 个亲本分别种植3 行,群体1 种植约10 行,群体2 种植约70 行,行长均为8 m,行距为0.80 m,株距为0.25 m。田间管理措施同当地常规棉田。
1.2 苞叶表型调查
开花后(7 月中旬至8 月底),调查棉花苞叶表型。与T582 表型一致的记为窄卷苞叶,与TM-1 或3-79 表型一致的记为正常苞叶,并进行卡方(χ2)适合性检验,以确定F2群体的苞叶性状是否符合3∶1 的分离比。
1.3 分子标记的开发
根据Li 等[4]的定位结果,窄卷苞叶目标基因被定位在A03 染色体长臂上分子标记NAU3016和NAU3172 之间,目标基因与这2 个分子标记的遗传距离分别为0.3 cM 和4.7 cM。基于此,本研究结合海岛棉3-79[9]和陆地棉TM-1[10]参考基因组数据,利用IGV(integrative genomics viewer)软件来查找Indel 位点,选取插入或缺失10 bp 以上的位点,共设计了20 对Indel 分子标记引物。所有引物序列均由DNAMAN 软件设计获得,由生工生物工程(上海)股份有限公司合成(引物序列见表1)。
采用十六烷基三甲基溴化铵(Cetyltrimethylammonium bromide,CTAB)法提取T582 和3-79亲本的基因组DNA。采用TaqDNA 聚合酶(购自康为世纪生物科技股份有限公司)进行聚合酶链式反应(polymerase chain reaction,PCR),PCR 反应体系:缓冲液10 μL,模板DNA 1 μL,10 μmol·L—1正反向引物各1 μL,加ddH2O 至20 μL。PCR反应程序:94 ℃5 min;94 ℃30 s,55 ℃30 s,72 ℃1 min,35 个循环;72 ℃5 min 。将PCR 产物在3%(质量分数)的琼脂糖凝胶中电泳分离,以筛选出在亲本间有多态性的Indel 分子标记。
1.4 精细定位
利用筛选出的多态性Indel 分子标记,以提取的亲本及群体2 单株的基因组DNA 为模板进行PCR 扩增,利用琼脂糖凝胶电泳分析亲本及F2各个单株的扩增条带类型,统计交换单株数量,精细定位目的基因候选区间。DNA 提取、PCR扩增及琼脂糖凝胶电泳方法同1.3。
1.5 定位区间内候选基因预测
根据精细定位结果得到候选区间,在棉花功能基因组学数据库(https://cottonfgd.net/)中根据陆地棉TM-1 参考基因组[10]获得候选区间内功能基因的注释信息。从棉花多组学数据库(http://cotton.zju.edu.cn/)获取候选区间内的基因在TM-1 不同组织器官(根、茎、叶、花托、花瓣、花药、雌蕊、花萼、副萼、发育的胚珠和纤维)中的转录组测序数据,基于FPKM (fragments per kilobase of exon model per million mapped reads,每千个碱基的转录每百万映射读取的片段)对基因表达量数据进行log2(FPKM+1)标准化处理,使用R软件pheatmap 包绘制基因表达量的热图。
2 结果与分析
2.1 苞叶性状调查及遗传分析
对2 个F2群体进行田间性状调查,选取开花后15 天(15 days post anthesis,15 DPA)和30 DPA苞叶进行表型统计分析(图1)。群体1 中正常苞叶单株有285 株,窄卷苞叶单株有85 株,卡方适合性检验结果显示,分离群体的性状分离比为3∶1(χ2=0.86<χ20.053.84),符合孟德尔遗传分离定律。群体2 中正常苞叶单株2 015 株,窄卷苞叶单株652 株,卡方适合性检验结果显示,分离群体的性状分离比为3∶1(χ2=0.45<χ20.05=3.84),符合孟德尔遗传分离定律(表2)。因此,棉花窄卷苞叶基因fg为隐性单基因调控,与前人的研究结果[5]一致。

图1 正常苞叶和窄卷苞叶形态对比Fig.1 Morphology comparison of normal bract and frego bract

表2 F2 作图群体中窄卷苞叶和正常苞叶分离比Table 2 Segregation ratio of frego bract and normal bract in the F2 mapping populations
2.2 Indel 标记的开发与筛选
根据Li 等[4]对fg的定位结果,本研究在A03染色体上物理区间1.87~3.85 Mb 内,开发了20对Indel 分子标记,在亲本T582 和3-79 之间进行多态性筛选,共筛选到6 对具有多态性的Indel分子标记,分别为M1、M2、M3、M4、M5 和M6(引物序列信息参见表1)。
2.3 窄卷苞叶基因fg 的精细定位
利用筛选出的6 对多态性Indel 分子标记,首先对群体2 的300 个单株(其中正常苞叶单株237 株,窄卷苞叶单株63 株)的基因组DNA 进行扩增检测,将候选区间缩小在分子标记M1 与M4 之间,物理距离为861 kb(图2)。
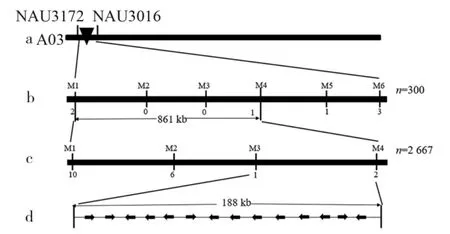
图2 fg 基因的定位Fig.2 Mapping of fg gene
利用M1、M2、M3、M4 分子标记对群体2 中2 667 个单株的基因组DNA 进行检测,根据重组数据,进一步将候选区间缩小至分子标记M3 与M4 之间,分子标记序列被锚定到陆地棉遗传标准系TM-1 基因组[10]A03 染色体2 757 376~2 946 245 bp,物理距离为188 kb(图2)。
2.4 候选区间基因预测
结合基因组注释结果,188 kb 定位区间内存在14 个功能注释基因(表3)。其中,Gh_A03G021400、Gh_A03G021500编码乙醛酸/ 羟基丙酮酸还原酶HPR3;Gh_A03G021600编码RALF-like 26 蛋白;Gh_A03G021700编码STAY-GREEN 1 蛋白(叶绿体);Gh_A03G021800编码 Agamous-like MADS-box 蛋白 AGL19;Gh_A03G021900编码同源异型盒蛋白knotted-1-like 7;Gh_A03G022000、Gh_A03G022100、Gh_A03G022200、Gh_A03G022300、Gh_A03G022400、Gh_A03G022500均编码抗病蛋白;Gh_A03G022600编码双功能dTDP-4-脱氢鼠李糖-3,5-差向异构酶/dTDP-4-脱氢鼠李糖还原酶;Gh_A03G022700编码羟甲基戊二酰辅酶A 合酶。

表3 定位区间内候选基因的注释信息Table 3 Annotation information of candidate genes in the mapping interval
根据这14 个功能注释基因在棉花不同组织器官中的相对表达量数据,在花萼和副萼中Gh_A03G021700、Gh_A03G021900、Gh_A03G022600、Gh_A03G022700基因的表达量较高;Gh_A03G021500、Gh_A03G022600、Gh_A03G022700基因在3DPA、5DPA 胚珠和10DPA纤维中高表达;Gh_A03G021900基因在花药、雌蕊、10 DPA 胚珠和20 DPA 的纤维中高表达;Gh_A03G021400、Gh_A03G021600、Gh_A03G021800、Gh_A03G022000、Gh_A03G022100、Gh_A03G022200、Gh_A03G022300、Gh_A03G022400、Gh_A03G022500基因在不同组织器官(根、茎、叶、花托、花瓣、花药、雌蕊、花萼、副萼、发育的胚珠和纤维)中的表达量均较低(图3)。
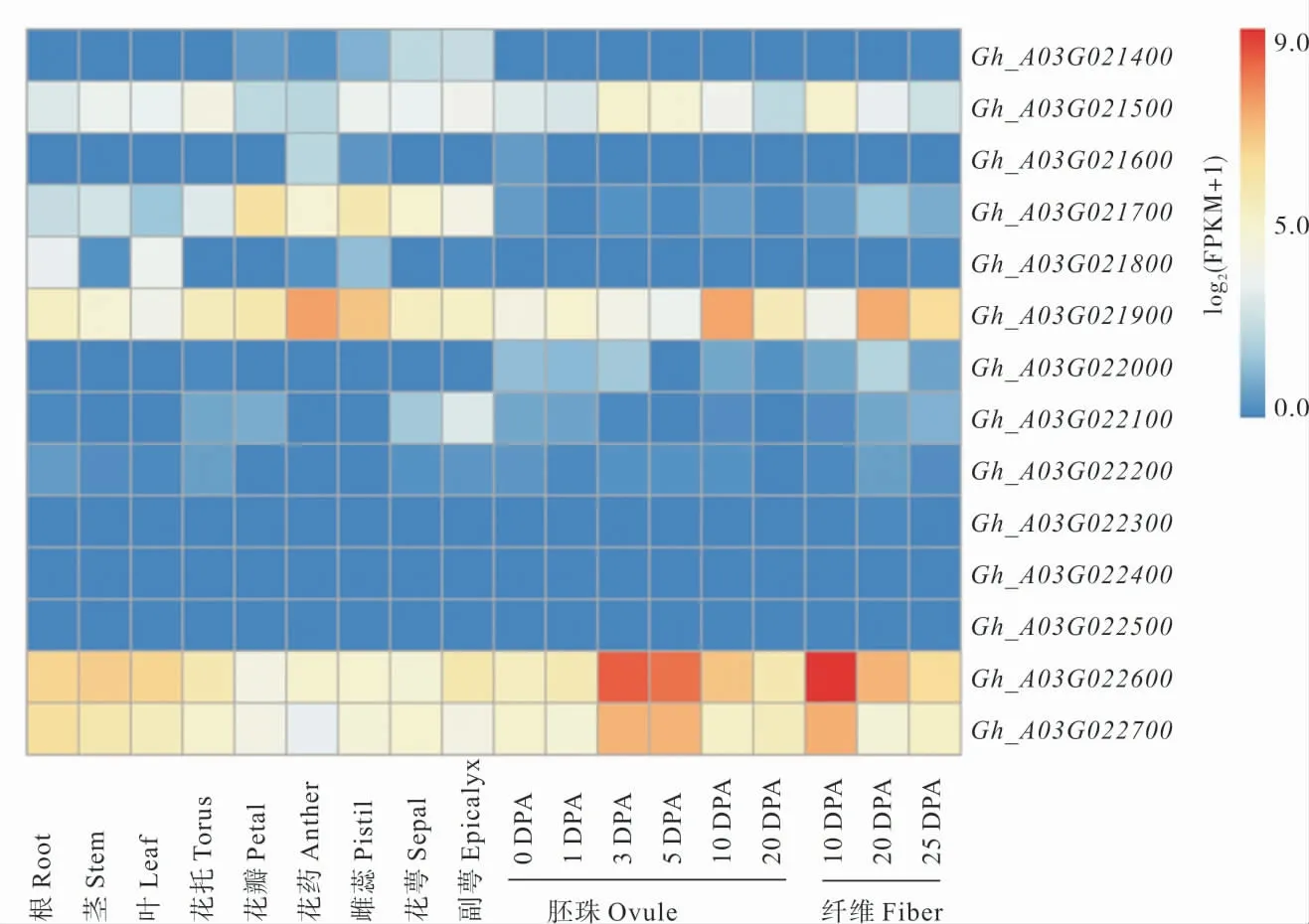
图3 定位区间内14 个注释基因的表达模式分析Fig.3 Expression pattern analysis of 14 annotated genes in the mapping interval
3 讨论
新疆是我国最大的机采棉生产基地。机采技术减少了劳动力投入,降低了棉花生产成本,大大提高了棉农的植棉经济收益,但机采棉普遍含杂率较高,给棉花的后续加工带来了一定的困难。原棉杂质主要包括干枯叶片、干枯苞叶和异性纤维等;其中干枯叶片和异性纤维能够通过及时采摘吐絮籽棉,严格遵守采收加工规定等方法得到有效控制;而干枯苞叶难以剔除,原因在于苞叶紧贴棉铃,吐絮后苞叶虽然枯死但仍紧贴在籽棉上不能自行脱落,在机械采摘时易将其混入籽棉[11]。棉花窄卷苞叶突变体在吐絮期表现为苞叶自然下卷,至采收期干枯苞叶与籽棉分离,且处在铃壳之下,因此可以降低机采棉的含杂率,利于提高棉花纤维品质。
棉花窄卷苞叶突变体表现为苞叶狭长、扭曲翻转生长,花蕾和棉铃裸露,植株对棉铃象甲、棉铃虫、棉红铃虫、金刚钻等害虫具有一定抗性[4,12]。杀虫剂对窄卷苞叶棉株的使用效果更好,研究表明,喷洒药剂后附着在此类花苞和棉铃上的杀虫药剂量是正常苞叶的7 倍[13]。本研究对棉花窄卷苞叶基因fg进行了精细定位,并对定位区间内的14 个候选基因进行了功能注释,发现有6个基因被注释为抗病蛋白,但其具体的作用机制尚不清楚。另外,有研究表明窄卷苞叶基因对棉花生育期、产量、衣分、单株果枝数、蕾铃脱落率和纤维品质均无明显不良效应[14-16]。综上,棉花窄卷苞叶突变体具有降低机采棉含杂率、增强植株抗虫性等优势,且对棉花产量和纤维品质无显著不利影响,具有一定的育种利用价值。
近年来,棉花基因组学的快速发展为棉花功能基因的定位及克隆奠定了坚实的基础[10,17-20],混合分组分析法[21]、全基因组关联分析[22](genomewide association study, GWAS)等方法也逐渐成为挖掘功能基因的重要手段,有效促进了棉花功能基因的研究进程。在前人研究基础上,本研究将棉花fg基因定位在A03 染色体上分子标记M3和M4 之间,区间大小为188 kb;区间内有14 个注释的功能基因,其中Gh_A03G021700、Gh_A03G021900、Gh_A03G022600、Gh_A03G022700基因在花萼、副萼中高表达,因此推测这4 个基因为可能的窄卷苞叶候选基因。未来将通过进一步的精细定位、图位克隆、功能验证等方式确定棉花窄卷苞叶的调控基因并解析其作用机制。
4 结论
棉花窄卷苞叶由隐性单基因调控。在前人工作基础上,利用陆地棉T582 和海岛棉3-79 构建F2分离群体,将窄卷苞叶基因fg精细定位在A03 染色体上分子标记M3 与M4 之间,物理距离为188 kb。该区间内存在14 个功能注释基因,其中Gh_A03G021700、Gh_A03G021900、Gh_A03G022600和Gh_A03G022700基因在花萼和副萼中高表达。本研究为棉花窄卷苞叶基因fg的图位克隆和育种利用奠定了基础。
猜你喜欢
安徽农学通报(2022年6期)2022-04-07 21:30:29
今日农业(2021年15期)2021-11-26 03:30:27
今日农业(2021年11期)2021-08-13 08:53:24
今日农业(2019年13期)2019-01-03 15:05:47
畜牧与饲料科学(2018年3期)2018-05-08 02:48:35
上海农业学报(2017年3期)2017-04-10 12:39:20
湖南林业科技(2017年1期)2017-02-06 05:28:55
上海农业学报(2016年2期)2016-10-27 00:49:47
湖南农业科学(2014年22期)2014-09-28 03:23:28
遗传(2014年3期)2014-02-28 20:58:49
