
兴隆山自然保护区种子植物的垂直分布格局研究
2023-08-14 06:26:04安永梅杜维波周晓雷张晓玮武利玉
西北植物学报 2023年6期
安永梅,杜维波,周晓雷,张晓玮,武利玉
(甘肃农业大学 林学院,兰州 730000)
植物多样性随环境梯度的变化规律一直是植物生态学研究的核心问题之一[1]。山地生态系统因具有浓缩的环境梯度和生境异质性高的特点,为各植物类群提供了天然的生存场所和分化条件,成为了植物多样性的富集地[2],也是研究植物多样性的热点区域,一直备受各类学者的关注[3]。山地生态系统由于海拔升高使得气候条件、植被类型、土壤等特征发生了相应的变化,从而形成了植被垂直带,对山地植物多样性沿植被垂直带海拔升高的分布规律进行研究,可以揭示植物长期演替下的生态适应性。国内外开展的有关山地植物多样性随海拔梯度变化的研究表明,山地植物多样性垂直分布格局存在单峰曲线、先降后升、单调递增或递减和无明显变化5种分布模式[4-8],这可能与山体所处的区域位置、气候条件、山体高度和生活型等方面的差异相关。已有研究表明,在热带地区山地植物多样性随着海拔升高以单调递减变化趋势为主,亚热带地区主要呈单峰曲线的变化趋势,但针对温带半干旱区研究较少变化趋势尚不明确[9-13]。
兴隆山自然保护区地处中国西北部,植被呈垂直地带性分布,目前,关于该保护区的研究主要集中在植物保护利用[14]、资源调查[15]和土壤理化性质[16]等方面,而针对植物区系成分和系统发育结构沿植被垂直带的变化规律尚不明晰。因此研究半干旱区山地植物垂直分布格局,对阐明植物多样性与环境间的关系具有重要意义。基于此,本研究运用植物区系地理学和系统发育学的相关理论和方法,对保护区各植被垂直带种子植物生活型、区系地理成分的变化趋势进行分析,进一步揭示植物多样性沿山地环境变化的垂直分布规律,可以为山地植物多样性保护、植物资源合理开发利用和引种驯化等方面提供科学的理论依据[17],利用物种间的系统发育关系探明该保护区系统发育结构,对了解植物起源、分化、物种适应性及群落构建机制具有重要意义。
1 研究区概况
兴隆山国家级自然保护区(103°50′-104°10′E,35°38′-35°58′N)位于甘肃省兰州市榆中县境内,在植物地理区划上是黄土高原、青藏高原和蒙新高原的交汇地带,主要是由马衔山、兴隆山及沟谷丘陵地组成,其山脉属于祁连山东延余脉,总面积为33 301 hm2,海拔1 800~3 670 m[18]。保护区内气候特征表现为大陆性温带半湿润气候,年平均气温在3~7 ℃,年均降水量在600 mm以上,平均无霜期为103 d[15]。随着海拔的升高,保护区内土壤和植被均呈垂直地带性分布,马衔山的褶穹和兴隆山褶凹的构造单元可视为同一巨大山体研究其植被垂直带[19],各垂直带间存在着一定的过渡性。
2 材料和方法
2.1 植物名录的构建
研究以《甘肃兴隆山国家级自然保护区资源本底调查研究》的种子植物名录为基础[19],搜集并查阅文献、书籍和标本记录[20-24]对保护区植物遗漏种进行增补。在此基础上,结合保护区的地形、生境和植被群落类型等特征,通过样线法进行野外实地调查,记录所观察到的植物种类、生境特征、海拔高度等信息,采用《中国生物物种名录》2022版(http://sp2000.org.cn)对植物科、属、种名及拉丁名进行核对校正,对异名、重复、无海拔及栽培种数据进行修正和剔除。通过实地调查并参考文献[9]将保护区划分为6个植被垂直带(表 1),并根据植物的海拔分布范围整理出6个植被垂直带的植物名录,用于后续分析。
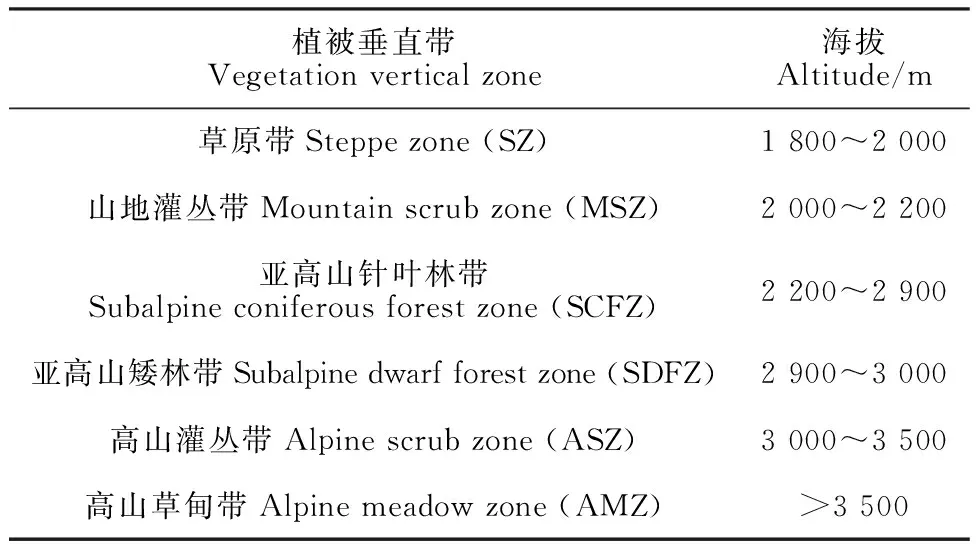
表1 兴隆山自然保护区的植被垂直带Table 1 Vertical zone of vegetation in Xinglong Mountain Nature Reserve
2.2 群落相似性
Jaccard相似性系数表征植被垂直带间物种更替的速率,其值越小,则表示植被带间共有物种越少,物种更替速率越大[25],其计算公式为:
(1)
式中:c为两植被带共有物种数;a和b分别为两植被带的物种总数,当q在0~0.25间表示极不相似性,在0.25~0.5间表示中等不相似,在0.5~0.75间表示中等相似,在0.75~1间表示极相似[26]。
2.3 植物区系成分
根据《中国维管植物科属志》[27]对中国种子植物属的区系分布类型界定的划分方法,将兴隆山自然保护区种子植物属的区系成分划分为世界广布型(T1)、中国特有成分(T15)、热带区系成分(Tr)和温带区系成分(Tw)4类进行统计分析[28]。
2.4 系统发育指数的选取
研究利用R软件中的V. PhyloMaker程序包[29],以兴隆山自然保护区种子植物的拉丁名为基础构建了带有枝长的系统发育树(图1)。将系统发育树与植被垂直带物种数据相匹配计算系统发育多样性 (phylogenetic distance,PD)和净种间亲缘关系指数(net relatedness index,NRI),PD值越大表明该植被带在保护区系统发育多样性的比重越高[30],NRI侧重从整体出发反映研究群落物种所形成的系统发育结构[31]。计算各植被垂直带所有物种对的平均谱系距离(mean phylogenetic distance, MPD),保持丰富度与发生频率不变,运用独立置换的方法将各植被带内的植物在物种库中随机抽取999次,从而获得各植被带植物在随机零模型下的MPD分布,最后利用随机分布结果将观察值标准化,从而得出NRI[32-33],其计算公式为:
图1 兴隆山自然保护区种子植物系统发育树Fig.1 Phylogenetic tree of seed plants in Xinglong Mountain Nature Reserve
(2)
式中:MPD为观察值;MPD′为随机零模型期望值;SD(MPD′)为标准偏差。NRI是MPD标准化效应指数的负值,Mpd.obs.p(P-value of observed mpd vs. null communities)是观察值与零模型期望值相比的显著度,若NRI >0且Mpd.obs.p <0.05时,则表明该植被带的系统发育结构为聚集显著;若NRI <0且Mpd.obs.p >0.95时,则表明该植被带系统发育结构发散显著;NRI = 0则表明系统发育结构呈随机分布。NRI的绝对值要小于1.96,否则被认为系统发育结构显著偏离零模型[34-35],计算过程中不使用多度加权,以上计算均在R软件的Picante包中进行。
3 结果与分析
3.1 各植被垂直带种子植物的数量特征和相似性
3.1.1 各植被垂直带种子植物数量分布特征如表2所示,按最新的分类系统划分,兴隆山自然保护区种子植物隶属于87科387属889种,构成各植被垂直带植物物种数量分别为:草原带40科128属200种,81科304属661种,亚高山针叶林带81科304属661种,亚高山矮林带37科83属137种,高山灌丛带31科75属135种,高山草甸带26科57属119种。植物科、属、种丰富度及其占比随着植被带海拔升高呈单峰分布格局,均在亚高山针叶林带达到峰值。
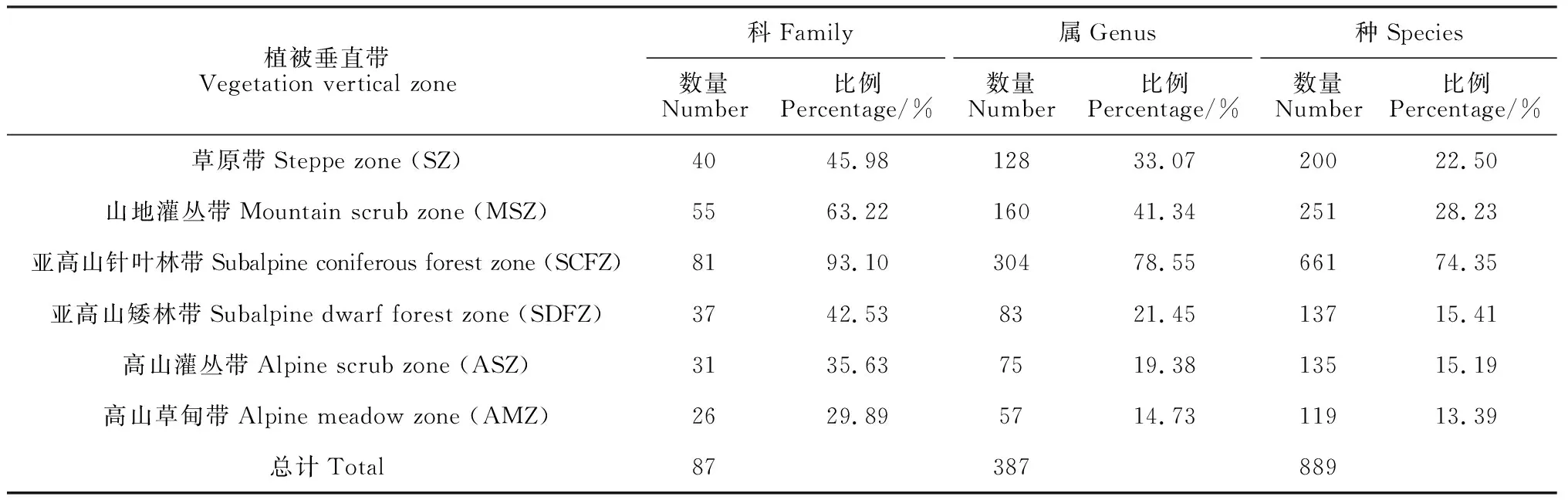
表2 兴隆山自然保护区各植被垂直带种子植物数量统计Table 2 Statistics on the number of seed plants in each vegetation vertical zone of Xinglong Mountain Nature Reserve
3.1.2 各植被垂直带的群落相似性从两植被垂直带间共有植物物种数量可以得出(表3),山地灌丛带和亚高山针叶林带间共有物种数最高,达181种,其次是草原带和亚高山针叶林带间,共有种数为109种,草原带和高山草甸带间共有种数最少,仅有2种。通过Jaccard相似性系数分析表明,亚高山矮林带和高山灌丛带间的相似性系数最大(0.402),其次是高山灌丛带和高山草甸带间(0.373),均属于中等不相似;其他各植被带间相似系数均在0~0.25间,属于极不相似。此外,各植被垂直带间的相似性随海拔高度差的增加而逐渐降低,海拔高度越接近的植被带间相似性越高。

表3 兴隆山自然保护区各植被垂直带间的相似性系数Table 3 Similarity coefficients among the vertical zones of vegetation in Xinglong Mountain Nature Reserve
3.2 各植被垂直带的生活型特征
如图 2所示,木本植物沿植被垂直带海拔升高呈先升后降的变化格局,乔、灌木峰值略有差异。灌木所占比例在山地灌丛带达到峰值,为19.12%;乔木所占比例呈“∧”字形,在亚高山针叶林带达到峰值,为4.39%,在草原带、高山灌丛带和高山草甸带均没有分布;藤本所占比例最低,在草原带至亚高山针叶林带变化较为平缓,随后下降至0%。在各植被垂直带中草本植物所占比例始终高于木本植物,沿植被带自下而上依次为90.50%、78.09%、79.12%、81.75%、84.44%和89.92%,呈先降后升的变化趋势。一年生草本所占比例随植被带海拔升高呈单调递减的变化趋势,从草原带的25%降低至高山草甸带的7.56%;多年生草本所占比例呈先降后升的变化趋势,这是由于在高海拔区域的高山草甸带中有大量多年生草本植物存在。上述结果表明,保护区内植物生活型所占比例的垂直变化趋势与植被垂直带类型变化相对应。
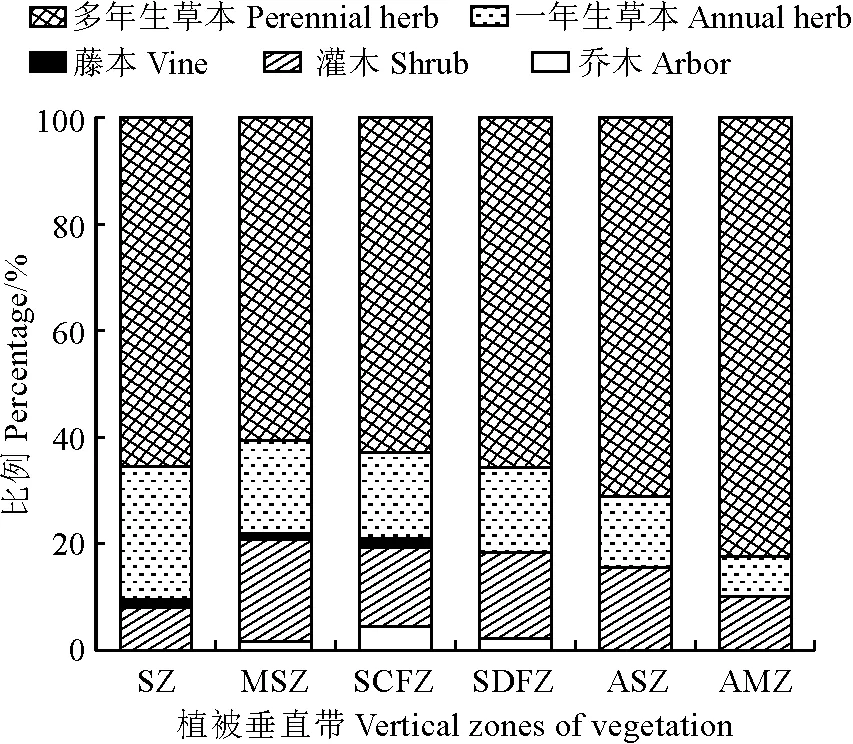
图2 各植被垂直带不同生活型所占比例Fig.2 Proportion of different life types in each vegetation vertical zone
3.3 各植被垂直带的植物区系成分
由图 3可知,在属水平上,兴隆山自然保护区种子植物温带成分(Tw)在各植被垂直带中所占比例不低于70%,远高于其他区系类型,且随植被带海拔升高整体呈逐渐增加的变化趋势,仅在高山草甸带出现小段下降;而热带成分(Tr)所占比例呈逐渐下降的变化趋势,仅在高山草甸带有略微上涨,各植被带占比低于8%,这表明该保护区植物主要呈温带性质。
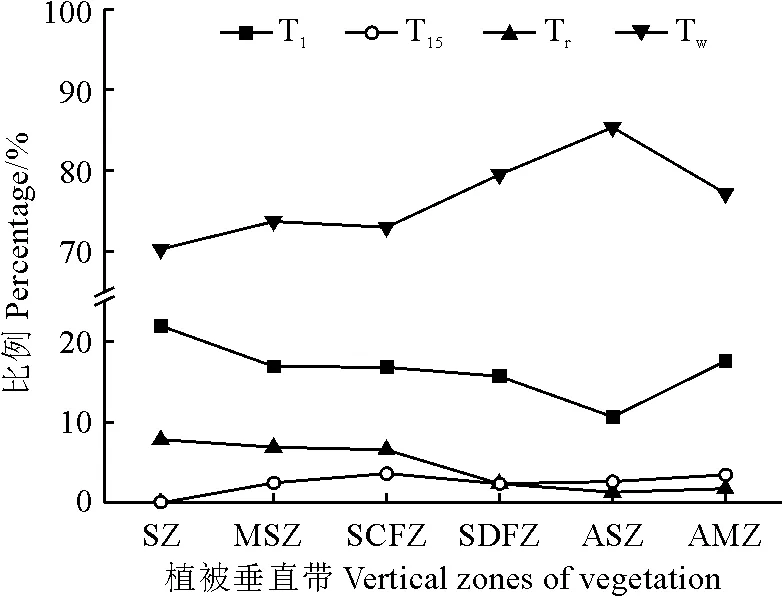
T1表示世界广布型,T15表示中国特有成分,Tr表示热带区系成分,Tw表示温带区系成分。图3 属的区系地理成分沿植被带的垂直变化趋势T1 is the world-wide type, T15 is the Chinese endemic component, Tr is the tropical component, and Tw is the temperate component.Fig.3 Vertical trends in the zonal geographic composition of genera along vegetation zones
世界广布型(T1)所占比例随着植被带海拔升高呈先降低后升高的变化趋势,在草原带达到峰值,占比为21.88%。中国特有成分(T15)所占比例总体最低,且变化趋势较为平缓,仅在2%~4%范围内波动。
3.4 系统发育多样性及其结构
由表 4可知,兴隆山自然保护区6个植被垂直带中的系统发育多样性指数范围在4 577.89~17 543.37之间,随着植物丰富度的增加,PD值也随之增大,通过线性回归模型可以得出保护区种子植物PD与Ntaxa呈显著正相关性(R2= 0.985,P<0.001,y= 22.952x+2 621.3)。由于草原带和山地灌丛带NRI绝对值大于1.96,表明两植被带的系统发育结构显著偏离零模型。亚高山针叶林带NRI = -1.017 <0,且Mpd.obs.p = 0.965 > 0.95,表明该植被带与零群落相比,系统发育结构呈显著发散;亚高山矮林带、高山灌丛带和高山草甸带的NRI分别为1.832、1.617和1.587,均大于0,且Mpd.obs.p分别为0.008、0.028和0.021,均小于0.05,则表明高海拔植被带(>2 900 m)的系统发育结构呈显著聚集。
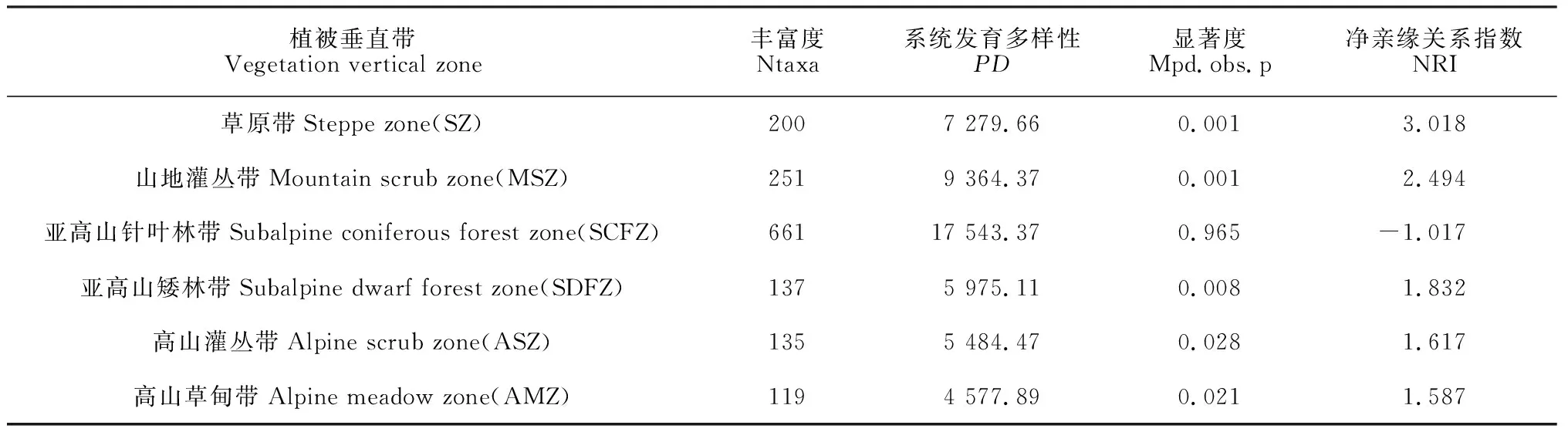
表4 各植被垂直带系统发育指数Table 4 Phylogenetic index of each vegetation vertical zone
4 讨 论
4.1 植物丰富度和相似性的垂直分布格局
兴隆山自然保护区种子植物丰富度沿植被垂直带海拔升高呈单峰分布格局,造成的原因:一是植被垂直带海拔梯度划分的不同,面积大小存在差异[36];二可能是由于低海拔区域(1 800~2 200 m)干扰因素强烈,此外,虽热量充足但水分限制作用较强,相反高海拔区域(>2 900 m)热量是主要的限制因子,相比较而言,中低海拔区域(2 200~2 900 m)拥有着较好的水热生境组合,因此单峰分布格局符合水分-能量假说[37-38];三是符合“中间膨胀理论”,各植被垂直带间存在着不同重叠的植物物种和分布范围,高低海拔间由于海拔跨度大,植物重叠范围较小,所以导致在亚高山针叶林带丰富度最高[39-40]。综上所述,保护区植物丰富度呈单峰曲线是由于面积大小、干扰因素、水热条件限制和中间膨胀效应综合作用下所导致的结果。
本研究表明,在各植被垂直带间Jaccard相似性系数呈中等不相似和极不相似水平,这与陈思艺等[41]对星斗山森林群落的研究结果一致,造成的原因可能是各植被带间坡度、坡向、水热条件和土壤类型等环境条件的差异,在一定程度上对植物物种间的交流产生了阻隔作用,因此导致各植被带间群落相似性较小[41]。海拔高度越靠近的植被带间相似性系数越大,可能是由于海拔高度差影响植物本身的扩散和定居,从而引起各植被带的植物物种组成和群落结构发生适应环境的改变[42]。
4.2 不同生活型的垂直分布格局
本研究表明,不同生活型植物在各植被垂直带的变化趋势和峰值有所不同,这是由于各生活型植物对不同海拔生境条件的敏感性不同[43]。灌木峰值低于乔木,这种不同生活型峰值在不同海拔的现象,在其他山地垂直分布格局研究中也有存在[44],乔木呈“∧”字形原因可能是低海拔水分限制和高海拔热量限制导致。一年生草本随植被带海拔升高呈单调递减的分布格局,这与刘彬等[45]对新疆天山南坡种子植物的研究结果相符。多年生草本呈先降后升的变化趋势,造成的原因可能是随着海拔升高水热成分逐渐降低,绝大多数一年生草本不能适应寒冷、低温的生境,而多年生草本可以通过形成轴根、密生及根茎等复杂的生存策略来适应高寒、生长季节缩短的气候特点,如西藏点地梅(Androsacemariae)、珠芽蓼(Bistortavivipara)等植物。
4.3 植物区系地理成分的垂直分布格局
植物分布区类型中属较科更能具体表明植物在历史进化过程中的分异状况和地理特征[46]。本研究表明,在属级水平上温带成分所占比例占绝对优势,符合保护区温带半湿润的气候特征,并沿植被带海拔升高整体呈上升的变化趋势,这与冯建孟等[47]研究结果一致,可能是由于温带区系成分植物具有耐低温、适宜在高海拔区域生存的适应性。王俊伟等[48]对藏南布丹拉山南坡植物的研究得出,热带成分所占比例随海拔升高呈下降变化趋势,这与本研究结果相符,表明随着海拔升高热带成分受温度-热量的影响逐渐增加,热带成分必然减少;热带成分在山顶的高山草甸带有略微的上涨,这可能与高海拔极端恶劣的环境导致植物分化强烈的结果有关[48]。综上所述,植物区系地理成分的垂直分布格局主要受当地气候特征和水热条件变化的共同影响。
4.4 系统发育结构的垂直分布格局
本研究表明,在中低海拔的亚高山针叶林带呈显著的发散结构,在高海拔区域(>2 900 m)的3个植被带中均呈显著的聚集结构,说明非随机格局在群落构建过程中起主要作用,中性理论预测系统发育结构的随机格局在兴隆山自然保护区并不适用,这与大多数研究[49]一致,为探讨物种共存和植物的生态适应性提供了新的角度。生境过渡和竞争排斥是生态位理论中主要的两种作用机制,前者作用使物种适应性相似,表现为系统发育结构聚集,而后者使亲缘关系近的物种出现排斥现象不能共存,致使系统发育结构发散[50]。兴隆山自然保护区在中低海拔表现出系统发育结构发散,可能是由于该区域存在以青杄为主的建群树种,裸子和被子植物的亲缘关系较远,并且该区域拥有着充足的自然资源和适宜的环境条件,物种间为竞争同一生存资源会使植物生态位逐渐分化,最终导致种间亲缘关系相对较远的植物共存[51];而高海拔植被带系统发育结构聚集,表明生境过滤在群落构建中起决定性作用,由于高海拔资源条件有限,在强烈的生境异质性和生理胁迫压力驱使下导致该区域亲缘关系近的物种聚集[52]。本研究从中低海拔至高海拔,系统发育结构由发散转变成聚集,这与Kembel等[53]的研究结果类似,表明植物物种间竞争作用减弱的同时伴随着环境压力的增强[54]。
5 结 论
通过对兴隆山自然保护区种子植物的研究表明:按最新的分类系统划分,保护区种子植物隶属于87科387属889种,植物科、属、种丰富度均呈单峰分布格局;各植被带间群落相似性系数呈中等不相似和极不相似水平,海拔越靠近的植被带间相似性系数越高;不同生活型植物所占比例的垂直变化趋势存在差异,木本植物所占比例沿植被垂直带海拔升高呈先升后降的变化趋势,而草本植物呈相反的变化格局。在属级水平上,温带成分在各植被垂直带占绝对优势,所占比例随着植被垂直带海拔升高整体呈上升的变化趋势,而热带成分所占比例呈相反的变化趋势。各植被垂直带中系统发育结构呈非随机分布,沿植被带海拔升高由发散转变为聚集,这对了解兴隆山自然保护区物种共存和群落构建机制具有重要意义。本研究尚未与土壤、水热条件和微生物等具体环境因子相结合,有待通过研究深入探讨种子植物多样性垂直分布格局的形成机制。
猜你喜欢
数学物理学报(2022年5期)2022-10-09 08:56:44
散文诗(2021年22期)2022-01-12 06:14:04
河北画报(2020年8期)2020-10-27 02:54:20
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
甘肃林业(2016年2期)2016-11-07 08:56:10
北方音乐(2016年12期)2016-08-23 03:20:04
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:51
甘肃林业科技(2016年1期)2016-03-11 17:49:05
甘肃林业科技(2016年2期)2016-03-10 14:54:25
