
入侵性害虫桦潜叶蜂生活史及土壤生态适应性*
2023-08-09 03:01邢蒙恩房鸿嫄刘瀚予杜志琦孙艳梅范文忠冯立超
林业科学 2023年5期
胡 靓 邢蒙恩 房鸿嫄 刘瀚予 杜志琦 王 楠 孙艳梅 范文忠 冯立超
(1. 吉林农业科技学院 吉林 132101;2. 北华大学 吉林 132013)
桦潜叶蜂(Profenusa thomsoni)最早在古北区英国、日本发现为害白桦(Betulaplatyphylla),随后传播至北美地区,发生大规模为害(Britton, 1924;Ross, 1936)。以幼虫潜食叶肉为害白桦叶片,严重时造成叶片脱落,叶片受害率达90%以上(Benson, 1959)。相关的研究报道,成虫活动受温度和相对湿度影响较大,温度为21 ~ 22 ℃和相对湿度70 %时最适宜产卵,行动活跃(Martin, 1960)。在加拿大及美国阿拉斯加地区,幼虫取食至6 龄后脱叶,在3~12 cm 土层内或落叶层近地表做土茧进入前蛹期(Digweedet al., 2009)。因此,桦潜叶蜂以老熟幼虫脱叶进入树盘周围土壤内做土茧,土壤生境对翌年春季成虫羽化出土具有重大影响。同时,通过对不同区域桦潜叶蜂为害率调查结果表明,在城市中的白桦比城郊村庄及天然林为害率更高(Digweedet al., 1997),这可能是由人类活动及地理隔离等的多重因素所主导,形成城市集中分布模式(Digweedet al., 2004)。同时,文献记载桦潜叶蜂主动扩散能力较弱(Digweedet al ., 1997)。在防控研究中,其中姬蜂科 (Ichneumonidae) 的Lathrolestes thomsoni和Lathrolestes soperi2 种寄生蜂在加拿大等地区可以有效地控制该害虫为害(Andersenet al., 2021),土壤中的寄生性真菌和捕食性线虫对桦潜叶蜂控制较差(Progaret al., 2015)。因此,研究桦潜叶蜂土壤中越冬虫态习性及生态环境因子对其生存的影响,对于入侵性害虫的防控具有重要意义。
目前,对桦潜叶蜂土壤中生活习性相关研究较少,且未见土壤理化性质对其生存影响相关报道。鉴于此,本文研究土中越冬的幼虫与土壤理化性质相关性,探讨土壤越冬桦潜叶蜂对土壤性质的适应性,明确土壤环境因子(含水量、温度、化学性质)对桦潜叶蜂生存及发育的影响。
1 材料与方法
1.1 物种基因鉴定及生物学特性观察
为明确吉林地区入侵的白桦潜叶蜂分类地位,通过形态特征与DNA(COI)序列比对,鉴定物种。采集成虫(孤雌生殖)3 头,用QIAGEN Blood & Tissue 试剂盒 (德国),根据无损法提取DNA(Taekulet al., 2014),保留提取过DNA 的样品作为凭证标本,供形态鉴定。使用徕卡DVM6 体视镜、联用Axiocam 506 color 数码照相系统(德国徕卡显微系统有限公司)对凭证标本拍摄,结果保存于吉林农业科技学院。
获得全基因组DNA 后,采用线粒体基因COI 通用引物(COI-F: 5' -CAACATTTATTTTGATTTTTTGG-3'和COI-R: 5' -TCCAATGCACTAATCTGCCATATTA-3')COI 进行PCR 扩增(Simonet al., 1994)。扩增体系为25 µL:PCR Master Mix(2×)12.5 µL,DNA 模板1 µL,上游和下游引物各0.5 µL,最后用ddH2O 补足至25 µL。PCR 扩增条件:94 ℃预变性3 min;94 ℃ 30 s,50 ℃30 s,72 ℃延伸30 s,共35 个循环;72 ℃后延伸5 min。PCR 产物取5 µL 1%的琼脂糖凝胶电泳检测,由上海生工生物工程有限公司采用桑格测序法 (Thermo ScientificTMGeneRuleTMr DNA Ladder Mix 测序仪,美国)进行双向测序。获得的原始序列在Geneious 软件中进行拼接并检查是否含终止密码子,排除假基因。对COI 序列与Wei,Taeger,Hari 等所发表的物种基因序列进行比对分析(Weiet al., 2006;Taeger, 2010;Haris,2021)。在MEGA 软件中使用MUSCLE Alignment 算法对序列进行并对,并用邻接法(Neighbor-Joining)构建进化树。
2020 年5—8 月,在实验室内和野外采用对比观察法调查记录虫体生物学特性。实验室内使用恒温培养箱,温度设置为(20 ± 1)℃,相对湿度75%,光周期模拟野外为白昼 : 黑夜 = 12 h∶12 h;在5 个培养箱内分别移入10 组盛有白桦林下原位土壤培养盒(长30 cm×宽20 cm×深15 cm),记录成虫羽化时间。羽化后成虫于养虫笼内使用新鲜白桦叶进行饲养,观察并记录产卵、幼虫生活史等特性。同时,在白桦林内观察记录对比观察野外潜叶蜂为害情况。
1.2 试验设计
选取吉林农业科技学院白桦种植林、吉林市绿化管理处第一苗圃白桦种植林,面积各5 000 m2,2 地相距4.8 km,进行野外调查,试验区远离大型工厂、居民建筑群等。试验保护区宽10 m,采用平行行法于地表设置60 目尼龙网罩,样方长30 cm×宽20 cm×深15 cm(Soperet al., 2015),相邻样点间隔10 m,采样点网罩布置图(图1),每个试验区各设置36 个采样点。

图1 潜叶蜂土壤越冬后成虫羽化定量观察Fig. 1 Quantitative observation of leaf miner adult emergence after overwintering
1.3 不同发育阶段虫量统计
越冬前老熟幼虫及越冬后羽化成虫统计。于采样地点设置面积大小一致的网笼布置于地表,在老熟幼虫集中脱叶阶段,统计网笼内掉落的幼虫数量。入土5 天后,采集网罩样方毗邻区域树盘周围面积大小相同的土壤样品,每样地区域共采集36 个。采用显微镜镜检土壤中幼虫数量,统计越冬前土壤内做土茧幼虫数量。春季越冬后成虫羽化出土阶段,统计网罩采样点处成虫羽化量。每天统计样方网罩内成虫羽化出土数量直至无成虫出土。
1.4 土壤理化性对桦潜叶蜂生存影响
为研究土壤含水量对桦潜叶蜂幼虫化蛹及成虫羽化的影响,于2020 年5 月白桦展叶初期,土壤平均温度为15 ℃左右时设2 个试验区。在吉林农业科技学院设置的样地内采用人工洒水车对白桦进行浇水,间隔3 天浇水1 次,参考吉林市5—6 月年平均降水量,每次浇水6.8 L·m-2,至成虫羽化。每天测定2个样地不同样点土壤15 cm 深处土壤含水量变化。以吉林市绿化管理处第一苗圃白桦样地处理作为对照,不进行浇灌水处理。分别采集2个试验地0~15 cm 的土壤各36 份,每份500 g,待用,采用常规试验方法进行测试。
采用碳氮分析仪Vario MACRO cube 测定土壤全氮(TN)和全碳(TC)(德国Elementar 分析传感系统股份有限公司);采用ICP-AES 9800 测定土壤总磷(TP)(日本岛津公司)。采用玻璃指示电极法[瑞士梅特勒—托利多国际贸易(上海)有限公司]测定土壤酸碱度(pH)(Spitzeret al., 2011);土壤含水量(SWC)采用烘干法(105±1) ℃烘干12 h 至恒重(Wilke,2005);土壤地温计用于检测采样点内土壤温度(ST)。
1.5 数据分析
采用t检验(Holm-Šídák analysis method for multiple comparisons)比较2 分布地白桦林内桦潜叶蜂羽化量差异性,显著性差异水平(P<0.01);采用相关性分析方法(correlation analysis)分析桦潜叶蜂土壤内越冬存活量与土壤含水量的相关性关系,同时结合冗余分析RDA(redundancy analysis)分析土壤理化性质对桦潜叶蜂的生存影响,采用蒙特卡洛检验 (Monte Carlo methods)检验对羽化的影响是否显著。
2 结果与分析
2.1 物种鉴定
采用形态学与线粒体DNA 细胞色素氧化酶亚基(COI)基因序列片段对比相结合进行物种鉴定,结果见图2。

图2 桦潜叶蜂相近属系统发育树及成虫鉴定特征Fig. 2 Leaf miner phylogenetic tree and adult identification characteristics
形态特征:雌性,成虫头部黑褐色,触角丝状11 节。前翅第3臀脉与第一臀脉远离;后翅第一缘室和臀室为开室;翅基片为黑色。中胸盾片中叶突具侧毛;第一腹节背板后缘弧形弯曲,亚中部最宽。锯状产卵器,橙黄色—深褐色,具明显的弧形锯齿。卵,初产白色,于叶片表皮内吸水膨胀后呈半透明,大小为0.5 mm;孵化过程颜色逐渐变深,孵化前为棕色。幼虫:初孵幼虫长1 mm。头部棕褐色,三角形;背腹扁平,前突,重度硬化。前胸宽于头部,腹面有一个大的中央和2个侧面的深棕色斑纹。虫体自前胸至腹部逐渐变细,透明可见肠道。中胸和后胸腹侧有小的椭圆形棕色斑纹。胸部和腹部的气门呈棕色,突出。老熟幼虫体呈圆柱形,暗白色,半透明。蛹:化蛹于幼虫分泌物黏贴所形成的土茧内,蛹长4~5 mm,复眼红棕色,头部半透明白色,体躯呈淡黄色。末期逐渐变成蓝灰色,最后变成黑色。
经成虫COI 序列测定,SC855_1基因序列与GenBank 数据库中Profenusa thomsoni序列100%的相似度 (图2)。
生活史及习性:桦潜叶蜂在吉林地区年发生2 代。6 月上旬,始见羽化的成虫,在白桦的下层幼嫩叶片以幼儿虫潜叶危害;6 月下旬至7 月上旬,第一代幼虫大多数幼虫脱叶落入浅土层中,少数于叶片内化蛹,并羽化出成虫,继续危害白桦中、上部嫩叶,8 月下旬至9 月上旬,以老熟幼虫脱叶入土,主要在5~10 cm 做土茧越冬;世代不整齐。观察发现,在干旱年份直至降雨成虫才陆续羽化。
成虫喜树冠下层叶片产卵,卵产于叶片上表皮和栅栏层之间,平均每叶4~6 枚卵,偶有10 枚卵以上。成虫虫口密度大,全部叶片均会被产卵,多虫在同一叶片产卵,但新叶不会被利用。空气相对湿度高于70%,温度20~25 ℃利于成虫产卵。第2代成虫羽化后逐渐向树冠中、上层叶片产卵。卵孵化时间为10 天。幼虫共6 龄;老熟幼虫脱叶后掉落至地表,在土壤缝隙及浅土层中做土茧。
2.2 桦潜叶蜂土壤中不同发育阶段虫量变化
桦潜叶蜂老熟幼虫脱叶入土量、入土后做茧量及成虫羽化量变化见图3。

图3 不同区域桦潜叶蜂幼虫越冬阶段及越冬后虫体存活量Fig. 3 Overwintering stages and post-overwintering survival of P.thomsoni larvae in different regions
通过对冬前土壤中幼虫做茧情况调查表明,越冬前2样地白桦林土壤内平均做茧率分别86.47%和89.26%,在2样地内老熟幼虫做茧量无差异(P>0.05);翌年春季浇水处理后,浇水样地(Plot 2)成虫羽化量极显著高于对照样地(Plot 1)羽化量(P<0.01)(图3)。
2.3 土壤理化性对桦潜叶蜂的影响
试验地人工浇水及对照处理,桦潜叶蜂成虫羽化量对比分析见图4。

图4 土壤含水量及温度与桦潜叶蜂羽化量关系Fig. 4 P. thomsoni eclosion in relation to soil water content and temperature
不同试验区样方的土壤含水量对羽化的影响分析,差异极显著(P<0.01)(图4a);成虫羽化量与土壤含水量呈显著正相关关系(P<0.01,R=0.85)(图4b)。对照样地内土壤含水量低,桦潜叶蜂成虫羽化量受到抑制。土壤pH、氮含量与羽化量正相关(图5a);人工浇水处理提高了土壤含水量,对成虫羽化具有显著促进作用(P<0.01),而土壤化学性质对成虫羽化量影响较小(图5b)。
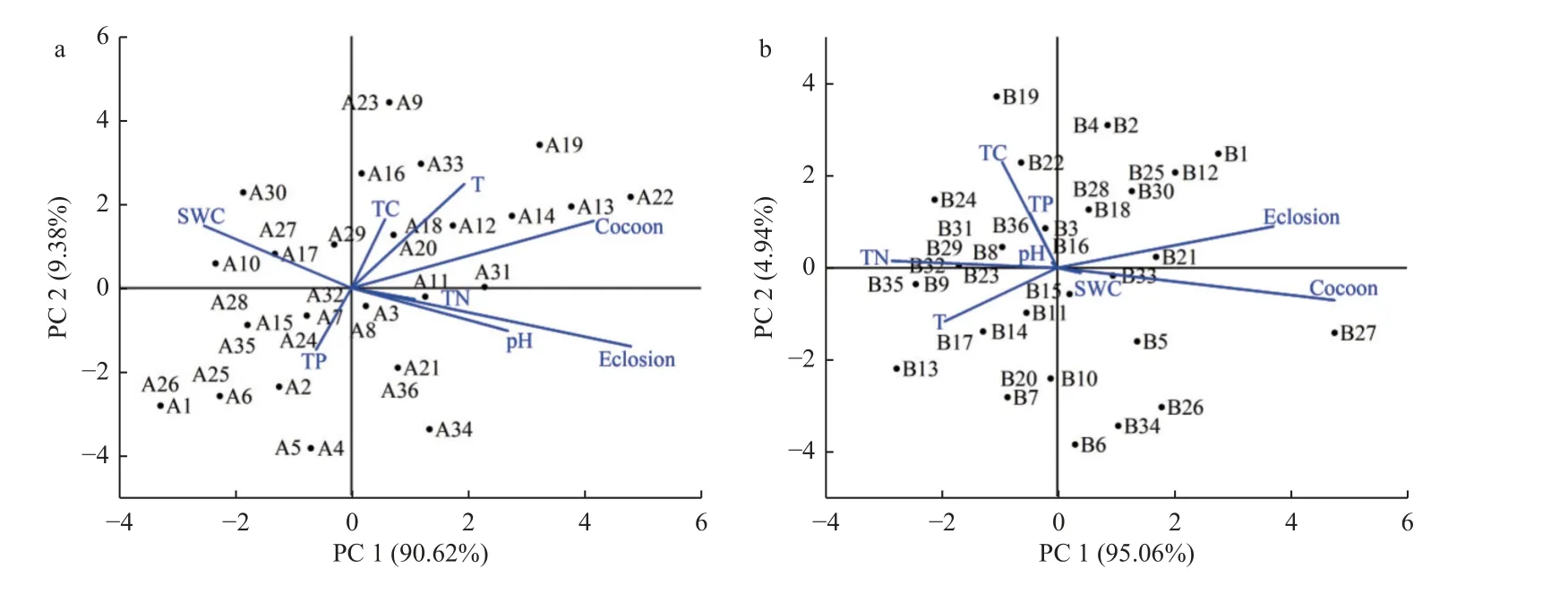
图5 土壤理化性质对桦潜叶蜂羽化作用Fig. 5 Effect of soil physicochemical properties on adults eclosion
3 讨论
3.1 气候因子对桦潜叶蜂分布决定作用
桦潜叶蜂在北美地区每年发生1 代,以孤雌生殖形式完成繁殖(Digweed, 2006)。在吉林地区每年发生2 代,第一代化蛹于叶片上表皮和栅栏层之间,第2代化蛹于土壤内。通过对吉林市区及周边山区、人工林及天然次生林白桦危害情况调查,发现仅在市区内2个区域发生为害,发生面积约为12 hm2,分布范围受限的主要原因是其传播主要途径为人为传播,而主动飞行扩散能力较弱(Digweedet al., 2009;Digweedet al., 2004)。因此,桦潜叶蜂为害发生的主要区域与人类活动频繁或集中性有关。
吉林地区(42°—44°N)冬季平均温度比阿拉斯加和加拿大(54°—71°N)高,土壤内越冬温度对桦潜叶蜂越冬的致死率会降低。适应低温生存的昆虫在高温区生存受到抑制,限制了物种分布区域(Coulsonet al., 1996;Irwinet al., 2000)。因此,桦潜叶蜂在北半球由高纬度向低纬度地区入侵过程中,随着温度升高,其分布区域可能会受限。在本研究中,土壤温度几乎未影响成虫羽化,可能是由于春季土层温度维持在10~15 ℃,此温度区间有利于大多数土壤动物生存(Sulkavaet al., 1996)。在冬季,森林土壤内10 cm 土层内温度在-3 ℃以上(Weigelet al., 2021),由于动物体内代谢及其他生理变化对机体组织的保护,此温度并未对大多数土壤动物构成死亡威胁(Sulkavaet al.,2003)。水分是生物体的必要物质,适宜的土壤含水量是维持土壤动物生存的基本条件;遇干旱胁迫时,绝大多数土壤动物都会改变自身生存状态,通过降低代谢速率等生理活动,进入休眠、滞育等生存状态,以最大程度的适应土壤环境(Danks, 2000;Holmstrup,2014)。春夏交替季节是桦潜叶蜂成虫羽化出土的时间,在本研究中,受降水影响,2020 年5—6 月吉林地区降水量严重减少,土壤表层含水量严重降低,平均含水量不足14%。人工浇水处理区,土壤含水量维持在14.3%~30.8%之间,直接改变了桦潜叶蜂羽化时间,提高羽化量。以上结果表明,温度升高导致水分的加速散失,对冬季生存的节肢动物构成严重威胁(Everattet al., 2015)。因此,土壤含水量在干旱季节决定了桦潜叶蜂对生存选择,土壤含水量是限制虫口密度的关键抑制因子。
3.2 土壤化学性质及生物因子对桦潜叶蜂生存影响
白桦林下土壤其他化学性质几乎不影响桦潜叶蜂越冬幼虫成活率,这可能是由于老熟幼虫脱叶入土做土茧期间分泌黏液类物质,将周围土壤颗粒黏聚于体躯外表形成土茧,而本身并不直接与土壤长期接触,且体壁分泌黏液所形成的保护层直接隔离外部土壤与体躯直接接触(Martin, 1960),因此,对其生存影响较弱。本研究表明,在自然条件下,土壤含水量与桦潜叶蜂的成虫羽化量和羽化时间相关,含水量相对较高的区域羽化量大,羽化时间提前;相反羽化量降低,羽化时间延长。此外,由于土壤内复杂的地下生存环境条件;尽管土壤化学性质如pH、碳、氮、磷等物质对桦潜叶蜂影响较小,但天敌及微生物对进入前蛹的虫体的捕食或寄生作用,也是制约翌年成虫羽化的影响因素之一(Progaret al., 2015;Soperet al., 2019;Schönroggeet al., 1992)。
本研究通过对土壤含水量控制,明确桦潜叶蜂越冬虫态对水分的适应性。在相对可控的土壤环境条件下,通过树盘土壤翻耕,破坏土茧周围土壤水分环境,对削减虫口密度具有一定的作用。对于人为传播等控制措施,加强对土壤、白桦苗木调运等检疫措施,以及合理使用杀虫剂。由于本研究样地处于同一区域并无差异,因此温度对成虫羽化的作用并未体现。桦潜叶蜂在吉林地区首次发现为害白桦,受其研究范围受限,对于桦潜叶蜂的扩散、大面积发生为害等与当地气候、白桦栽培情况、生态环境以及寄生性微生物和天敌等关系研究需进一步开展。
4 结论
桦潜叶蜂是一种入侵性有害生物,主要危害白桦,在吉林地区每年发生2 代,以幼虫在土壤内越冬。土壤理化性质是决定桦潜叶蜂翌年成虫发生量关键影响因素,含水量相对较高的土壤促进了桦潜叶蜂越冬虫态的发育,而相对干旱的土壤抑制成虫羽化,羽化时间延长。
猜你喜欢
作文新天地(小学版)(2022年8期)2022-05-30
山西林业(2021年2期)2021-07-21
文苑(2019年20期)2019-11-20
文苑·经典美文(2019年10期)2019-10-16
山东林业科技(2019年2期)2019-06-03
第二课堂(初中版)(2018年7期)2018-09-08
湖北植保(2017年4期)2017-08-31
小溪流(画刊)(2017年6期)2017-07-10
华人时刊(2016年16期)2016-04-05
贵州林业科技(2016年4期)2016-03-02
