
延龄草总皂苷对卒中后认知障碍大鼠学习记忆能力和神经元突触损伤的影响
2023-08-23 10:39王刚杨丹段壬泽杨丽君陈娟赵方毓陈显兵
中国中医药信息杂志 2023年8期
王刚 ,杨丹 ,段壬泽 ,杨丽君 ,陈娟 赵方毓 ,陈显兵
1.湖北民族大学附属民大医院,湖北 恩施 445000; 2.湖北民族大学医学部,湖北 恩施 445000
卒中后认知障碍(post-stroke cognitive impairment,PSCI)是指在卒中后6个月内发生的从轻度认知损害到痴呆的一系列综合征,是临床常见的脑血管病变及神经退行性疾病[1]。2020年发布的《中国卒中报告》显示,我国卒中患病率为1.11%,年发病率为0.25%,病死率为0.15%,其中约1/3的卒中患者会经历PSCI[2]。
PSCI发病机制复杂,目前普遍认为,脑血管病变引起脑部缺血或出血,进而导致脑神经解剖结构损害是其主要发病机制[3]。在卒中期间,被剥夺正常代谢底物耗氧和耗能的神经元在几秒内停止正常运作,并在2 min内显示出结构性损伤的迹象[4]。随着能量依赖过程的加重,神经元无法维持其正常生理功能,导致细胞凋亡,最终损害感觉和运动功能。近年来,随着神经外科手术的发展,越来越多的患者在急性脑卒中后幸存下来,但大多数患者术后仍患有持续的感觉运动和认知功能障碍[5]。
延龄草为百合科延龄草属植物延龄草Trillium tschonoskiiMaxim的干燥根茎,味甘,性平,有小毒。研究发现,延龄草总皂苷具有抗氧化、延缓衰老、促进学习记忆等作用[6-7],但其对脑卒中后引发的神经疾病的作用研究较少。本研究采用改良线栓法制备PSCI大鼠模型,初步探讨延龄草总皂苷对PSCI大鼠学习记忆能力及海马神经元突触损伤的影响及作用机制,为PSCI临床治疗提供实验依据。
1 材料与方法
1.1 动物
清洁级6~7周龄雄性SD大鼠60只,体质量(200±20)g,三峡大学实验动物中心提供,动物许可证号SCXK ( 鄂) 2017-0012, 动物合格证号42010200005116。饲养于湖北民族大学医学部清洁动物房,温度23~25 ℃,相对湿度50%~60%,适应性饲养1周。本研究经湖北民族大学医学伦理委员会审批(202159)。
1.2 药物
延龄草采自恩施,由湖北民族大学中药实验室鉴定为百合科延龄草属植物延龄草的干燥根茎。药材洁净后,经干燥、粉碎、提取、药液过滤、浓缩干燥,最终得含量为61.48%的延龄草总皂苷,称取1 g延龄草总皂苷,加入100 mL双蒸水溶解备用。盐酸多奈哌齐片,批号0000001782,浙江华海药业,5 mg/片,将5 mg药片研粉,加入111 mL双蒸水溶解备用。
1.3 主要试剂与仪器
TUNEL细胞凋亡试剂盒(批号E-CK-A321,武汉伊莱瑞特生物),高尔基染色液(批号GMS80020.3,美国GENMED),尼氏染色液、SDS-PAGE快速配制试剂盒、BCA蛋白浓度测定试剂盒、Akt抗体(批号分别为C0117、P0012AC、P0010、AA326,上海碧云天生物),PSD95 抗体、SYN 抗体、Caspase-3 抗体、HRP标记山羊抗兔IgG、HRP标记山羊抗小鼠IgG(批号分别为ab18258、ab52636、ab32351、ab6721、ab6789,英国Abcam),Bcl-2抗体、Bax抗体(批号分别为A19693、A19684,武汉ABclonal),PI3K抗体、p-Akt 抗体(批号分别为4255S、4060S,美国Cell Signaling Technology),β-actin抗体(批号66009-1-Ig,武汉三鹰生物)。
JSZ5B型体视显微镜(江南公司),Top Scan2.00水迷宫系统(美国Clever Sys),VT1200S震荡切片机(德国Leica Biosystems公司),1510型全波长酶标仪(美国Thermo公司),Tanon-5200全自动化学发光成像系统(上海天能科技公司),5810R高速冷冻离心机(德国Eppendorf公司),Western blot电泳及转膜装置(美国Bio-Rad公司)。
1.4 分组、造模及给药
60只大鼠随机分为造模组50只和假手术组10只,造模组参照文献[8]采用改良线栓法制备PSCI大鼠模型,假手术组仅切开皮肤,分离左侧颈总动脉后立即缝合切口,不插入线栓。术后按照Longa 5分法[8]对大鼠进行神经功能评分,评分1~3分为造模成功,共得到30只造模成功大鼠。将成模大鼠随机分为模型组、延龄草总皂苷组和盐酸多奈哌齐组,每组10只,延龄草总皂苷组予10 mg/mL延龄草总皂苷溶液100 mg/kg灌胃,盐酸多奈哌齐组予0.045 mg/mL盐酸多奈哌齐溶液0.45 mg/kg灌胃,灌胃体积10 mL/kg,假手术组和模型组予等体积双蒸水灌胃,连续28 d。
1.5 Morris水迷宫实验
给药结束当天开始定位航行实验,系统自动将圆形水面分为4个象限,求生平台放在第3象限低于水面0.6~1 cm处,将大鼠面向池壁分别于第1、2、4象限放入,系统自动记录大鼠60 s内寻找平台所需时间,即逃避潜伏期。每日1次,连续5 d,第6日休息。第7日上午进行正式定位航行实验,将大鼠沿第1象限放入,记录大鼠逃避潜伏期。下午进行空间探索实验,撤去平台,将大鼠沿第1象限放入,记录大鼠60 s内穿越平台次数及目标象限停留时间比(第3象限停留时间÷总时间×100%)。
1.6 HE、尼氏染色
行为学实验结束后,每组随机选取2只大鼠,10%水合氯醛0.3 mL/100 g腹腔注射麻醉,经心脏灌注多聚甲醛固定后,断头取全脑,沿大脑视交叉处做4~5 mm冠状切片,将切片置于4%多聚甲醛中固定48 h。常规脱水、石蜡包埋后,行4 μm 石蜡切片,经HE、尼氏染色后,中性树胶封片,扫描切片,观察脑组织病理变化。
1.7 TUNEL染色
石蜡切片常规脱蜡后置于PBS中备用,滴加适量蛋白酶K工作液覆盖组织,37 ℃反应20 min。PBS洗涤切片3次,每次5 min,滴加适量1×DNaseⅠ Buffer工作液,室温平衡5 min,滴加适量DNaseⅠ工作液,37 ℃孵育10~30 min。PBS洗涤切片3次,每次5 min,滴加适量TdT Equilibration Buffer覆盖组织,37 ℃孵育10~30 min。滴加适量阳性标记工作液,湿盒37 ℃避光反应60 min,PBS洗涤3次,每次5 min,滴加适量DAPI,室温避光孵育5 min,PBS洗涤切片4次,每次5 min,滴加抗荧光淬灭封片剂,盖玻片封片,荧光显微镜下观察海马CA1区神经元凋亡情况。
1.8 高尔基染色
行为学实验结束后,每组随机选取3只大鼠进行高尔基染色。大鼠经心脏灌注生理盐水后,沿海马区冠状位做5 mm切块进行媒染,2周后将组织放入震荡切片机中,行40 μm切片,严格按试剂盒说明书将切好的组织依次放入染液中染色,明胶封片。采用Image Pro Plus 6.0软件对图像进行分析,计算树突棘密度。
1.9 Western blot检测
将各组剩余5只大鼠麻醉,断头取脑,冰上分离左侧海马,按1 mg组织加入9 μL裂解液、0.09 μL蛋白酶抑制剂进行组织匀浆、超声,4 ℃、12 000 r/min离心15 min,取上清液,BCA试剂盒检测蛋白浓度。制备12%分离胶,每孔上样4 μL,依次进行电泳、转膜、血清封闭15 min、TBST洗膜,将膜置稀释后的一抗中(PI3K 1∶1 000、p-Akt 1∶2 000、Akt 1∶1 000、Bcl-2 1∶1 000、Bax 1∶1 000、Caspase-3 1∶5 000、SYN 1∶5 000、PSD-95 1∶1 000),4 ℃孵育过夜。TBST 洗膜,置于稀释后的二抗中,室温孵育1 h,TBST洗膜,滴加ECL显色液进行曝光,采用GIS 1D图像分析软件计算蛋白灰度值。
1.10 统计学方法
2 结果
2.1 延龄草总皂苷对模型大鼠学习记忆能力的影响
与假手术组比较,模型组大鼠Morris水迷宫实验逃避潜伏期显著增加,穿越平台次数及目标象限停留时间比显著减少(P<0.01);与模型组比较,延龄草总皂苷组和盐酸多奈哌齐组大鼠Morris水迷宫实验逃避潜伏期显著减少,穿越平台次数及目标象限停留时间比显著增加(P<0.05,P<0.01)。见表1。
表1 各组大鼠Morris水迷宫实验结果比较(±s)

表1 各组大鼠Morris水迷宫实验结果比较(±s)
注:与假手术组比较,**P<0.01;与模型组比较,#P<0.05,##P<0.01
?
2.2 延龄草总皂苷对模型大鼠脑组织病理变化的影响
假手术组大鼠海马CA1 区及皮质区神经元饱满丰富、排列整齐、数量较多,细胞核着色清晰,未见明显细胞坏死及空泡化现象;模型组大鼠海马CA1区及皮质区神经元排列紊乱、数量较少,出现核固缩,尼氏体数量减少;延龄草总皂苷组及盐酸多奈哌齐组大鼠海马CA1 区及皮质区神经元较饱满、数量增多,空泡化及坏死细胞明显减少,尼氏体数量增加。见图1、图2。
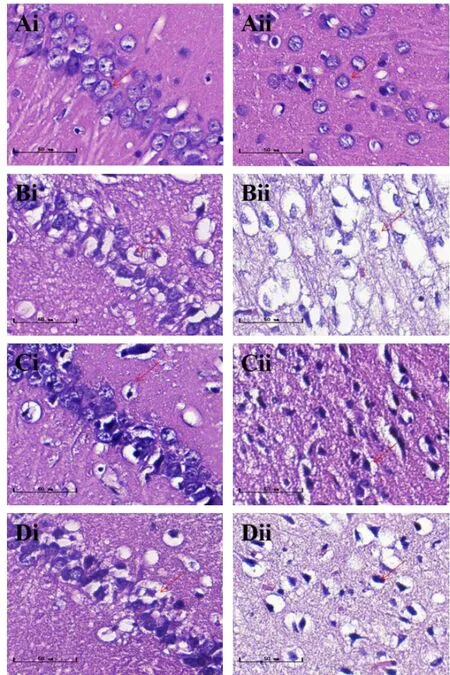
图1 各组大鼠脑组织形态(HE染色,标尺=50 μm)

图2 各组大鼠脑组织形态(尼氏染色,标尺=50 μm)
2.3 延龄草总皂苷对模型大鼠海马CA1区神经元凋亡的影响
TUNEL 染色蓝色为细胞核,绿色为凋亡细胞。与假手术组比较,模型组大鼠海马CA1区凋亡神经元数量增加;与模型组比较,延龄草总皂苷组和盐酸多奈哌齐组大鼠海马CA1 区凋亡神经元数量减少。见图3。

图3 各组大鼠海马CA1区神经元凋亡阳性表达(TUNEL染色,×200)
2.4 延龄草总皂苷对模型大鼠海马组织树突棘密度的影响
与假手术组比较,模型组大鼠海马组织树突棘密度显著减小(P<0.01);与模型组比较,延龄草总皂苷组和盐酸多奈哌齐组大鼠海马组织树突棘密度显著增加(P<0.01)。见图4。

图4 各组大鼠海马组织树突棘密度比较(高尔基染色,×1 000,±s,每组3只)
2.5 延龄草总皂苷对模型大鼠海马组织蛋白表达的影响
与假手术组比较,模型组大鼠海马组织Bcl-2、SYN、PSD-95、PI3K、p-Akt蛋白表达显著降低(P<0.01),Bax、Caspase-3蛋白表达显著升高(P<0.01);与模型组比较,延龄草总皂苷组和盐酸多奈哌齐组大鼠海马组织Bcl-2、SYN、PSD-95、p-Akt蛋白表达显著升高(P<0.01),Bax、Caspase-3蛋白表达显著降低(P<0.01,P<0.05),延龄草总皂苷组PI3K蛋白表达显著升高(P<0.01)。见图5、表2。

图5 各组大鼠海马组织Bcl-2、Bax、Caspase-3、SYN、PSD-95、PI3K、Akt、p-Akt蛋白免疫印迹
表2 各组大鼠海马组织Bcl-2、Bax、Caspase-3、SYN、PSD-95、PI3K、Akt、p-Akt蛋白表达比较(±s)

表2 各组大鼠海马组织Bcl-2、Bax、Caspase-3、SYN、PSD-95、PI3K、Akt、p-Akt蛋白表达比较(±s)
注:与假手术组比较,**P<0.01;与模型组比较,#P<0.05,##P<0.01
?
3 讨论
认知是脑的高级功能,海马及大脑皮层是大脑记忆网络的重要组成部分。大脑缺血后可引起海马体及皮质区神经元损伤,从而导致认知功能减退或永久性记忆损害,是脑卒中最严重的功能障碍之一[9-10]。本研究采用Morris水迷宫实验评价大鼠学习记忆能力,结果显示,模型组大鼠逃避潜伏期显著增加,目标象限停留时间比显著减少,表明造模后大鼠出现认知功能障碍。经延龄草总皂苷干预后,大鼠逃避潜伏期显著减少,目标象限停留时间比显著增加,表明延龄草总皂苷对急性脑缺血引起的认知功能障碍具有明显改善作用。
神经元损伤的常见原因之一是缺氧诱导的细胞凋亡[11-12]。急性缺血性脑卒中后,大脑中动脉供血区域的循环氧气输送减少,造成局部神经元快速凋亡。本研究采用HE染色及尼氏染色对大鼠缺血侧脑组织进行观察,模型组大鼠大脑皮质区由缺血、缺氧造成的神经元坏死最为明显,核固缩及空泡化现象突出,海马CA1区出现神经元排列疏松、尼氏体数量减少、核固缩现象。经延龄草总皂苷干预后,大脑皮质区神经元空泡化及核固缩现象减轻,海马CA1区空泡化不明显,说明延龄草总皂苷对缺血、缺氧造成的神经元丢失及损伤具有抑制作用。
细胞凋亡在脑缺血发作数小时后开始,受Bcl-2家族、Caspase家族等多基因调控,Bcl-2是抗凋亡蛋白,可被Bax干扰,导致细胞凋亡。Caspase-3是大脑发育的重要蛋白,在介导神经元凋亡中起着重要作用。脑缺血后,Bcl-2表达减少,Bax表达增加,Bcl-2和Bax结合形成凋亡二聚体,从而激活Caspase,触发依赖性凋亡级联反应,诱导细胞凋亡。本研究发现,模型组大鼠海马CA1区凋亡细胞明显增加,延龄草总皂苷干预后,海马CA1区凋亡细胞明显减少;Western blot结果显示,延龄草总皂苷能上调海马组织Bcl-2蛋白表达,下调Bax及Caspase-3蛋白表达,提示延龄草总皂苷能抑制神经元凋亡,对神经元起保护作用。
神经元和突触是海马依赖性学习记忆的生物学基础。研究显示,缺血性脑卒中损害神经网络和细胞间信号传递的完整性,并导致认知障碍[13]。PSD-95 和SYN是突触可塑性的重要指标,参与兴奋性突触的成熟和突触接触的稳定[14]。树突棘的可塑性是认知功能的结构基础,与神经元回路的重新连接有关,树突棘可塑性增强有助于皮质功能恢复[15]。PI3K/Akt信号通路是细胞代谢、生长、凋亡等生理活动的经典通路,可通过调控氧化应激、细胞凋亡等过程参与缺血性脑卒中发生发展[16-17]。缺血性脑卒中发生后,人体释放神经细胞生长因子、整合素等激活酪氨酸激酶受体,磷酸化PI3K和Akt,触发PI3K/Akt通路激活。本研究发现,模型大鼠予延龄草总皂苷后,海马组织PSD-95、SYN、PI3K、p-Akt蛋白表达和树突棘密度均显著增加,表明延龄草总皂苷可以激活PI3K/Akt信号通路,从而对PSCI 大鼠海马神经元突触损伤发挥保护作用。
综上所述,延龄草总皂苷可以改善PSCI大鼠学习记忆能力,减轻神经元突触损伤,其机制可能与激活PI3K/Akt信号通路,上调Bcl-2蛋白表达,下调Bax、Caspase-3蛋白表达,抑制细胞凋亡有关。本研究从神经元凋亡和突触损伤角度初步探讨延龄草总皂苷对PSCI大鼠的保护作用及机制,鉴于PI3K/Akt信号通路调控靶点的复杂性及中药治疗多靶点、多途径等特点,今后将从其他途径进行深入研究。
猜你喜欢
现代食品科技(2022年8期)2022-09-02
中学生数理化·高一版(2022年3期)2022-04-05
世界最新医学信息文摘(2021年12期)2021-06-09
临床荟萃(2020年1期)2020-12-13
中成药(2018年9期)2018-10-09
电子测试(2018年11期)2018-06-26
中成药(2017年9期)2017-12-19
中学生数理化·七年级数学人教版(2017年4期)2017-07-08
中成药(2017年6期)2017-06-13
安徽医科大学学报(2016年12期)2017-01-15
