
抑制GhSAMS 基因表达降低陆地棉对棉铃虫的抗性
2023-08-23 07:42杨利艳李芳蒲哲王创云秦丽霞
棉花学报 2023年3期
杨利艳,李芳,蒲哲,王创云,秦丽霞*
(1.山西师范大学生命科学学院,太原 030031;2.山西农业大学农学院,太原 030031)
棉花是我国重要的经济作物之一,在国民经济中占有重要地位。陆地棉(Gossypium hirsutum)是棉花的主要栽培种[1-2]。棉花在整个生育期经常遭到各种虫害的威胁,导致棉花产量和纤维品质严重下降。我国棉田害虫有300 多种,主要虫害有30 余种[3],其中棉铃虫(Helicoverpa armigera)是蕾铃期的主要害虫,加强棉铃虫的防治对确保棉花丰收至关重要[4]。我国科研工作者已成功利用基因工程手段创制了多个转苏云金芽孢杆菌(Bacillus thuringiensis,Bt)基因抗虫棉品种(系),大大降低了棉铃虫对棉花的危害。但长期使用单一或少数几个抗虫基因必然会加速害虫产生抗性。近年来,为有效防治虫害,减轻农业生产经济损失,化学农药的投入和使用量不断增加,造成严重的田间污染,同时导致害虫产生更强的耐药性,进一步提高了虫害防治工作的难度并且增加了投入费用。因此,挖掘更多的抗虫基因对于提高作物的抗虫性及延缓害虫抗性的产生具有重要意义。
在植物体内,S- 腺苷甲硫氨酸合成酶(S-adenosylmethionine synthetase, SAMS)作为辅助因子参与细胞内的多种生理生化反应。SAMS是主要的甲基供体,参与蛋白质、核酸、多糖和脂肪酸的转甲基反应;SAMS 参与植物细胞专一性的代谢途径,是植物激素乙烯生物合成的前体物质[5],也是细胞壁成分相关的苯丙烷合成代谢所必需的。植物体内存在多个SAMS 编码基因[6]。前人研究表明,陆地棉SAMS基因家族有16 个成员,其启动子区存在多种参与防御和应激反应的顺式作用元件[7]。因此,陆地棉SAMS基因可能在多种胁迫响应中发挥重要作用。石蒜(Lycoris radiata)SAMS基因的转录水平在盐胁迫下明显提高,过表达SAMS基因可提高大肠杆菌(Escherichia coli)的耐盐性[8]。大豆(Glycine max)SAMS家族基因在涝害胁迫的早期表达量降低,在干旱胁迫的后期则上调表达[9]。陆地棉在接种黄萎病菌后,SAMS基因的表达量逐渐增加,表明其可能在抗黄萎病防卫反应中发挥重要作用[10]。利马豆(Phaseolus lunatus)在受到二斑叶蛾侵袭或茉莉酸处理后,叶片中SAMS的表达量均增加,推测SAMS可能通过茉莉酸途径在虫害响应中发挥作用[11]。斜纹夜蛾(Spodoptera litura)幼虫取食转GmSAMS1烟草叶片后,幼虫的相对生长率显著降低,表明大豆GmSAMS1基因增强了转基因烟草对斜纹夜蛾的抗性[12]。
活性氧(reactive oxygen species,ROS)是植物代谢的副产物。在正常情况下,植物体内ROS 的产生和清除受抗氧化系统的精细调控而处于动态平衡。当植物遇到非生物或生物胁迫时,植物体内ROS 的动态平衡被打破,导致植物体内积累大量的ROS。研究表明,以H2O2为代表的ROS在植物对非生物及生物胁迫的响应中起着重要作用[13-14]。
本课题组前期研究发现,大刍草(Zea maysssp.parviglumis)被粘虫 (Mythimna separateWalker)取食后,叶片中BtSAMS1的表达量较取食前增加了12.17 倍,由此我们推测BtSAMS1基因可能参与植物的抗虫防御响应[15-16]。基于此,本研究以陆地棉品种R15 为材料,分析了Bt SAMS1的同源基因GhSAMS在棉铃虫取食后的表达模式,并利用病毒诱导的基因沉默(virus-induced gene silencing, VIGS)技术抑制GhSAMS的表达,分析了沉默植株在受到棉铃虫取食后的表现,探究GhSAMS基因的抗虫功能,为提高棉花对棉铃虫的抗性提供候选基因资源,为延缓棉铃虫抗性的发生奠定基础。
1 材料与方法
1.1 材料
1.1.1植物材料及生长条件。供试棉花材料为常规陆地棉品系R15,由棉花生物育种与综合利用全国重点实验室 (原棉花生物学国家重点实验室)郝福顺教授提供。将棉花种子用98%(质量分数)的浓硫酸脱绒,在清水中多次洗涤,用清水浸泡催芽12 h 后,种植于人工气候室,培养条件为:温度(26±1)℃,光周期为16 h 光照/8 h 黑暗,相对湿度为70%。待子叶完全展平,挑选长势一致的植株用于后续实验。
1.1.2载体和菌株。pTRV1 辅助质粒由山西师范大学李柯老师惠赠,pNC:TRV2 质粒由中国热带农业科学院热带生物技术研究所言普老师提供,pTRV2::GhCLA1重组质粒由棉花生物育种与综合利用全国重点实验室郝福顺教授提供。大肠杆菌DH5α 和农杆菌GV3101 为本实验室保存。
1.1.3棉铃虫。本研究所用棉铃虫为购自河南全应商贸有限公司(河南郑州)的人工饲养品系。将棉铃虫在人工气候培养室培养至2 龄初期用于试验。饲养条件同1.1.1。
1.2 GhSAMS 的表达模式
当棉花幼苗长出2 片真叶时,选取长势一致的12 株棉花,每株放3 头2 龄初期的棉铃虫,取食24 h、48 h、72 h 后,对有明显虫食痕迹的棉花叶片进行取材,以未取食叶片为对照(取食0 h),置于液氮中速冻,于—80 ℃冰箱保存,用于后续试验。
提取棉花叶片总RNA,并反转录合成第一链cDNA。棉花总RNA 提取试剂盒及反转录试剂盒均购自北京全式金生物技术有限公司,具体操作步骤参考试剂盒说明书。采用Perfect-StartTMGreen qPCR SuperMix(+DyeⅠ/+DyeⅡ)试剂盒(北京全式金生物技术有限公司),以棉花GhUBQ7为内参基因,进行实时荧光定量聚合酶链式反应(real-time quantitative polymerase chain reaction,qRT-PCR)。引物序列见表1。每个样本进行3 次重复。使用2—ΔΔCt 法计算目的基因的相对表达量。
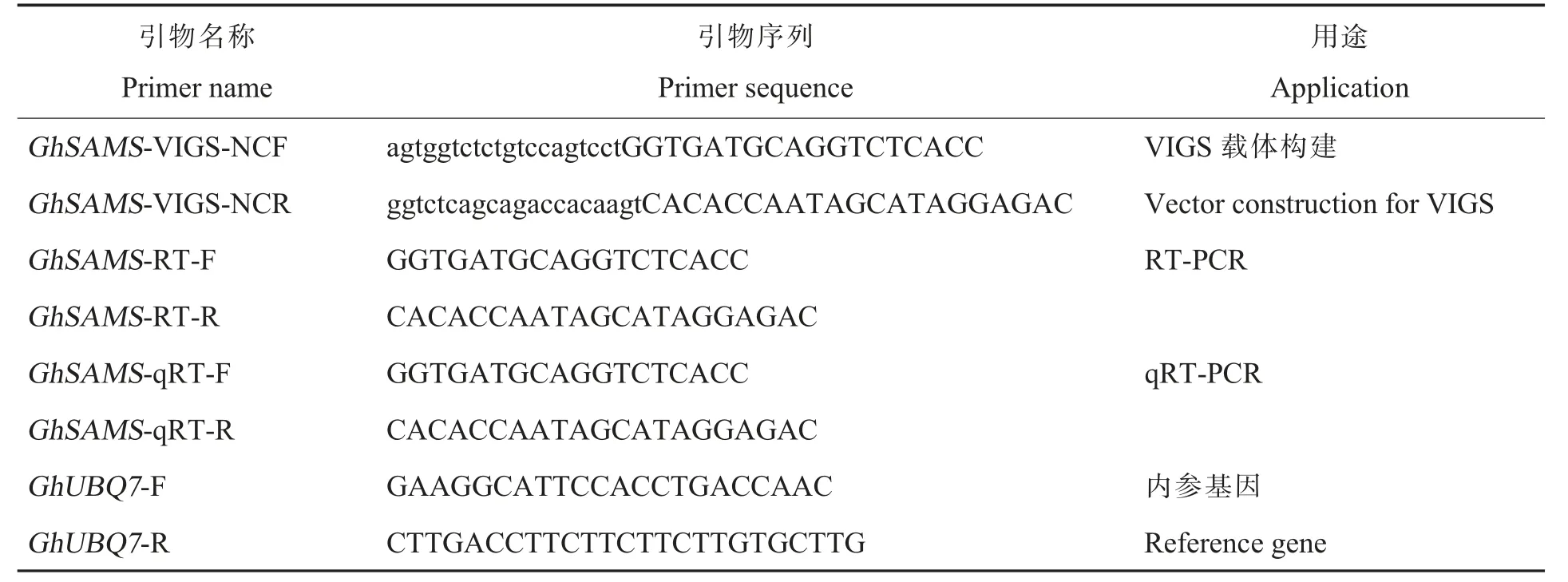
表1 本研究中所使用的引物序列Table 1 Primer sequences used in this study
1.3 pTRV2::GhSAMS 载体构建
以实验室前期筛选到的对粘虫取食强烈响应的BtSAMS1基因(基因ID:MK894430.1)为查询序列,在美国国家生物技术信息中心(National Center for Biotechnology Information, NCBI)中进行Blastp 比对。在Conserved Domain(CDD)[17]数据库(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)中查询鉴定棉花SAMS 蛋白保守结构域,去除无SAMS 典型保守结构域的蛋白序列,获得陆地棉SAMS基因204 bp 的保守序列。
提取棉花幼苗叶片总RNA,并反转录合成第一链cDNA,方法同1.2。根据SAMS基因保守序列,使用SnapGene 软件设计引物,并在上下游引物的5’端分别加上通用接头序列,引物序列见表1。以cDNA 为模板,进行PCR 扩增,获得目的片段,所用试剂盒为2×TransStartFastPfu PCR SuperMix(北京全式金生物技术有限公司)。PCR 反应体系为:cDNA 模板1 μL、2×TransTartFastPfu PCR SuperMix 25 μL、上下游引物各1 μL,ddH2O 补足到50 μL。PCR 反应程序为:95 ℃预变性1 min;95 ℃变性20 s、55 ℃退火20 s、72 ℃延伸15 s;72 ℃延伸5 min。将纯化后的目的片段插入沉默载体pNC:TRV2 中,并转化至DH5α 菌株,经测序验证后得到重组载体pTRV2::GhSAMS。
1.4 重组载体的转化和农杆菌侵染
将pTRV1、pNC:TRV2 空载体、重组载体pTRV2::GhSAMS和pTRV2::GhCLA1通过冻融法分别转化至农杆菌GV3101。挑取单菌落进行PCR 检测,将阳性单克隆在LB 液体培养基(含50 μg·mL—1卡那霉素和20 μg·mL—1利福平)中培养,培养条件为28 ℃、200 r·min—1,待OD600为1.5 时收集菌体。使用农杆菌侵染液(含10 mmol·L—1MgCl2、10mmol·L—12-吗啉乙磺酸和200μmol·L—1乙酰丁香酮)进行重悬至菌液OD600为1.5 左右,于室温下黑暗静置4 h 后,将pTRV1 的农杆菌重悬液分别与pTRV2 空载体(CK)、pTRV2::GhSAMS和pTRV2::GhCLA1的农杆菌重悬液按照1∶1 体积比混匀[18]。选取子叶完全展开、真叶还未长出的棉花幼苗,使用无针头的注射器,在棉花子叶背面按压注射,叶片的浸染面积需达到70%以上。2 周后观察pTRV2::GhCLA1植株的白化现象。
1.5 GhSAMS 基因沉默效率检测
以棉花内参基因GhUBQ7为参照,分别提取侵染2 周后的CK 植株、pTRV2::GhSAMS植株和同一时期未进行侵染处理的野生型(wild type,WT)植株的叶片总RNA。以反转录后的cDNA为模板,采用半定量反转录聚合酶链式反应(reverse transcription polymerase chain reaction,RT-PCR)检测GhSAMS基因的沉默效率。
1.6 棉铃虫生长发育测定
将2 龄初期的棉铃虫饥饿处理12 h,记录饥饿处理后棉铃虫的体重(W0)和体长(L0)。选取长势一致的WT、CK、pTRV2::GhSAMS棉花叶片,分别放入做好标记的带有双层湿润滤纸的培养皿中,每组设置3 个培养皿,每个培养皿放入1片叶和1 头饥饿处理的棉铃虫,将培养皿放置在温度为28 ℃、光周期为16 h 光照/8 h 黑暗条件下,每天更换新鲜叶片。喂食5 d 后,测量棉铃虫的体重(W1)和体长(L1)。体重变化量(mg)=W1—W0,取3 次重复的平均值;体长变化量(mm)=L1—L0,取3 次重复的平均值。
1.7 棉铃虫取食偏好性观察
在同一个培养盒中放入长势一致的WT、CK、pTRV2::GhSAMS棉花叶片各3 片,在3 种叶片等距的位置放置20 头2 龄初期棉铃虫使其自由取食,于24 h 和48 h 后观察棉铃虫对3 种材料叶片的取食情况。共设置3 次生物学重复。
1.8 棉花叶片受害等级
选取长势一致的WT、CK、pTRV2::GhSAMS棉花叶片,分别放入铺有双层湿润滤纸的培养皿中,在每个培养皿中放入1 片叶和1 头饥饿处理12 h 的2 龄初期棉铃虫,然后将培养皿放置在温度28 ℃、光周期为16 h 光照/8 h 黑暗条件下培养5 d,每个处理进行3 次生物学重复。5 d 后观察叶片被取食状况,分析棉花叶片受害等级[19]。
1.9 叶片染色
细胞中的过氧化氢(hydrogen peroxide,H2O2)与二氨基联苯胺(3,3’-diaminobenzidine,DAB)反应,形成黄褐色沉淀,通过观察染色情况可分析组织细胞中H2O2的积累情况。分别取WT 和pTRV2::GhSAMS植株的叶片,每5 片叶放入一个烧杯中,加DAB 染液浸没棉花叶片,用锡箔纸包裹烧杯,于室温条件下在水平摇床上缓慢振荡(80~100 r·min—1)过夜。将叶片取出加入无水乙醇于沸水中煮5~10 min 后,待叶绿素完全脱去,置于无水乙醇中待后续观察[20]。
1.10 数据处理与分析
采用SPSS 19.0 软件进行单因素方差分析。采用邓肯氏新复极差法(Duncan’s)进行多重比较。
2 结果与分析
2.1 棉铃虫取食下GhSAMS 的表达模式分析
为了解棉铃虫取食后GhSAMS基因的表达模式,对棉花植株进行虫食胁迫处理,并利用qRT-PCR 分析目的基因的相对表达量。结果表明,GhSAMS基因受到棉铃虫的诱导表达。与处理0 h 相比,叶片被取食24 h、48 h 和72 h后,GhSAMS的表达量均显著增加。其中,取食24 h 后GhSAMS的表达量最高,取食48 h 后GhSAMS的表达量较24 h 显著降低,取食72 h 后该基因的表达量较48 h 显著升高(图1)。表明GhSAMS的表达受棉铃虫取食诱导,可能与棉花对棉铃虫的抗性响应有关。
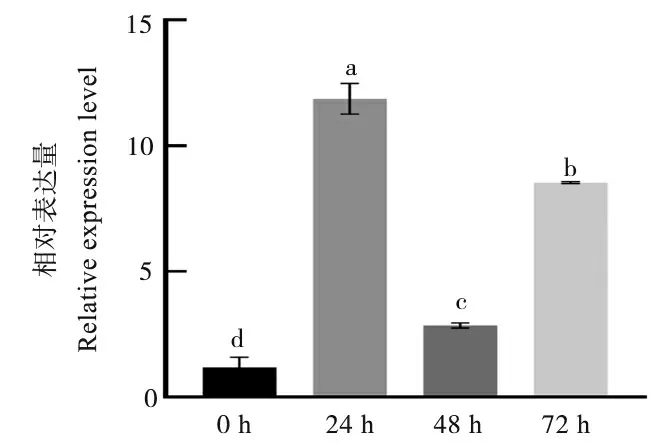
图1 GhSAMS 基因在棉铃虫胁迫下的表达分析Fig.1 Expression analysis of GhSAMS gene under the stress of H.armigera
2.2 GhSAMS 沉默植株的获得
pTRV2::GhCLA1棉花植株在侵染7 d 后真叶叶脉及边缘部分出现白化现象,到第14 天左右白化现象扩散至整个叶片,此时沉默效果最佳(图2A)。侵染后第14 天,利用半定量RT-PCR方法检测pTRV2::GhSAMS棉花叶片中GhSAMS基因的表达量,结果表明其表达受到明显抑制(图2B),说明成功获得了GhSAMS沉默植株,可用于后续的表型鉴定。

图2 pTRV2::GhSAMS 棉花植株表型及沉默效率检测Fig.2 Phenotype and silencing efficiency detection of pTRV2::GhSAMS plants
2.3 GhSAMS 沉默对棉铃虫生长发育的影响
取食5 d 后,取食WT 叶片的棉铃虫体重变化量显著高于CK,二者的体长变化量无显著差异;取食pTRV2::GhSAMS叶片的棉铃虫体重变化量显著高于WT 和CK,其体长变化量较CK 显著增加,与WT 差异不显著(图3)。说明棉铃虫在GhSAMS沉默棉株上发育更好,即抑制GhSAMS基因的表达有利于棉铃虫的生长发育。
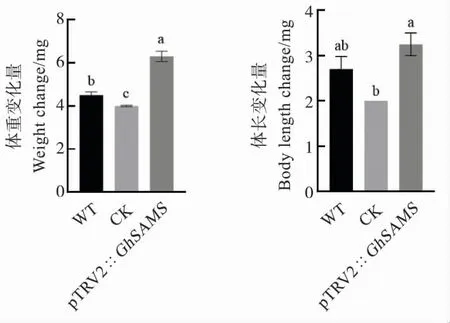
图3 不同棉花叶片对棉铃虫生长发育的影响Fig.3 Effects of different cotton leaves on the growth and development of H.armigera
2.4 棉铃虫取食偏好性观察
观察棉铃虫对不同类型棉花叶片的取食偏好,24 h 后75%的棉铃虫(15 头)趋向于pTRV2::GhSAMS棉花叶片,20%的棉铃虫(4头)趋向于WT 棉花叶片,5%的棉铃虫(1 头)趋向于CK 棉花叶片;48 h 后再观察,pTRV2::GhSAMS棉花叶片被取食程度最大(图4),以上结果表明棉铃虫更喜食GhSAMS沉默棉株叶片。

图4 棉铃虫对不同叶片的取食偏好性Fig.4 Feeding preference of H.armigera to differentcotton leaves
2.5 棉花叶片受害等级分析
观察WT、CK 和pTRV2::GhSAMS叶片被棉铃虫取食状况,进行叶片受害等级分析[19]。棉铃虫取食5 d 后,WT、CK 和pTRV2::GhSAMS叶片的受害等级分别为2 级、3 级和4 级,说明沉默棉花GhSAMS基因后,棉花叶片受棉铃虫的危害程度更大(图5)。

图5 棉铃虫取食后不同棉花叶片的受害情况Fig.5 Damage profile of different cotton leaves after H.armigera attack
2.6 叶片DAB 染色分析
为探究GhSAMS基因是否通过影响ROS的产生而参与提高对棉铃虫的抗性,通过DAB染色观察比较棉铃虫取食0 h 和24 h 的棉花叶片中的H2O2含量,结果表明,棉铃虫取食24 h后,与WT 相比,pTRV2::GhSAMS棉花叶片DAB 染色更深(图6)。表明沉默GhSAMS基因会导致棉花叶片中积累更多的H2O2。此外,我们还发现,H2O2在叶柄即叶脉交汇处积累量较多。

图6 DAB 染色分析棉花叶片的H2O2 含量Fig.6 H2O2 content in cotton leaves analyzed by DAB staining
3 讨论
SAMS 催化甲硫氨酸和三磷酸腺苷(adenosine triphosphate,ATP)合成S- 腺苷甲硫氨酸,是植物合成乙烯和多胺等过程中的关键酶,与许多抗逆反应相关的次生代谢物质的合成密切相关,在植物胁迫应答中扮演着重要角色。
目前诸多研究发现SAMS基因受多种生物及非生物胁迫诱导表达。余涛等[21]在烟草中克隆了一个SAMS基因,发现其受氧化胁迫、盐胁迫及高温胁迫诱导表达。Gupta 等[22]研究表明白杨树SAMS基因受高浓度CO2和O3的诱导表达。宋修鹏等[23]发现甘蔗ScSAM在黑穗病菌胁迫下上调表达。拟南芥、水稻SAMS基因在干旱胁迫下表达量升高[24]。番茄SAMl和SAM3受盐、甘露醇和脱落酸的诱导表达[25]。本研究发现陆地棉GhSAMS基因在棉铃虫取食后表达量明显升高,因此推测GhSAMS基因可能参与了抗棉铃虫响应。利用VIGS 技术沉默GhSAMS基因,发现基因沉默棉花植株更易受到棉铃虫的侵害,具体表现为:棉铃虫取食GhSAMS沉默棉株叶片后,其体重与体长的变化量均大于WT 和CK,棉铃虫更喜食GhSAMS沉默棉株叶片;GhSAMS沉默棉花叶片受棉铃虫的危害程度更大。综合分析表明GhSAMS基因对棉铃虫的抗性起正调控作用,未来可通过转基因过表达或基因编辑技术等进一步验证该基因的功能。本研究仅分析了离体棉花叶片对棉铃虫的抗性,后续可对棉花的蕾铃及整个植株对棉铃虫的抗性进行研究。此外,棉花的害虫并非只有棉铃虫,还有棉花蚜虫、烟粉虱、棉盲蝽等,GhSAMS基因是否参与对不同类型害虫的抗性响应,后续可进行系统研究。
前人研究表明,H2O2信号转导途径除了在植物的抗病反应中起着重要作用外[26-28],在植物抗虫响应中亦发挥着重要作用[29]。本研究中,对棉铃虫取食24 h 的棉花叶片进行DAB 染色,观察到pTRV2::GhSAMS棉花叶片组织中积累了更多的H2O2。表明沉默GhSAMS基因后,棉铃虫取食会诱导植株产生更多的H2O2,而H2O2的过量增加会破坏植株体内ROS 的动态平衡,从而对植物细胞的结构功能造成氧化损伤,导致pTRV2::GhSAMS棉花植株更易受到棉铃虫的取食侵害。
4 结论
GhSAMS基因受棉铃虫取食的诱导表达。利用VIGS 技术瞬时沉默GhSAMS基因导致棉花植株对棉铃虫的抗性降低,推测GhSAMS基因对棉铃虫的抗性起正调控作用。GhSAMS可作为提高棉花棉铃虫抗性的候选基因。
猜你喜欢
少儿科学周刊·儿童版(2022年10期)2022-06-27
空间科学学报(2021年1期)2021-05-22
小读者(2020年4期)2020-06-16
环境保护与循环经济(2017年5期)2018-01-22
新疆农垦科技(2016年2期)2016-08-21
中国果菜(2016年9期)2016-03-01
现代农业(2016年5期)2016-02-28
现代农业(2016年4期)2016-02-28
中国蔬菜(2015年9期)2015-12-21
现代农业(2015年5期)2015-02-28
