
高温胁迫对红彩瑞猎蝽存活率及捕食作用的影响
2023-08-18 10:12:54曾涛游梓翊夏长剑孙郑陈德鑫邓海滨
中国烟草科学 2023年3期
曾涛 游梓翊 夏长剑 孙郑 陈德鑫 邓海滨


摘要:為探明高温条件下红彩瑞猎蝽耐受能力及对斜纹夜蛾幼虫的控害潜能,在室内测定了红彩瑞猎蝽经不同高温胁迫48 h 后的卵孵化率,5龄若虫羽化率,3、4、5龄若虫和雌、雄成虫存活率,评估了红彩瑞猎蝽不同虫态在高温条件下24 h 内对斜纹夜蛾3龄幼虫的捕食能力。结果表明,高温胁迫对红彩瑞猎蝽各虫态的存活率、卵孵化率和羽化率有显著影响,经42℃处理48 h 后红彩瑞猎蝽雌成虫和5龄若虫的存活率最高,达66.67%;红彩瑞猎蝽存活率、卵孵化率和羽化率在27~33℃时随温度升高而增加,在33℃后均随着温度升高而降低,36℃后存活率、卵孵化率和羽化率均下降明显。红彩瑞猎蝽各虫态在30~33℃范围对斜纹夜蛾的捕食量和捕食效能随温度升高而增大,在33~39℃范围,捕食量和捕食效能随温度升高而降低,红彩瑞猎蝽各虫态对斜纹夜蛾的捕食功能反应类型符合 Holling Ⅱ型,随着猎物密度的增加,对猎物的搜寻效应逐渐减弱,个体间存在竞争和相互干扰作用。可见红彩瑞猎蝽对高温逆境有一定的耐受能力,其适宜生存的高温范围为33~36℃。关键词:高温胁迫;红彩瑞猎蝽;存活率;捕食功能;斜纹夜蛾
中图分类号: S435.72 文献标识码: A 文章编号:1007-5119(2023)03-0053-09
Effects of High Temperature Stress on the Survival and Functional Response of Rhynocorisfuscipes to the Larvae of Spodoptera litura
ZENG Tao1, YOU Ziyi1, XIA Changjian2, SUN Zheng1, CHEN Dexin2, DENG Haibin1*
(1. Tobacco Research Institute of Guangdong Province, Shaoguan, Guangdong 512029, China;2. Haikou Cigar Research Institute,Hainan Provincial Branch of China National Tobacco Corporation (CNTC), Haikou 571100, China)
Abstract: To evaluate the survival and predation rate of the predatory bug Rhynocorisfuscipes against Spodoptera litura under high temperature stress, hatching rate of eggs, emergence rate of 5th nymphs, survival rate of different life stages of R. fuscipes were measured in the laboratory, after being subjected to high temperature stress at 30℃, 33℃, 36℃, 39℃ and 42℃ for 48 hours respectively. Daily prey consumption of different life stages of R. fuscipes on S. litura larvae at 30℃, 33℃, 36℃, and 39℃ respectively in 24 hours was recorded. The results showed that there was a significant effect of high temperature on survival, eclosion and egg hatching rate, also the prey consumption of R.fuscipes. The survival rate of female adults and 5th instar nymphs was 66.67% after 48 hours of high temperature stress at 42℃. Survival rate, egg hatching rate and eclosion rate increased with temperature from 27℃ to 33℃, and decreased gradually with the increase of temperature after 33℃. Survival rate, egg hatching rate, and eclosion rate all decreased significantly when the temperature exceeds 36℃. The predation efficiency of predation quantity decreases gradually with the increase of temperature in the range of 33-39℃. The predatory functional response type of different states of R. fuscipes to S. litura conforms to Holling II disc equations. The searching efficiency of R. fuscipes against S. litura larvae decreased with increasing prey density, and the predation amount and predation rate of R. fuscipes decreased gradually after the interference presented. The results showed that R.fuscipes has a certain tolerance to high temperature stress, and its suitable temperature range for survival is 33-36℃.
Keywords: high temperature stress; Rhynocorisfuscipes; survival rate; functional response; Spodoptera litura
红彩瑞猎蝽(Rhynocorisfuscipes)属半翅目(Hemiptera)猎蝽科(Reduviidae)昆虫[1]。在印度、越南、马来西亚、老挝和日本等国以及我国贵州、西藏、四川、江西、湖南、福建、广东、云南和海南等地均有分布[2]。据国外研究报道,红彩瑞猎蝽捕食范围广,能捕食棉花上的棉二点红蝽(Dysdercuscingulatus)、棉铃虫(Helicoverpaarmigera)、棉斑实蛾(Eariasinsulana)、斜纹夜蛾(Spodoptera litura )和水稻上的稻纵卷叶螟(Cnaphalocrocis medinalis )、二化螟( Chilo suppressalis)、褐飞虱(Nilaparvata lugens)等害虫,是重要的捕食性天敌[3-7]。在烟草上的研究表明,红彩瑞猎蝽对烟蚜(Myzuspersicae)、烟青虫(Helicoverpaassulta)和斜纹夜蛾有较强的控害能力[8-9]。红彩瑞猎蝽是南方烟区的优势天敌种群,邓海滨等[10]研究了温度对其发育历期的影响,郭义等[11]研究了低溫饲养对红彩瑞猎蝽生长发育的影响。南方烟田斜纹夜蛾发生高峰期恰逢高温季节,高温条件是否会影响红彩瑞猎蝽生长发育、存活率及其对斜纹夜蛾的捕食作用,目前未见相关研究报道。
温度胁迫耐受性研究是昆虫生态学研究中的一个热点,环境温度过高,会超出昆虫生存的适宜范围,极端高温甚至会导致昆虫的迅速死亡[12]。例如黑纹透翅花蝽(Montandoniolamoraguesi)在15~35℃范围内生长发育正常,而温度升至35℃时,死亡率显著提高[13]。温度变化对天敌昆虫捕食功能等也有显著影响,加州新小绥螨(Neoseiuluscalifornicus)对朱砂叶螨(Tetranychuscinnabarinus)雌成螨的瞬时攻击率在19~31℃温度范围内随温度的升高而逐渐减小[14]。军配盲蝽(Stethoconus japonicus )成虫对悬铃木方翅网蝽(Corythuchaciliatav)的捕食效能在15~35℃范围内随着温度的升高先增强后减弱[15]。为了更合理利用和保护现有天敌物种,有必要开展高温对天敌生长发育及捕食影响的评估研究。
本研究测定不同高温条件对红彩瑞猎蝽不同虫态存活率、卵孵化率和5龄若虫羽化率,以及对捕食斜纹夜蛾幼虫的影响,以期为夏季高温条件下保护红彩瑞猎蝽并利用该天敌对斜纹夜蛾进行防治提供理论依据。
1 材料与方法
1.1 供试材料
试验所需红彩瑞猎蝽和斜纹夜蛾均为广东省烟草科学研究所室内繁殖3代以上种群,试验所用人工气候培养箱型号为江南仪器厂 ARMA-580型,试验过程设定相对湿度(70±5)%,光周期16 h。
1.2 试验方法
1.2.1 不同高温对不同虫态猎蝽存活率、羽化率和卵孵化率的影响试验设27℃为对照温度,30℃、33℃、36℃、39℃和42℃等5个高温梯度处理。将单个卵块和单头不同虫态红彩瑞猎蝽用毛笔挑入28 mL 塑料杯(上底直径39 mm,下底直径30 mm,高32 mm ,杯盖有透气孔)中,塑料杯底放一小块湿润的脱脂棉,每个温度设4个重复,每个重复5 头猎蝽或5块卵块,在上述温度条件下处理48 h ,然后转入27℃培养箱继续观察,每隔8 h 观察记录红彩瑞猎蝽3、4、5龄若虫和雌、雄成虫的存活数量、卵孵化率,5龄若虫羽化率。
1.2.2 不同高温条件下红彩瑞猎蝽对斜纹夜蛾的捕食功能及搜寻效应因预备试验中观察到温度达到42℃时,斜纹夜蛾3龄幼虫在24 h 内出现自然死亡情况,故捕食功能试验设30、33、36和39℃4个温度处理,斜纹夜蛾3龄幼虫设置10、20、30、40、50头密度,观察在上述温度条件下24 h 内红彩瑞猎蝽3、4、5龄若虫和雌成虫对斜纹夜蛾3龄幼虫捕食量情况。每个处理重复6次。
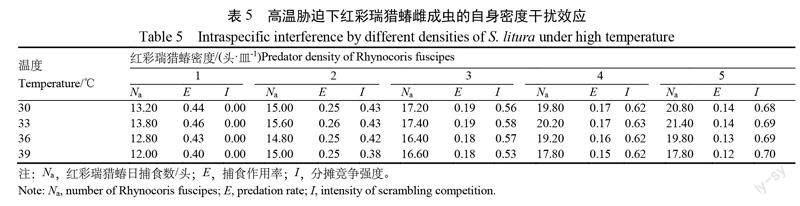
1.2.3 不同高温条件下红彩瑞猎蝽捕食斜纹夜蛾自身密度干扰效应将雌成虫密度为1、2、3、4、5头的红彩瑞猎蝽,分别与30头斜纹夜蛾3龄幼虫组合,置于直径为15 cm培养皿中,放入温度分别为30、33、36、39℃的恒温培养箱中,每个温度处理重复5次,24 h 后统计斜纹夜蛾幼虫死亡数量。
1.3 数据处理
对试验所得数据用 Excel 2010和 SPSS 26.0软件进行整理和单因素方差分析(One-Way ANOVA),应用 Duncan 氏新复极差法进行差异显著性检验。
捕食功能反应:第一步,根据猎物密度与被捕食量之间的 Logistic 回归方程确定捕食反应的功能反应类型[16],方法是多项式模型:
其中, N0为猎物初始量,Ne 为猎物被捕食的数量。 P0,P1,P2,P3是要估计的常数、一次、二次和三次方系数,用最大似然法对被食猎物与 N0比例数据进行拟合(SAS Institute Inc, CATMOD 程序),检测拟合式中有无显著正或负的线性系数。如果最终估得的参数 P1=0,表示捕食者的捕食量随着猎物密度增加而直线上升,属于 Holling I 型; P1>0且 P2<0,表示捕食者的捕食量随猎物密度呈“S ”波动,对应的功能反应是 Holling III 型;P1<0,表示捕食者的捕食量随着猎物密度增加而增加,之后处于一个平稳状态,则对应的功能反应是 II 型反应。 Holling II 型功能反应模型:
其中 Na 为猎物被捕食数量, a 为瞬时攻击率, Tr 为总试验时长(Tr=1 d),N 为猎物密度, Th 为捕食1头猎物所需要的时间。
第二步,用最小二乘法以及回归分析求出对应的模型方程,进而得出方程中包含的生物量。
搜寻效应使用方程 0 1 h a aT N S + = 进行拟合[17] ,其中 S 为搜寻效应, a 为红彩瑞猎蝽对斜纹夜蛾幼虫的瞬时攻击率,Th 为捕食1头斜纹夜蛾所需要的时间, N0为斜纹夜蛾密度。
自身密度干扰效应用 Hassell-Verley 模型E=QP-m 拟合[18] ,捕食作用率 Na E NP = ,式中 Na 为被捕食猎物数量、N 为猎物初始数量、 P 为捕食者初始密度、 Q 为搜寻常数、 m 为干扰系数,方程直线化为lnE=ln Q–mlnP。
2 结果
2.1 高温胁迫对红彩瑞猎蝽存活率、卵孵化率和羽化率的影响
红彩瑞猎蝽在高温胁迫下各虫态的存活率均随着温度升高逐渐降低(表1)。温度在27~30℃时,红彩瑞猎蝽各虫态存活率均为100%,温度达到33℃时,3龄若虫存活率降为95.23%,其他各虫态存活率为100%,当温度达到42℃时,各虫态中存活率最高的是雌成虫和5龄若虫(66.67%),3龄若虫的存活率最低,只有52.38%。
高温对红彩瑞猎蝽卵孵化率有明显影响,温度从27℃升高到33℃时,卵孵化率略有升高,当温度从33℃继续升高时,卵孵化率随着温度升高而逐渐降低(图1)。各温度处理中,以33℃时卵孵化率最高,达到97.88%,經42℃温度处理后,卵孵化率降低到45.89%,与30℃和33℃处理差异达显著水平(F=339.134,df=5,p <0.001)。
高温胁迫对红彩瑞猎蝽羽化率的影响结果见图2,温度为27~33℃时,红彩瑞猎蝽羽化率随温度升高而增大,在33℃时羽化率最高(95.24%),温度升高到36℃后,红彩瑞猎蝽羽化率开始出现下降,但变化不显著。温度升高到39℃时,红彩瑞猎蝽的羽化率相对30、33℃处理显著下降,当温度达到42℃时,红彩瑞猎蝽的羽化率进一步下降为57.14%,与30、33℃处理间差异达显著水平(F=3.733,df=5,p=0.004<0.01)。
2.2 高温胁迫对红彩瑞猎蝽捕食斜纹夜蛾的影响
不同虫态红彩瑞猎蝽在不同温度下对斜纹夜蛾3龄幼虫的捕食量如表2所示,在30~39℃范围内,不同虫态红彩瑞猎蝽对斜纹夜蛾3龄幼虫捕食量均随着猎蝽虫龄增大而增加,相同虫态红彩瑞猎蝽捕食量随着温度先增加然后降低,其中温度从30℃升高到33℃时,捕食量随温度增加而增加,温度从33℃升高到39 ℃时,捕食量随着温度升高而逐渐降低。同一虫态红彩瑞猎蝽在各温度条件下,对斜纹夜蛾3龄幼虫的捕食量随着猎物密度增加而增加,当猎物密度增加到40头时,捕食量增速放缓。
回归分析结果显示(表3),红彩瑞猎蝽不同虫态在不同温度下对斜纹夜蛾密度处理的一次方系数 P1均小于0。并且红彩瑞猎蝽对斜纹夜蛾3龄幼虫的捕食量均随着猎物密度的增加逐渐增加(图3),然后变成缓慢增加或不再增加,并趋于稳定,呈负密度制约关系。表明不同温度下红彩瑞猎蝽各虫态对斜纹夜蛾3龄幼虫的捕食功能类型均属于Holling Ⅱ型。
根据 Holling Ⅱ圆盘方程拟合不同温度条件下各虫态红彩瑞猎蝽捕食功能反应参数(表4)。在各参数中,瞬时攻击率 a 和捕食能效 a/Th 是衡量天敌昆虫对害虫捕食能力强弱的依据,从表4可知,红彩瑞猎蝽3龄若虫在30℃下捕食能力最强(a=0.88, a/Th=19.1304),之后随着温度升高,捕食能力逐渐减弱。4龄若虫、5龄若虫和成虫均在33℃下捕食能力最强,之后随着温度升高,各虫态红彩瑞猎蝽的捕食能力逐渐减弱。
2.3 高温条件下红彩瑞猎蝽对斜纹夜蛾的寻找效应
在30~39℃各温度条件下,红彩瑞猎蝽成虫对斜纹夜蛾3龄幼虫的寻找效应均与猎物密度呈负相关(图4),即随着猎物密度的增加,对猎物的搜寻效应逐渐减弱。在相同猎物密度下,雌成虫的寻找效应在33℃最高,在30℃与36℃时寻找效应相似,39℃时寻找效应最低。同一猎物密度下,红彩瑞猎蝽雄成虫在33℃下寻找效应最高,其次为30℃和36℃ , 39℃条件下寻找效应最低。
2.4 高温条件下自身密度对红彩瑞猎蝽捕食的干扰反应
在不同密度条件下,红彩瑞猎蝽雌成虫均在33℃条件下捕食量达到最高,39℃捕食量最低(表5)。在温度30~39℃范围内,随着红彩瑞猎蝽密度的增加,被捕食的斜纹夜蛾3龄幼虫总数量和分摊竞争强度随之增加,同时,捕食作用率 E 逐渐降低,干扰效应明显。
采用 Hassell-Varley 干扰模型对不同温度条件下红彩瑞猎蝽雌成虫受自身密度干扰的捕食作用进行拟合(表6),捕食作用率与红彩瑞猎蝽密度之间的模型拟合的 R2均大于0.9,表明模型拟合度较高,搜寻系数 Q 在33℃时达到最大,随后随着温度的升高而降低。相互干扰系数 m 则随温度的上升而升高,在39℃达到最大值,表明33~39℃温度范围内,红彩瑞猎蝽在33℃下的搜寻能力较强,且自身干扰系数未达到最高,是较适宜红彩瑞猎蝽雌成虫捕食生存的温度。
3 讨论
过高的温度会抑制昆虫的生长发育[19]。研究表明,阿里山潜蝇茧蜂(Fopiusarisanus)在38℃条件下,幼虫和蛹存活率均随处理时间增加而下降[20],但极短时高温处理对小菜蛾(Plutellaxylostella)生长发育没有显著影响[21]。广聚萤叶甲(Ophraellacommuna)随着温度从28℃升高到47℃时,寿命从88.0 d 缩短至51.3 d[22]。本研究结果表明,红彩瑞猎蝽3、4、5龄若虫和成虫在36℃高温下,依然能保持较高的存活率,5龄若虫羽化率和卵孵化率也能维持较高水平,但在39℃以后各虫态存活率下降明显,羽化率和卵孵化率也迅速降低,说明30~36℃仍在红彩瑞猎蝽适宜生存的温度范围内,但39~42℃高温条件对红彩瑞猎蝽生长发育有明显影响。对比在温度达36℃时,益蝽(Picromeruslewisi)卵孵化率为0,不能完成发育[23],经35℃高温处理后,双刺益蝽(Picromerusbidens)卵[24]和蠋蝽(Arma chinensis)卵[25]均不能孵化,红彩瑞猎蝽对高温耐受能力明显更强。
高温胁迫还可能导致天敌的捕食功能反应参数发生改变,掌握不同温度下的功能反应参数,有利于根据温度变化预测天敌控制效能[26]。红彩瑞猎蝽在27℃条件下对斜纹夜蛾幼虫的捕食反应属于 Holling II 型[9],温度升高至30~39℃时,红彩瑞猎蝽对斜纹夜蛾的捕食量、瞬时攻击率和捕食效能随着温度增加先升高而后逐渐减小,不同虫态红彩瑞猎蝽在不同高溫条件下,捕食功能反应依然属于 Holling II 型。红彩瑞猎蝽雌成虫在各温度条件下的搜寻效应均随着斜纹夜蛾密度增加而下降,这与蠋蝽对斜纹夜蛾幼虫的搜寻效应研究结论相同[27],随着温度的升高,红彩瑞猎蝽搜寻效应逐渐下降,这与刀角瓢虫(Serangiumjaponicum)[28]和海岛小花蝽(Oriusmaxidentex)[29]受高温胁迫后对猎物寻找效应的结果相似。双尾新小绥螨(Neoseiulusbicaudus)雌成螨[30]和加州新小绥螨[31]对猎物的瞬时攻击率和捕食效能在温度升高到30℃时开始下降,这与本文对红彩瑞猎蝽高温胁迫下捕食效能变化规律类似。红彩瑞猎蝽在33℃以后对斜纹夜蛾幼虫捕食量出现下降,与叉角厉蝽(Eocantheconafurcellata)若虫在温度高于32℃时对草地贪夜蛾日捕食量由上升转为下降趋势研究结果相同[32]。随着温度上升,红彩瑞猎蝽雌成虫自身密度干扰作用在39℃时最强,这与叉角厉蝽在37℃条件下种内竞争和自我干扰作用最强[31]的结论相似。高温胁迫也会导致某些天敌功能反应类型改变,烟盲蝽(Nesidiocoris tenuis )在温度从25℃升高到35℃时,对烟粉虱(Bemisiatabaci)的功能反应类型由 Holling II 型转变 Holling Ⅲ型[33],说明不同天敌种类的耐热能力不同,高温胁迫对其捕食作用的影响也不同。
4 结论
结果表明,30~36℃条件下短时高温胁迫不会对红彩瑞猎蝽生长发育和捕食作用产生不利影响,但温度达到42℃后,红彩瑞猎蝽生长发育和对斜纹夜蛾捕食能力受到明显抑制,说明该天敌对高温逆境具有一定耐受力。在夏季炎热天气将红彩瑞猎蝽应用于田间防治斜纹夜蛾时,释放时间应尽量挑选在清晨或傍晚,避免将红彩瑞猎蝽持久暴露在高温条件下而影响其防效。红彩瑞猎蝽对斜纹夜蛾实际防效还受到降雨、日照及其他生物因子的影响,有待进一步研究。
参考文献
[1] 黄霞.广西猎蝽科昆虫分类研究[D].桂林:广西师范大学,2007. HUANG X. Systematic study on Reduviidae(Heteroptera) from Guangxi[D]. Guilin: Guangxi normal university, 2007.
[2] 赵萍,袁继林.贵州真猎蝽亚科昆虫名录及区系分析[J].贵州农业科学,2011,39(7):99-102.
ZHAO P, YUAN J L. The insect list and faunal analysis of harpactorinae in Guizhou province[J]. Guizhou Agricultural Sciences, 2011, 39(7):99-102.
[3] AMBROSE D P. Bioecology of Rhinocorisfuscipes Fabr.(Reduviidae) a potential predator on insect pests[J]. Uttar Pradesh J.Zool, 1986, 6(1):36-39.
[4] SHAYARAJ K, SELVARAJ P. Life table characteristics of Rhynocorisfuscipes Fabricius in relation to sex ratios[J]. Ecology, Environment and Conservation, 2003, 9(2):115-119.
[5] AMBROSE D P, CLAVER M A. Functional and numerical responses of the reduviid predator Rhynocorisfuscipes F. (Het. Reduviidae) to ctton leafworm Spodoptera lituora F. (Lep., Noctuidae)[J]. J. Appl. Ent., 1997(121):331-336.
[6] AMBROSE D P, CLAVER M A. Functional and numerical responses of the reduviid predator, Rhynocorisfuscipes F.(Het. Reduviidae) to cotton leafworm Spodoptera litura F. (Lep. Noctuidae)[J]. Journal of Applied Entomology, 2010, 121(1):331-336.
[7] SHANKER C. Biology, Predatory Potential and Functional Response of Rhynocorisfuscipes (Fabricius)(Hemiptera: Reduviidae) on rice brown planthopper, Nilaparvatalugens (St?l.) (Homoptera: Delphacidae)[J]. J. Exp. Zool. India., 2018, 21(1):259-263.
[8] 邓海滨,吕永华,田明义,等.红彩真猎蝽对烟蚜的捕食功能反应及寻找效应[J].中国烟草学报,2015,21(5):74-78.
DENG H B, DENG H B, TIAN M Y, et al. Functional responses and searching efficiency of Harpactorfuscipes on predation of Myzuspersicae[J]. Acta TabacariaSinica, 2015, 21(5):74-78.
[9] 邓海滨,王珍,陈永明,等.红彩真猎蝽对斜纹夜蛾和烟青虫的捕食功能反应[J].广东农业科学,2012,39(13):107-109.
DENG H B, WANG Z, Chen Y M, et al. Predation of Harpactorfuscipes on Helicoverpaassulta and Spodoptera litura[J]. Guangdong Agricultural Sciences, 2012, 39(13):107-109.
[10] 邓海滨,吕永华,邱妙文,等.捕食性天敌红彩真猎蝽的生物学特性研究[J].中国烟草科学,2014,35(2):109-112.
DENG H B, LYU Y H, QIU M W, et al. Studies on the biological characteristics of predatory bug Harpactorfuscipes[J]. Chinese Tobacco Science, 2012, 39(13):107-109.
[11] 郭义,肖俊健,李敦松,等.低温饲养对红彩瑞猎蝽生长发育的影响[C]//病虫防护与生物安全——中国植物保护学会2021年学术年会论文集,2021:145.
GUO Y, XIAO J J, LI D S, et al. Effects of low temperature breeding on the growth and development of Rhynocorisfuscipes [C]//. Disease and insect protection and biological safety——Proceedings of the 2021 Annual Academic Conference of the Chinese Society for Plant Protection, 2021:145.
[12] 任麒麟.短時高温胁迫对草地贪夜蛾存活和繁殖的影响[D].贵阳:贵州大学,2021.
REN Q L. Effects of brief high-temperature stress on the survival and reproduction of fall armyworm (Spodoptera frugiperda)[D]. Guiyang: Guizhou University, 2021.
[13] 阮传清,SENGONCA C,刘芸,等.捕食性天敌黑纹透翅花蝽的生物学特性[J].中国生物防治,2008,24(3):199-204.
RUAN C Q, SENGONCA C, LIU Y, et al. Biological characteristics of Montandoniolamoraguesi (Hemiptera: Anthocoridae), a predator of Gynaikothripsficorum (Thysanoptera: Thripidae)[J]. Chinese Journal of Biological Control, 2008, 24(3):199-204.
[14] 李庆,崔琦,蒋春先,等.加州新小绥螨对朱砂叶螨的控制作用[J].植物保护学报,2014,41(3):257-262.
LI Q, CUI Q, JIANG C X, et al. Control efficacy of Chinese Neoseiuluscalifornicus (McGregor) population on Teranychuscinnabarinus (Boisduval)[J]. Acta PhytophylacicaSinica, 2014, 41(3):257-262.
[15] 罗优,肖峰,黄敏,等.温度对军配盲蝽生长发育及繁殖的影响[J].应用昆虫学报,2022,59(1):134-141.
LUO Y, XIAO F, HUNAG M, et al. Effects of temperature on the development and reproduction of Stethoconus japonicus (Hemiptera)[J]. Chinese Journal of Applied Entomology, 2022, 59(1):134-141.
[16] JULIANO S A. Nonlinear curve fitting: predation and functionalresponse curves[M]//SCHEINER S. Design and analysis of ecological experiments.2nd edition. New York: Chapman & Hall, 2001.
[17] 邓士群,张雯林,肖建飞,等.高效氯氟氰菊酯对东亚小花蝽捕食功能反应的影响[J].中国生物防治学报,2020,36(1):40-45.
DENG S Q, ZHANG W L, XIAO J F, et al. Effect of Lambda-cyhalothrinon on predatory function response of Oriussauteri[J]. Chinese Journal of Biological Control, 2020, 36 (1):40-45.
[18] HASSELL M P. A population model for the interaction betweenCyzenis albicans and Operophterabrumata at WythamBershire[J]. J. Anim. Ecol., 1969, 38:567-576.
[19] 刘安然.温度对昆虫繁殖力的影响及其机制初探[J].江西农业,2020(6):105,107.
LIU A R. Effect of temperature on insect fecundity and its mechanism[J]. Jiangxi Agriculture, 2020(6):105, 107.
[20] 郭俊杰.温度胁迫对阿里山潜蝇茧蜂的影响[D].福州:福建农林大学,2013.
GUO J J. Effects of temperature stress to Fopiusarisanus[D]. Fuzhou: Fujian Agriculture and Forestry University, 2013.
[21] 刘大成,杨中侠,张薇.极端高温和持续中等高温对小菜蛾耐热性的影响[J].激光生物学报,2019,28(4):374-379.
LIU D C, YANG Z X, ZHANG W. Stage-specific heat tolerance of Plutellaxylostella under extreme and moderate high temperatures[J]. Acta Laser Biology Sinica, 2019, 28(4):374-379.
[22] 张斯琴.广聚萤叶甲对高温变温的适应性[D].福州:福建师范大学,2018.
ZHANG S Q. The adaptability of Ophraellacommuna to high temperature change[D]. Fuzhou: Fujian Normal University, 2018.
[23] 唐藝婷.一种新天敌—益蝽的生物防治潜能研究[D].北京:中国农业科学院,2020.
TANG Y T. Study on potential of a novel natural enemy insect Picromeruslewisi Scott in biological control[D]. Beijing: Chinese Academy of Agricultural Sciences, 2020.
[24] 纪宇桐,薛传振,周磊,等.温度对捕食性蝽类昆虫生长发育影响的研究进展[C]//病虫防护与生物安全——中国植物保护学会2021年学术年会论文集,2021:144.
JI Y T, XUE C Z, ZHOU L, et al. Research progress on the effect of temperature on the growth anddevelopment of predacious bugs [C]//Pest protection and biosafety - Proceedings of the 2021 Annual Conference of the Chinese Society for Plant Protection, 2021:144.
[25] 李姿莹,杜钦祥,谢鹏飞,等.温度对蠋蝽 Arma chinensis 及其子代生物学特性的影响[J].中国植保导刊,2021,41(6):17-22.
LI Z Y, DU Q X, XIE P F, et al. Effects of temperature on the biological characteristics of Arma chinesis and its offspring[J]. China Plant Protection, 2021, 41(6):17-22.
[26] 胡长效,曹丹,徐万泰.不同温度和空间对龟纹瓢虫捕食褐软蚧的影响[J].河南农业科学,2019,48(12):73-78.
HU C X, CAO D, XU W T. Effect of different temperatures and spatial scales on predation of coccus hesperidum by Propylaea japonica[J]. Journal of Henan Agricultural Sciences, 2019, 48(12):73-78.
[27] 唐艺婷,王孟卿,李玉艳,等.蠋蝽对斜纹夜蛾幼虫的捕食作用[J].中国烟草科学,2020,41(1):62-66.
TANG Y T, WANG M Q, LI Y, et al. Predation of Arma chinensis on Spodoptera litura Larvae[J]. Chinese Tobacco Science, 2020, 41(1):62-66.
[28] YAO, F L, ZHENG Y, DING X L, et al. Effects of heat shock onsurvival and predation of an important whitefly predator, Serangium japonicum[J]. EntomologiaExperimentalis Et Applicata, 2019, 167(5):476-489.
[29] 李金磊,韩姗妮,周世豪,等.温度对海岛小花蝽5龄若虫捕食茶黄蓟马功能反应的影响[J].热带作物学报,2022,43(8):1671-1677.
LI J L, HAN S N, ZHOU S H, et al. Effects of temperature on the functional response of the fifth instar Oriusmaxidentex Ghauri Nymphs to Scirtothrips dorsalis Hood[J]. Chinese Journal of Tropical Crops, 2022, 43(8):1671-1677.
[30] 朱安迪,王映山, 方晨,等.不同溫度下双尾新小绥螨对西花蓟马的捕食功能反应[J].环境昆虫学报,2022, 44(2):430-439.
ZHU A D, WANG Y S, FANG C, et al. Functional response of predatory mite Neoseiulusbicaudus on western flower thrips, Frankliniella occidentalis at different temperatures[J]. Journal of Environmental Entomology, 2022, 44(2):430-439.
[31] 李庆,崔琦,蒋春先,等.加州新小绥螨对朱砂叶螨的控制作用[J].植物保护学报,2014,41(3):257-262.
LI Q, CUI Q, JIANG C X, et al. Control efficacy of Chinese Neoseiuluscalifornicus (McGregor) population on Tetranychuscinnabarinus (Boisduval)[J]. Acta PhytophylacicaSinica, 2014, 41(3):257-262.
[32] 张晓滢,彭之琦,陆永跃,等.不同温度条件下叉角厉蝽对草地贪夜蛾幼虫的捕食作用[J].环境昆虫学报,2022,44(2):273-280.
ZHANG X Y, PENG Z Q, LU Y Y, et al. Functional response of Eocantheconafurcellate (Wolff) on Spodoptera frugiperda (Smith) larvae at different temperatures[J]. Journal of Environmental Entomology, 2022, 44(2):273-280.
[33] ZIAEI MM A, SAMIH M A, Namvar P, et al.Temperature-dependent functional response of Nesidiocoris tenuis (Hemiptera: Miridae) to different densities of pupae of cotton whitefly, Bemisiatabaci (Hemiptera: Aleyrodidae)[J]. European Journal of Entomology, 2017, 114:325-331.
猜你喜欢
建材发展导向(2021年11期)2021-07-28 06:57:22
当代水产(2020年10期)2020-03-17 07:02:48
当代水产(2019年8期)2019-10-12 08:57:26
当代水产(2019年7期)2019-09-03 01:01:56
天津农业科学(2016年3期)2016-03-12 14:55:12
江苏农业科学(2015年5期)2015-10-20 00:48:01
江苏农业科学(2015年8期)2015-09-10 15:30:55
安徽农学通报(2015年4期)2015-07-30 23:59:23
江苏农业科学(2015年1期)2015-04-17 23:57:37
天津农业科学(2015年1期)2015-01-12 22:07:26
