
晚白海棠×华月苹果杂交种实生苗若干性状的遗传分析
2023-08-14 06:34:39李娇安秀红邵建柱李武兴田义
江苏农业科学 2023年13期
李娇 安秀红 邵建柱 李武兴 田义


摘要:以苹果品种晚白海棠为母本、栽培品种华月为父本构建的杂交群体为试材,通过对杂交种实生苗的果实、叶片等若干性状的遗传变异和遗传倾向进行分析,鉴定苹果若干性状的遗传规律,以期为杂交育种亲本的选配提供参考。结果表明,晚白海棠×华月苹果杂交种实生苗存在丰富的遗传多样性。杂交种实生苗的单果质量、果实横径、果实纵径、果形指数、叶片长度、叶片宽度、可溶性固形物含量等性状均呈正态分布,單果质量、果梗粗、果实横径、果实纵径间呈极显著正相关。杂交群体单果质量的变异系数为41.20%,杂交群体的单果质量更倾向于小果型母本晚白海棠,低于低值亲本后代占比超过84%,表现出性状退化的趋势。杂交种实生苗果实果形主要以双亲果形为主,果形指数的变异系数只有6.74%,有向圆形果变异的趋势。果形指数、色差指数、可溶性固形物等重要性状的遗传传递力均达到90%以上。可溶性固形物低于低值亲本比率为44.09%,可滴定酸低于低值亲本比率达65.59%,但在杂交种实生苗中也出现了一些性状表型优于高值亲本的株系。
关键词:苹果;杂交种实生苗;性状;遗传倾向
中图分类号:S661.03 文献标志码:A
文章编号:1002-1302(2023)13-0169-06
中国是世界苹果生产大国,但主栽品种中具有自主知识产权品种的占有率却很低[1]。为了保障我国苹果产业可持续健康发展,满足人们对苹果品种多样化的需求,必须尽快培育出多种类型的苹果优质新品种[2]。然而,由于苹果树童期长、树体高大、自交不亲和等因素,导致苹果育种周期长、成本高、难度大。遗传规律研究是苹果育种的重要基础,对缩短苹果育种周期和提高苹果育种效率具有重要意义。迄今为止,关于杂交种实生苗若干性状的遗传分析已有许多报道。闫忠业等通过对红玉×金冠苹果杂交种实生苗进行遗传规律研究发现,果糖、苹果酸等是数量性状并由微效基因控制;草酸、琥珀酸等可能是质量性状,由2对主效基因控制且受到微效基因的影响[3]。赵亚楠等通过对苹果梨×八月红杂交种实生苗6个性状的遗传倾向进行研究发现,果实大小、果形指数、果心大小、可溶性固形物含量4个性状符合正态分布或者偏正态分布,果肉硬度、可滴定酸含量2个性状为主基因和多基因模型共同作用的多峰分布[4]。秦冠×富士苹果[5]、京白梨×鸭梨[6]、宁海白×大房枇杷[7]、黑比诺×马瑟兰葡萄[8]等不同种类果树上的遗传分析均表明,果实大小是受多基因控制的数量性状。
刘振中等对富士×粉红女士苹果杂交F1代的遗传变异分析发现,果形为趋中变异,果实大小、可溶性固形物等为退化的负向趋势[9]。韩婷婷等对金冠×红勋1号苹果杂交F1代群体研究发现,糖酸比以及花青苷含量表现出超亲优势[10]。陈学森等对新疆红肉苹果与苹果品种杂交种F1代群体进行研究,发现后代结果年龄早,果皮和果肉总酚遗传能力强[11]。王家珍等对南果×红巴梨杂交后代进行遗传倾向分析发现,叶缘形态、叶尖形状和母本一样,叶片大小介于亲本之间,果形大多数为圆形,果实质量有向小果退化的趋势,果实底色为母性遗传[12]。刘有春等对越橘半同胞系杂交后代研究表明,糖酸含量存在衰退变异的趋势[13]。吴斌等通过对百香果杂交子代进行遗传分析发现,总酸、可溶性固形物含量受母本影响大[14]。刘政海等通过对威代尔×霞多丽葡萄杂交后代进行遗传倾向分析发现,酸糖含量呈现较广的分离,趋向于低糖和高酸亲本[15]。王勇等对红地球葡萄杂交后代进行遗传规律研究发现,脆肉和果粒性状呈减弱趋势[16]。王燕等对中国樱桃、龙眼、红阳猕猴桃、山梨与中华猕猴桃种间杂交后代进行遗传分析发现,果实大小为趋小,呈退化的负向趋势,可溶性固形物表现为超亲遗传[17-20]。
本研究以苹果野生资源晚白海棠为母本、栽培品种华月为父本构建的遗传群体为研究对象,通过对杂交种实生苗群体的果实以及叶片等若干性状进行调查评价,探讨相关性状的遗传规律,旨在为重要性状候选基因的挖掘提供坚实的基础,并筛选出优异新种质。
1 材料与方法
1.1 试验材料
试验材料为辽宁兴城中国农业科学院果树研究所苹果品种晚白海棠×华月的F1代杂交种实生苗276株,于2016年播种,2018年定植于实生苗圃,株行距1.5 m × 4 m。亲本和杂交种实生苗均于田间自然生长,无疏花疏果、套袋等处理措施。
晚白海棠:平均单果质量为52 g;果实形状为近圆形或长圆形;果皮颜色为黄白,阳面有少许红晕,8月上旬果实成熟。
华月:以华富为父本、金冠为母本杂交选育而成,平均单果质量为216 g;果实形状为圆柱形;果皮颜色为黄绿色,阳面带红晕[21]。10月中下旬果实成熟[22]。
1.2 试验方法
于2019—2021年对晚白海棠×华月杂交种实生苗的重要性状进行调查。每个单株树冠四周选取20个典型果,用电子天平称单果质量;用游标卡尺测果梗长和果梗粗,果实横纵径、叶片长度和宽度等。可溶性固形物用ATC手持折光仪测定,可滴定酸用酸度计进行测定。调查标准严格按照王昆等的《苹果种质资源描述规范和数据标准》 [23]进行规范调查。另外用HP-200型精密色差仪测定果实色差。用标准白板校准L*值(值越大亮度越高)、a*值(正值越大红色越深,负值越小绿色越深)、b*值(正值越大黄色越深,负值越小蓝色越深)、C*值(色饱和度,值越大颜色彩度越纯)和H°值(色调值,180°表示纯绿色,90°表示黄色,0°表示纯红色)。
1.3 数据分析
按照王昆等的《苹果种质资源描述规范和数据标准》 [23]对描述型性状进行赋值标准化处理。统计数据用Excel、SPSS软件进行处理和分析[24]。
遗传传递力:Ta=x/F×100%,x为子代均值,F为亲中值。
变异系数:CV=σ/x×100%,x为子代均值,σ为标准差。
2 结果与分析
2.1 杂交种实生苗描述型性状的多样性分析
苹果是高度杂合的二倍体,杂交后代性状分离明显。杂交种实生苗在果实和叶片上均出现了明显的分离。杂交种实生苗不同单株主要描述型性状的分布频率和遗传多样性统计见表1。梗洼深度主要类型为深,分布频率为61.70%;萼洼深度主要类型为浅,分布频率为67.02%;果点大小主要类型为小,分布频率为54.26%;果点状态主要为平、凸2种,分布频率分别為52.13%、47.87%;果实大部分无棱起,分布频率为77.42%。大部分叶片性状表型叶缘为锐锯齿,分布频率为66.86%,叶尖为渐尖,分布频率为61.54%,叶面为抱合状,分布频率为61.54%。梗洼广度、果点密度的多样性指数较高,分别为1.10、1.09;萼洼广度的多样性指数最低,仅为0.34,说明萼洼广度为低变异性状,遗传较为稳定。
2.2 杂交种实生苗数量性状的遗传变异规律
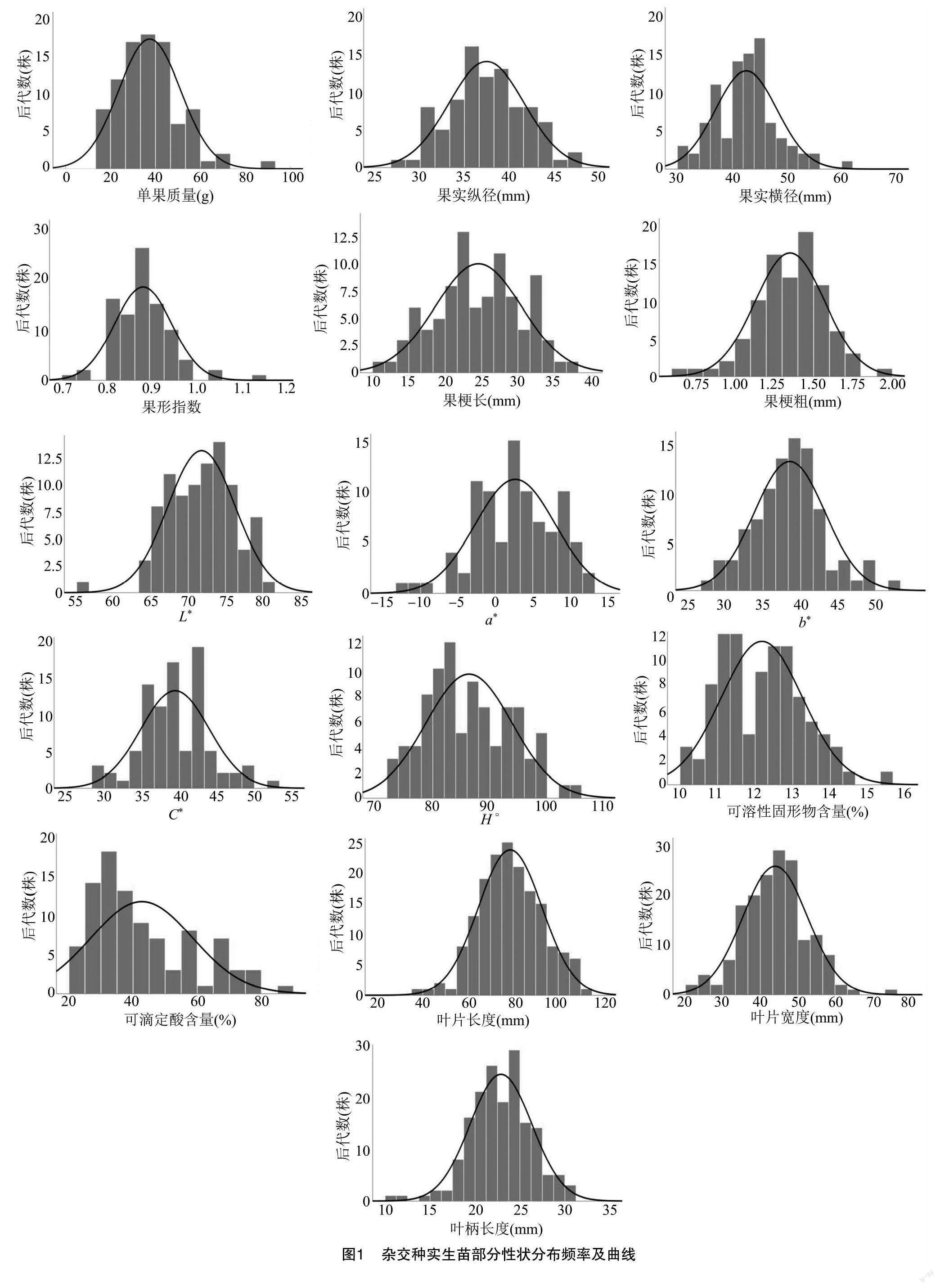
对晚白海棠×华月苹果杂交种实生苗群体各数值型性状进行测定及数据分析(图1)显示:单果质量、果实横纵径、果形指数、叶片长度、叶片宽度、色差、可溶性固形物等重要性状分布基本符合正态分布,说明这些性状是数量性状,受多个基因控制。单果质量中值左侧的频率明显大于右侧,说明杂交种实生苗果实有变小的趋势;果实横、纵径平均值附近分布较为集中;色差b*、叶片长度、叶片宽度、叶柄长度分布较为集中。另外杂交种实生苗出现了大量低酸个体,为偏离正态分布,暗示可滴定酸可能存在主效基因。分别对晚白海棠×华月苹果杂交种实生苗F1群体的16个数量性状进行测定。由表2可知,杂交种实生苗数值型性状指标如下:单果质量为13.40~93.17 g,果实纵径为22.65 ~47.59 mm,果实横径为24.26 ~60.00 mm,果形指数为0.72~1.06,果梗长为10.09~35.48 mm,果梗粗为0.65~1.97 mm,可溶性固形物含量为 10.18%~14.34%,叶片长度为42.11 ~108.22 mm,叶片宽度为21.13 ~60.77 mm,叶柄长度为14.39~30.51 mm。杂交种实生苗的主要数量性状变异系数为6.74%~ 41.20%,单果质量变异系数(41.20%)>可滴定酸含量(39.02%)>果梗长(33.62%)>叶柄长度(32.06%)>果梗粗(27.48%)>叶片宽度(21.47%)>叶片长度(20.12%)>可溶性固形物含量(8.77%)>果形指数(6.74%)。综合比较杂交群体变异系数发现,16个数量性状中有8个变异系数大于20%,具有丰富的多样性。果形指数平均值为0.89,变异系数最小,表明晚白海棠×华月苹果杂交种实生苗的果形变异程度小,分离程度低。单果质量的变异系数最大,表明杂交种实生苗在单果质量性状上分离明显。
由表2可知,单果质量平均值为35.82 g,明显低于中亲值134.00 g,低于低亲比率高达84.44%;果实纵径平均值为36.93 mm,显著低于中亲值51.22 mm,低于低亲比率为48.89%;果实横径平均值为41.79 mm,低于中亲值60.01 mm,低于低亲比率高达80.00%;果梗长平均值为24.84 mm,低于中亲值28.06 mm,低于低亲比率为13.33%;果梗粗平均值为1.31 mm,低于中亲值2.14 mm,低于低亲比率达66.67%;可滴定酸含量平均值为0.41%,低于中亲值0.66%,低于低亲比率达65.59%。因此,与其父本华月相比,上述杂交种实生苗果实相关的重要性状更倾向于母本晚白海棠,多数量性状均表现出衰退变异。果形指数超高亲比率为54.44%,低于低亲比率为14.44%,色差a*、b*、C*的超高亲率分别为45.56%、62.22%、65.56%,表明杂交群体有明显的超亲遗传趋势,杂交种实生苗易出现果形指数、色差a*、b*、C*高于高亲的单株。色差H°超低亲率为51.11%,远远大于超高亲比率值(13.33%)。可溶性固形物含量低于低亲比率为44.09%,可滴定酸含量超低亲率为65.59%。叶片长度、宽度的超低亲率分别为80.36%、50.00%,由此可知,杂交种实生苗易出现低酸、叶片小等低于低亲的单株。色差的遗传传递力L*为95.65%,b*为104.47%,C*为106.48%,果形指数的遗传递力为104.71%,叶柄长度的遗传传递力为89.12%,较高的遗传传递力表明这些性状均能稳定地从亲本遗传给后代。
2.3 杂交种实生苗部分数量性状的相关性分析
表3中晚白海棠×华月苹果杂交种实生苗10个表型性状的相关性分析表明,单果质量与果梗粗度、果实横径、果实纵径的相关系数分别为0.453、0.955、0.867,呈极显著正相关,说明果实横纵径越大,果梗越粗,果实单果质量越大。叶片长度与叶片宽度相关系数为0.792,呈极显著正相关,而叶片长度与与果实单果质量呈显著正相关,暗示在本杂交组合内,通过对实生苗叶片长度的测定,即可对果实的大小进行简单的预测。
3 讨论与结论
我国苹果育种起步较晚,开展苹果重要性状的遗传机制研究,进行相关基因挖掘及分子标记的开发,对于缩短苹果育种周期,提高苹果育种效率,早日实现我国苹果主栽品种本土化具有重要意义。果形是评价苹果果实外观品质的一个重要指标,通常高桩型苹果更符合大众的审美,更易被消费者接受。Brown认为苹果的果形是由多基因控制的数量性状,杂交种后代的平均果形指数值接近亲本的平均值[25]。刘志等研究发现,果形主要是以亲本的果形为主,同时杂交种实生苗果形有变圆趋势[26]。本研究杂交种实生苗果形指数的变异系数只有6.74%,杂交种实生苗果形以双亲为主,有向圆形果变异的趋势,这与刘志等的研究结果[26]一致。
可溶性固形物以及可滴定酸含量对于果实的口感、风味、营养价值等方面均有着重要的影响。刘家成等通过对杏杂交种实生苗遗传倾向分析后发现,可溶性固形物含量呈偏低遗传的倾向[27]。刘佳棽等通过对红芙蓉×中油5号杂交F1代遗传倾向分析发现,可溶性固形物平均值虽然比双亲本低,但也有少数单株存在超双亲现象;另外果实出现偏酸型占优势的现象[28]。赵习平等通过研究杏杂交种实生苗的遗传变异规律发现,可溶性固形物呈连续变异且有衰退的趋势,但也有部分单株高于高亲值[29]。本研究中可溶性固形物超低亲率为44.09%,可滴定酸超低亲率为65.59%,但同时也有少许单株高于高亲值。由上述可知,虽然杂交种实生苗出现了加性效应数量性状的解体,但是由于新的累加效应,在后代中出现了超高亲株系,说明晚白海棠×华月杂交种实生苗中有望选育出高可溶性固形物、酸度适宜的优良个体。
本研究所用母本晚白海棠单果质量不足父本华月单果质量的1/4,F1代杂交群体单果质量的变异系数为41.20%,其中超低亲占比超过84%,表现出明显的性状退化趋势。此前,多位研究者利用不同研究群体也得到了类似的结果。例如,韩婷婷等以金冠×红勋1号杂交F1代群体为试材,单果质量的变异系数在0.5以上,杂交群体中单果质量趋向于小果型亲本[10]。此外,新疆红肉苹果与苹果品种杂交种F1群体的果实性状分析[11]和富士杂交后代果实大小的调查[26]中均获得了类似的研究结果。本研究认为,从育种者的角度来讲,果实变小可能是一种不良的表型,但从苹果生存和进化的角度看,却可能是一种进化的趋势;因为果实单果质量虽然变小,但单个果实中的种子数量并不会随之减少,这就意味着树体在生产相同质量果实的情况下,可以产生更多的种子,从而使个体有更多的繁殖后代的机会。另外,尽管杂交种实生苗果实趋小变异已被广泛证实,但不同组合之间所获得的QTL位点却存在明显差异。例如,Potts等在Co-op 17×Co-op 16分离群体第3、5连锁群上检测到2个与果实大小性状连锁的QTL[30];Kostick等利用蜜脆衍生种质资源将果实大小QTL定位于8号和16号染色体上[31];Liu等利用富士×红肉苹果F1代将果实大小QTL定位于第6、17连锁群[32]。本研究对晚白海棠×华月苹果杂交种实生苗的若干性状进行了遗传分析,旨在为相关性状进行QTL分析及候选基因筛选奠定基础,同时也获得了多个性状优良的个体。
参考文献:
[1]陈 红,王 倩,高 强. 我国苹果产业发展及其影响因素分析:基于7个主产省份的面板数据[J]. 中国果树,2019(1):92-95.
[2]董志丹,宋尚伟,宋春晖,等. 我国育成苹果品种的系谱分析及其育种启示[J]. 中国农业科学,2020,53(21):4485-4496.
[3]闫忠业,伊 凯,刘志,等. 红玉×金冠苹果杂交后代果实糖酸组分遗传分析[J]. 果树学报,2017,34(2):129-136.
[4]赵亚楠,欧春青,王 斐,等. 梨F1代群体果实性状的遗传分析及相关性研究[J]. 西北农业学报,2018,27(12):1811-1818.
[5]王亚杰,孟 蕊,武月妮,等. 秦冠、富士苹果杂交后代果实性状遗传趋势分析[J]. 西北农业学报,2014,23(4):52-59.
[6]崔艳波,陈 慧,乐文全,等. 京白梨与鸭梨正反交后代果实性状遗传倾向研究[J]. 園艺学报,2011,38(2):215-224.
[7]赵崇斌,郭乙含,李舒庆,等. 宁海白×大房枇杷F1杂交群体果实性状的相关性及遗传分析[J]. 果树学报,2021,38(7):1055-1065.
[8]刘政海,董志刚,李晓梅,等. 酿酒葡萄黑比诺与马瑟兰杂交后代果实性状遗传倾向分析[J]. 中国果树,2020(6):29-35.
[9]刘振中,樊红科,高 华,等. 苹果杂交后代果实性状遗传变异分析[J]. 北方园艺,2012(5):5-8.
[10]韩婷婷,杨天资,赵培磊,等. 金冠和红勋1号杂交后代遗传多样性分析[J]. 青岛农业大学学报(自然科学版),2021,38(1):1-6.
[11]陈学森,张 晶,刘大亮,等. 新疆红肉苹果杂交种一代的遗传变异及功能型苹果优株评价[J]. 中国农业科学,2014,47(11):2193-2204.
[12]王家珍,李俊才,沙守峰,等. 南果梨×红巴梨正反交后代遗传倾向分析[J]. 北方园艺,2022(18):42-46.
[13]刘有春,鄂辉邦,刘 成,等. 越橘半同胞系杂交后代果实糖酸性状的变异和遗传倾向[J]. 果树学报,2016,33(6):664-675.
[14]吴 斌,张越根,黄东梅,等. 黄金百香果杂交子代果实性状遗传倾向分析[J]. 果树学报,2022,39(9):1587-1596.
[15]刘政海,董志刚,李晓梅,等. 威代尔与霞多丽葡萄杂交F1代果实性状遗传倾向分析[J]. 果树学报,2020,37(8):1122-1131.
[16]王 勇,伍国红,李玉玲,等. 红地球葡萄杂交F1代重要果实性状遗传倾向分析[J]. 果树学报,2015,32(6):1099-1106.
[17]王 燕,刘针杉,张 静,等. 中国樱桃杂交F1代花和果实若干性状遗传倾向分析[J]. 园艺学报,2022,49(9):1853-1865.
[18]黄爱萍,邓朝军,陈秀萍,等. 龙眼杂交后代果实性状遗传变异研究[J]. 福建农业学报,2006,21(4):342-345.
[19]李明章,邱利娜,王丽华,等. 红阳猕猴桃杂交F1代果实主要经济性状遗传倾向分析[J]. 果树学报,2011,28(1):51-54,189.
[20]韩 飞,赵婷婷,刘小莉,等. 山梨猕猴桃与中华猕猴桃种间杂交后代果实性状的遗传倾向分析[J]. 植物科学学报,2022,40(4):505-512.
[21]王 强,杨振英,张利义,等. 苹果新品种华月[J]. 中国果树,2013(6):34-36,86.
[22]杨振英,康国栋,王 强,等. 黄色苹果新品种华月[J]. 园艺学报,2010,37(11):1877-1878.
[23]王业社,侯伯鑫,索志立,等. 紫薇品种表型多样性分析[J]. 植物遗传资源学报,2015,16(1):71-79.
[24]王 昆,刘凤之,曹玉芬,等. 苹果种质资源描述规范和数据标准[M]. 北京:中国农业出版社,2005,:49-64.
[25]Brown A G.The inheritance of shape,size and season of ripening in progenies of the cultivated apple[J]. Euphytica,1960,9(3):327-337.
[26]劉 志,伊 凯,王冬梅,等. 富士苹果果实外观品质性状的遗传[J]. 果树学报,2004,21(6):505-511.
[27]刘家成,章秋平,牛铁泉,等. 串枝红与赛买提杏正、反交后代果 实性状遗传倾向分析[J]. 果树学报,2020,37(5):625-634.
[28]刘佳棽,王尚德,蒋海月. 红芙蓉×中油5号组合F1果实主要性状的遗传倾向研究[J]. 安徽农业科学,2012,40(21):10794-10795.
[29]赵习平,马之胜,宣立锋,等. 杏杂交后代果实性状的遗传变异研究[J]. 河北农业科学,2005,9(1):28-31.
[30]Potts S M,Khan M A,Han Y P,et al. Identification of quantitative trait loci (QTLs) for fruit quality traits in apple[J]. Plant Molecular Biology Reporter,2014,32(1):109-116.
[31]Kostick S A,Luby J J.A pple fruit size QTLs on chromosomes 8 and 16 characterized in ‘Honeycrisp-derived germplasm[J]. Agronomy,2022,12(6):1279.
[32]Liu Z C,Bao donge,Liu D L,et al. Construction of a genetic linkage map and QTL analysis of fruit-related traits in an F1 red Fuji×Hongrou apple hybrid[J]. Open Life Sciences,2016,11(1):487-497.
猜你喜欢
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:08
临床医药文献杂志(电子版)(2017年11期)2017-05-17 04:47:56
小学阅读指南·低年级版(2016年11期)2017-02-06 23:28:39
作文与考试·小学高年级版(2016年17期)2016-12-23 20:21:17
小朋友·快乐手工(2016年10期)2016-12-08 06:15:06
小学生导刊(低年级)(2016年8期)2016-09-24 22:22:30
阅读与作文(小学低年级版)(2016年3期)2016-03-08 22:20:53
中学生物学(2016年8期)2016-01-18 09:08:21
水土保持通报(2014年5期)2014-06-09 08:27:06
新疆农垦科技(2014年6期)2014-02-28 19:20:08
