
长白山地区黄檗阔叶混交林主要乔木种群生态位特征
2023-08-08 03:29栾兆平郭滨德吴金卓
森林工程 2023年4期
关键词:黄檗
栾兆平 郭滨德 吴金卓


摘 要:黄檗是国家二级保护植物。通过定量分析黄檗阔叶混交林中主要乔木种群的生态位特征,可以为黄檗的保护和维持物种多样性提供理论依据。基于吉林省第九次森林资源连续清查数据,通过对长白山地区黄檗阔叶混交林群落的典型样地进行调查,利用重要值、Levins生态位宽度、Shannon-Wiener生态位宽度和Pianka生态位重叠度计算方法,分析黄檗阔叶混交林主要乔木种群的生态位特征。结果表明,长白山地区黄檗阔叶混交林有25个阔叶树种,占比96.15%,黄檗的重要值最大;Levins和Shannon-Wiener生态位宽度指数大小排序基本一致,生态位宽度最大的树种均为黄檗,其次为胡桃楸;黄檗、枫桦、胡桃楸和裂叶榆等物种的生态位重叠值较大,生态位宽度和生态位重叠度表现出明显的一致性。但是存在部分物种生态位宽度和生态位重叠度具有相反性,如鹅耳栎与槲树、刺槐、椴树,以及黄檗与红松,这与生态宽度受到物种生态习性、生境特征和空间分布等因素有关。
关键词:黄檗;生态位宽度;生态位重叠;重要值;阔叶混交林
中图分类號:S 文献标识码:A 文章编号:1006-8023(2023)04-0078-08
Niche Characteristics of Main Tree Populations in Broad-leaved Mixed
Forest of Phellodendron amurense in Changbai Mountain Area
LUAN Zhaoping1, GUO Binde1, WU Jinzhuo2*
(1.Forest Resources Monitoring Center of Key State-owned Forest Region, State Forestry and Grassland
Administration, Heilongjiang Jiagedaqi 165000, China; 2.College of Civil Engineering and Transportation, Northeast
Forestry University, Harbin 150040, China)
Abstract:Phellodendron amurense is national second-class protected plant. The quantitative analysis on the niche characteristics of dominant tree populations in broad-leaved mixed forest of Phellodendron amurense can provide theoretical basis for the protection of Phellodendron amurense and maintaining species diversity. Based on the 9th national forest inventory in Jilin Province, the Phellodendron amurense broad-leaved mixed forest community in Changbai Mountain was investigated and the methods of importance value, Levins niche width, Shannon-Wiener niche width and Pianka niche overlap were used to analyze the niche characteristics of dominant tree populations in this region. The results showed that, there were 25 broad-leaved species, accounting for 96.15% of the total, and the important value of Phellodendron amurense in the broad-leaved mixed forest was the largest. The Levins and Shannon-Wiener niche width indices were basically the same in order. The species with the largest niche width was Phellodendron amurense, followed by Juglans mandshurica. The niche overlap value of species such as Phellodendron amurense with Butula costata, Juglans mandshurica and Ulmus laciniata was large. The niche width and niche overlap degree showed obvious consistency. However, the niche width and niche overlap of some species were opposite, such as Carpinus turczaninowii vs. Quercus dentata, Robinia pseudoacacia, and Tilia tuan, Phellodendron amurense vs. Pinus koraiensis, which was related to the ecological habits, habitat characteristics and spatial distribution of the tree species.
Keywords:Phellodendron amurense; niche width; niche overlap; importance value; broad-leaved mixed foresd
收稿日期:2022-10-20
基金项目:国家林草生态综合监测评价(GXCZ-C-22780094)
第一作者简介:栾兆平,高级工程师。研究方向为国有森林资源监测。E-mail: 79697987@qq.com
通信作者:吴金卓,博士,教授。研究方向为森林作业与环境。 E-mail: wjz@nefu.edu.cn
引文格式:栾兆平,郭滨德,吴金卓.长白山地区黄檗阔叶混交林主要乔木种群生态位特征 [J]. 森林工程,2023,39(4):78-85.
LUAN Z P, GUO B D, WU J Z. Niche characteristics of main tree populations in broad-leaved mixed forest of Phellodendron amurense in Changbai Mountain area[J]. Forest Engineering,2023, 39(4):78-85.
0 引言
生態位是指一个特定群落的物种在一定时间空间上所占据的位置,能够揭示种间的相似程度,表示种群之间的相对关系和种群对于资源的利用程度。Grinnell于1917年给出了生态位的定义,随后Odum和Smalley对生态位的概念进行了完善[1],但是对于生态位内涵描述得比较抽象。彭文俊等[2]于2016年对生态学中生态位的内涵进行了系统研究。近年来,对于生态位理论的研究由昆虫、鸟类逐渐过渡到植物生态学领域[3],在种间排斥、物种共存和未来演替等方面发挥着关键的作用。
生态位理论是植物种群和群落生态学的重要理论之一[4-5],能够反映物种对光照、水分等环境资源的利用程度以及种间竞争程度,是解释种间共存现象以及种间竞争机制的基本理论[6]。生态位理论常用的量化指标主要包括生态位宽度和生态位重叠度。生态位宽度能够反映出群落中物种对环境资源的利用能力[7],物种具有较强的环境适应能力,分布范围较广泛,在群落中地位越高,对光照、水分等环境资源的利用越充分,生态位宽度就越大,反之,生态位宽度就小[8-9]。生态位重叠度能够体现生态位相似的物种间对环境资源的竞争关系[10-11]。生态位重叠度越高,说明2个物种对光照、水分等环境资源的需求和利用程度越相近,种间竞争也越激烈,产生竞争排斥现象;生态位重叠越低,说明2个物种对光照、水分等环境资源的需求和利用程度差异越大,种间竞争关系不明显[12-13]。生态位理论可以量化物种在群落中的地位,反映种群在生态系统中利用资源的能力、群落结构特征及未来演替方向[14]。因此,生态位理论在研究生物多样性保护及黄檗等保护物种的评价方面具有较高的生态学价值。
黄檗(Phellodendron amurense)是国家二级保护植物,为阳性树种,根系发达,对土壤的适应性较强,常生长在土层深厚、湿润、含腐殖质丰富的中性或微酸性土壤中。在长白山林区,主要分布在河谷及山地中下部的阔叶林或红松、云杉阔叶混交林中。本研究以长白山地区黄檗阔叶混交林为研究对象,通过计算Levins生态位宽度、Shannon-Wiener生态位宽度和生态位重叠度,定量分析黄檗阔叶混交林中主要乔木种群的生态位特征,了解主要乔木种群对水分、光照等环境资源的利用状况和种间竞争关系,以期为黄檗的保护和维持物种多样性提供一定的理论依据。
1 研究区概况
研究区位于吉林省境内长白山地区,该地区属于温带大陆性山地气候,冬季漫长而寒冷,夏季短暂而凉爽,年均温度为-7~3 ℃,7月份平均气温不超过10 ℃,最冷月份在1月,平均温度-20 ℃,年日照时数2 300 h左右,无霜期100 d。年降水量为700~1 400 mm,降水主要集中在6—9月份。该区域植被属长白山植物区系,地带性植物主要有长白落叶松(Larix olgensis)、红皮云杉(Picea koraiensis)、红松(Pinus koraiensis)、大青杨(Populus ussuriensis)和黄檗(Phellodendron amurense)等。
2 研究方法
2.1 样地设置及调查
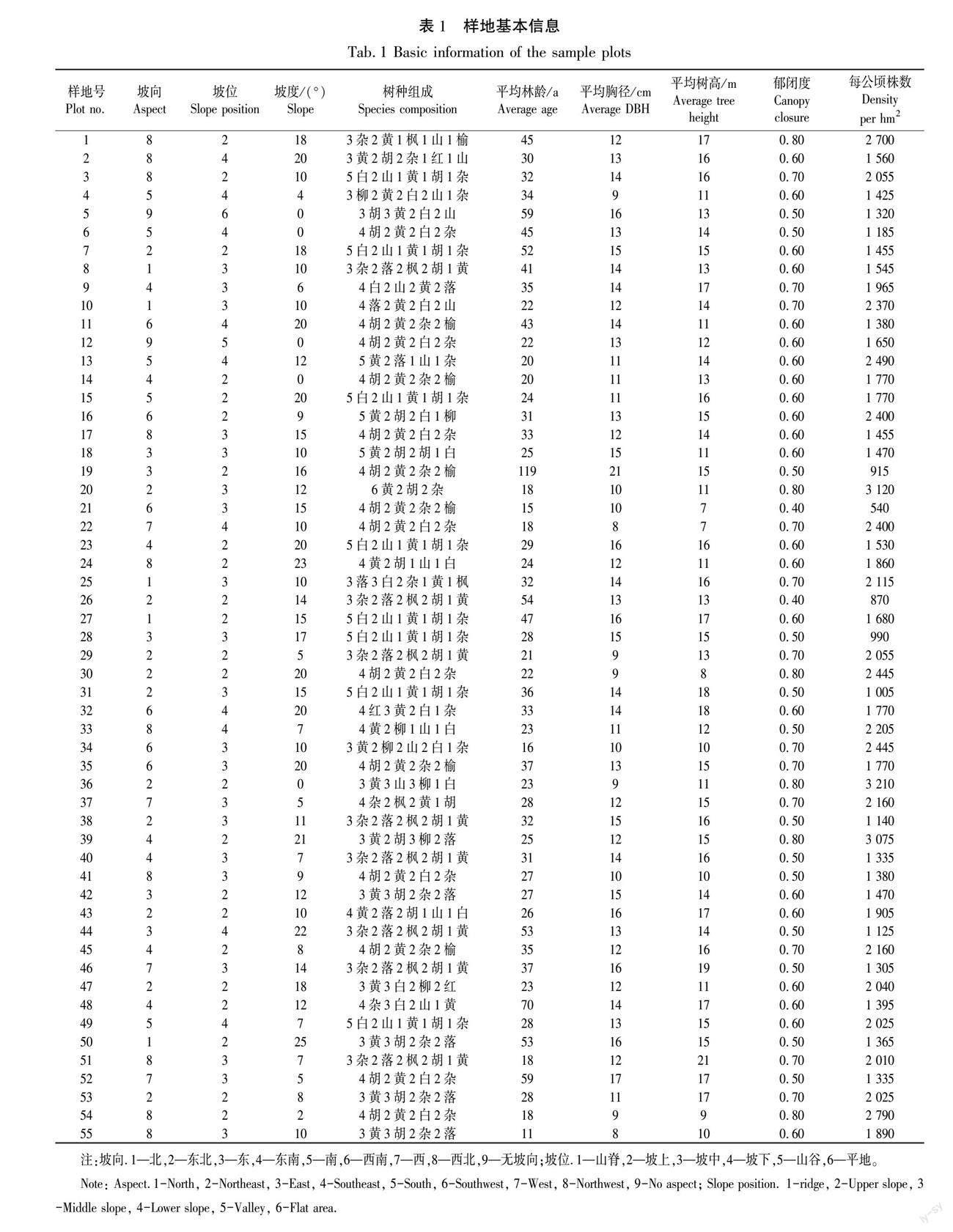
本研究利用吉林省第九次森林资源连续清查数据,在长白山地区选择55块正菱形黄檗阔叶混交林样地,样地大小为25.82 m×25.82 m,对样地内胸径≥5 cm的乔木树种进行每木检尺,并记录树种名称、胸径、树高等信息。样地基本信息见表1。
2.2 物种重要值计算
重要值是衡量物种多样性的指标,能够反映种群在植物群落中的重要程度和优势程度[15],可以通过相对多度、相对频度和相对优势度计算[16],计算公式如下
重要值=相对多度+相对频度+相对优势度3。 (1)
相对多度=某个物种的株数所有物种总株数×100%。 (2)
相对频度=某个物种的频度所有物种频度总和×100% 。 (3)
相对优势度=样方中该种个体胸高断面积和样方中全部个体胸高断面积总和×100%。(4)
2.3 生态位宽度计算
样地内资源状态可获得性和丰富度无法确定[17],因此本研究选择对丰富资源有更大权重的Levins[18]生态位宽度指数(BL)和对资源有更大权重的Shannon-Wiener[19]生态位宽度(BSW)来描述黄檗阔叶混交林主要乔木种群的生态位宽度。
BL和BSW计算如下所示。
BL=1N×∑rj=1P2ij。(5)
Bsw=∑rj=1(P2ij×lnPij)。(6)
式中:Pij表示物种i在样方j中的重要值占所有样方重要值总和的比例;N代表样方的数量;BL的阈值为[0,1],BL的值越趋向1,说明该物种的生态位宽度越宽;BSW的阈值为[0,lnN],BSW的值越趋向lnN,说明该物种的生态位宽度越宽[20]。
2.4 生态位重叠度计算
生态位重叠度能够反映不同物种在光照、营养和生存空间等方面对可利用的环境资源的竞争,表现为不同物种对某一环境资源在利用上的差异,计算公式如下
Oik=∑rj=1Pij×Pkj
∑rj=1P2ij×∑rj=1P2kj。 (7)
式中:Oik表示物种i和物种k的生态位重叠度,其阈值为[0,1],当Oik=0时,说明2个物种的生态位完全不重叠;当Oik=1时,说明2个物种的生态位完全重叠,Pij表示物种i在样方j中的重要值占所有样方重要值总和的比例;Pkj表示物种k在样方j中的重要值占所有样方重要值总和的比例。
3 结果与分析
3.1 主要乔木种群的重要值特征
黄檗阔叶混交林主要乔木树种重要值见表2。在调查的55个固定样地中,黄檗阔叶混交林主要乔木树种共26种6 588株,有25个阔叶树种,占比96.15%;重要值≥1%的乔木树种共23种,合计占比为97.87%,其中,排在前4位的优势树种分别为黄檗(14.84%)、胡桃楸(Juglans mandshurica,9.41%)、山杨(Populus davidiana,9.32%)和白桦(Betula platyphylla,7.42%),黄檗的重要值远大于其他物种。后4种弱势树种分别为榆树(Ulmus pumila,1.58%)、刺槐(Robinia pseudoacacia,0.80%)、稠李(Prunus padus,0.80%)和赤杨(Alnus japonica,0.53%)。
3.2 主要乔木种群的生态位宽度
生态位宽度是指植物群落中物种利用环境资源的总和,能衡量不同物种对环境资源的利用情况,反映不同物种的生态适应性和分布幅度。由图1可知,黄檗阔叶混交林主要乔木种群的重要值和生态位宽度大致呈正相关。由表3可知,在55块固定样地中,黄檗阔叶混交林主要乔木树种的Levins和Shannon-Wiener生态位宽度指数大小排序基本一致。重要值最大的黄檗,其生态位宽度(BL、BSW)也最大,其值分别为0.61和3.71;其次为胡桃楸,其值分别为0.49和3.46;前4种优势树种中,山杨和白桦的生态位宽度出现差异。鹅耳栎(Carpinus turczaninowii)的生态位宽度均最小,其值分别为0.05和1.26,后4位弱势树种的生态位宽度均出现了上升。
3.3 主要乔木种群的生态位重叠
黄檗阔叶混交林主要乔木树种生态位重叠见表4。由表4可知,群落乔木树种的重叠均值为0.28,重要值大于0.8的有5对,最大为刺槐和辽东栎(Quercus wutaishansea,0.96),其次为鹅耳栎与槲树(Quercus dentata,0.95)、刺槐和鹅耳栎(0.91)、黄檗和槲树(0.89)、黄檗和枫桦(0.88)。生态位宽度和生态位重叠之间存在一定的相关性,如生态位宽度最大的黄檗与枫樺、胡桃楸、裂叶榆等物种的生态位重叠值较大,分别为0.88、0.92和0.79;生态位宽度较小的鹅耳栎与赤杨、桑树(Morus alba)和山丁子(Malus baccata)等物种的生态位重叠值为0。但有些生态位宽度较小的物种之间,其生态位重叠值却较大,如鹅耳栎与槲树、刺槐、椴树的生态位重叠值分别为0.95、0.91、0.66;有些生态位宽度较大的物种之间,其生态位重叠值却较小,如黄檗与红松的生态位重叠值为0.03。
4 讨论
4.1 主要乔木种群的重要值和生态位宽度
重要值是表示物种在群落中重要性的指标[21]。由调查可知,群落内共有25个阔叶树种,占96.15%,黄檗为群落内的最优势种,在群落中占主导地位,这是因为所选区域是黄檗阔叶混交林,符合所选样地的特征。
生态位宽度受到种间竞争强度、物种对自然环境的适应能力及自然环境的分布状况等因素的共同影响[22],能够反映物种对资源的利用程度和环境的适应能力和在群落中的地位和分布情况[23]。本研究结果发现,黄檗阔叶混交林主要乔木树种的Levins 和Shannon-Wiener生态位宽度指数大小排序基本一致,生态位宽度最大的种群均为黄檗,其值分别为0.61和3.71。说明黄檗对光照、水分等环境因素的适应能力最强,对群落中资源的利用能力最好,在群落中的分布更为广泛,在整个群落中处于优势地位[24],对黄檗阔叶混交林群落的构建起到较为重要的作用。重要值和生态位宽度没有绝对的正相关或负相关关系,判断群落中物种对环境资源的利用状况需要考虑重要值和生态位宽度,并且生态位宽度还受到频度、空间、时间及环境因素的影响[25]。黄檗阔叶混交林主要乔木种群的重要值和生态位宽度大致呈正相关,但有些种群不符合这种关系,这与周梅妹等[26]研究结果相似。黄檗阔叶混交林中白桦具有较高的重要值,但生态位宽度相对较小,这可能是由于白桦是萌生,分布较为集中,因此其作用相对于生态位较宽的物种而言要弱。
4.2 主要乔木种群的生态位重叠
生态位重叠能够反映不同种群对资源利用的相似性和对空间位置占据的交错程度,在一定程度上可以表明种间在生长过程中的竞争关系[27],生态位重叠程度越高说明群落中不同物种间对自然资源的利用越相似,可能发生的种间竞争也越激烈[28]。陈玉凯等[22]研究发现,生态位越宽,对自然资源的利用能力就越强,同时与群落中其他物种的生态位重叠程度也越大,本研究中生态位宽度最大的黄檗与枫桦、胡桃楸和裂叶榆等物种的生态位重叠值较大,分别为0.88、0.92和0.79,符合此规律。同时,本研究还发现,有些生态位宽度较小的物种之间,其生态位重叠值却较大,如鹅耳栎与槲树、刺槐、椴树的生态位重叠值分别为0.95、0.91、0.66;有些生态位宽度较大的物种之间,其生态位重叠值却较小,如黄檗与红松的生态位重叠值为0.03,这可能是与群落中不同种群的环境分异及物种自身的生态习性相关。
5 结论
本研究基于吉林省第九次森林资源连续清查数据,通过对长白山地区黄檗阔叶混交林群落的典型样地进行调查,利用重要值、Levins生态位宽度、Shannon-Wiener生态位宽度和Pianka生态位重叠度计算方法,分析了黄檗阔叶混交林主要乔木种群的生态位特征,得出主要结论如下。
长白山地区黄檗阔叶混交林有25个阔叶树种,占比96.15%,黄檗的重要值最大;Levins和Shannon-Wiener生态位宽度指数大小排序基本一致,生态位宽度最大的树种均为黄檗,其次为胡桃楸;黄檗与枫桦、胡桃楸和裂叶榆等物种的生态位重叠值较大,生态位宽度和生态位重叠度表现出了明显的一致性。但是存在部分物种生态位宽度和生态位重叠度具有相反性,与生态宽度受到物种生态习性、生境特征和空间分布等因素有关。
【参 考 文 献】
[1]张光明,谢寿昌.生态位概念演变与展望[J].生态学杂志,1997,16(6):46-51.
ZHANG G M, XIE S C. Development of niche concept and its perspectives: A review[J]. Chinese Journal of Ecology, 1997, 16(6):46-51.
[2]彭文俊,王晓鸣.生态位概念和内涵的发展及其在生态学中的定位[J].应用生态学报,2016,27(1):327-334.
PENG W J, WANG X M. Concept and connotation development of niche and its ecological orientation[J]. Chinese Journal of Applied Ecology, 2016, 27(1): 327-334.
[3]白欢欢.黄山松群落主要木本植物营养生态位和种间联结特征研究[D].北京:中国林业科学研究院,2018.
BAI H H. Multi-dimension nutrition ecological niche and interspecific association characteristics research of dominant wood plants of Pinus taiwanensis communities[D]. Beijing: Chinese Academy of Forestry, 2018.
[4]高浩杰,袁佳,高平仕.舟山群島次生林优势种群的生态位与种间联结[J].热带亚热带植物学报,2017,25(1):73-80.
GAO H J, YUAN J, GAO P S. Niche and interspecific association of dominant species in secondary forest of Zhoushan islands[J]. Journal of Tropical and Subtropical Botany, 2017, 25(1): 73-80.
[5]吴倩楠,董建文,郑宇,等.百里杜鹃国家森林公园优势种生态位研究[J].南京林业大学学报(自然科学版),2017,41(2):175-180.
WU Q N, DONG J W, ZHENG Y, et al. Niches of the main plant species in Baili Rhododendron National Forest Park[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2017, 41(2): 175-180.
[6]潘高,张合平,潘登.湖南青羊湖国家森林公园针阔混交林主要种群生态位特征[J].中南林业科技大学学报,2017,37(2):69-75.
PAN G, ZHANG H P, PAN D. Niche characteristics of dominant populations within mixed broadleaf-conifer forest in Qingyanghu National Park[J]. Journal of Central South University of Forestry & Technology, 2017, 37(2): 69-75.
[7]陈俊华,刘兴良,何飞,等.卧龙巴朗山川滇高山栎灌丛主要木本植物种群生态位特征[J].林业科学,2010,46(3):23-28.
CHEN J H, LIU X L, HE F, et al. Niche characteristics of dominant woody populations in Quercus aquifoliodes shrub community in Balangshan Mountain in Wolong Natural Reserve[J]. Scientia Silvae Sinicae, 2010, 46(3): 23-28.
[8]庞久帅,原阳晨,周苗苗,等.槭叶铁线莲(Clematis acerifolia Maxim.)群落物种生态位研究[J].生态学报,2022,42(8):3449-3457.
PANG J S, YUAN Y C, ZHOU M M, et al. Species niche of Clematis acerifolia Maxim. community[J]. Acta Ecologica Sinica, 2022, 42(8): 3449-3457.
[9]俞昀,白小军,王志一.大兴安岭次生林区不同龄级落叶松(Larix gmelinii)生态位特征和竞争关系[J].生态学报,2022,42(12):4912-4921.
YU Y, BAI X J, WANG Z Y. Niche characteristics and competitive relationship of Larix gmelinii between different age classes in Greater Khingan Range secondary forest area[J]. Acta Ecologica Sinica, 2022, 42(12): 4912-4921.
[10]INGRAM T , COSTA-PEREIRA R, ARAUJO M S. The dimensionality of individual niche variation[J]. Ecology, 2018, 99(3): 536-549.
[11]梁玉莲,国庆喜.基于地形汇流值的东北天然次生林主要树种的生态位[J].生态学报,2010,30(11):2874-2882.
LIANG Y L, GUO Q X. Determining niches of the major tree species based on a flow accumulation technique in a natural secondary forest in Northeastern China[J]. Acta Ecologica Sinica, 2010, 30(11): 2874-2882.
[12]简尊吉,马凡强,郭泉水,等.三峡水库峡谷地貌区消落带优势植物种群生态位[J].生态学杂志,2017,36(2):328-334.
JIAN Z J, MA F Q, GUO Q S, et al. Niche of dominant populations in the water level fluctuation zone of canyon landform area of the Three Gorge Reservoir[J]. Chinese Journal of Ecology, 2017, 36(2): 328-334.
[13]趙家豪,叶钰倩,陈斌,等.江西武夷山南方铁杉针阔混交林主要植物生态位特征[J].林业科学,2021,57(1):191-199.
ZHAO J H, YE Y Q, CHEN B, et al. Niche characteristics of dominant plants of coniferous (Tsuga chinensis var. tchekiangensis) and broad-leaf mixed forest in Jiangxi Wuyishan, China[J]. Scientia Silvae Sinicae, 2021, 57(1): 191-199.
[14]赵阳,曹家豪,李波,等.洮河自然保护区大峪沟林区紫果云杉群落木本植物种群生态位特征[J].生态学报,2022,42(5):1865-1875.
ZHAO Y, CAO J H, LI B, et al. Niche of woody plant populations of Picea purpurea community in Dayugou forest area, Taohe Nature Reserve, Gansu Province[J]. Acta Ecologica Sinica, 2022, 42(5): 1865-1875.
[15]张仕艳,谢强,黄丽娟,等.广西灵渠流域青冈栎群落主要种群生态位分析[J].广西师范大学学报(自然科学版),2021,39(6):162-173.
ZHANG S Y, XIE Q, HUANG L J, et al. Niche analysis of main populations in Cyclobalanopsis glauca community in Lingqu Basin of Guangxi, China[J]. Journal of Guangxi Normal University (Natural Science Edition), 2021, 39(6): 162-173.
[16]王育松,上官铁梁.关于重要值计算方法的若干问题[J].山西大学学报(自然科学版),2010,33(2):312-316.
WANG Y S, SHANGGUAN T L. Discussion on calculating method of important values[J]. Journal of Shanxi University (Nat. Sci. Ed.), 2010, 33(2): 312-316.
[17]陈霞,袁在翔,金雪梅,等.紫金山针阔混交林主要植物种群生态位特征[J].中南林业科技大学学报,2020,40(8):113-119.
CHEN X, YUAN Z X, JIN X M, et al. Niche characteristic of main plant populations in a coniferous and broad-leaved mixed forest on Zijin mountain[J]. Journal of Central South University of Forestry & Technology, 2020, 40(8): 113-119.
[18]LEVINS R. Ordering the phenomena of ecology: evolution in changing environments[J]. Science, 1970, 167(3924): 1478-1480.
[19]COLWELL R K, FUTUYMA D J. On the measurement of niche breadth and overlap[J]. Ecology, 1971, 52(4): 567-576.
[20]姚慧芳,盧杰,屈兴乐,等.藏东南华山松林下灌草主要植物的生态位特征[J].林业资源管理,2021(6):69-75.
YAO H F, LU J, QU X L, et al. Niche characteristics of main shrub and grass plants under Pinus armandii forest in Southeast Tibet[J]. Forest Resources Management, 2021(6): 69-75.
[21]杨晓惠,林文树,刘曦,等.小兴安岭典型阔叶红松林幼树的群落结构动态研究[J].中南林业科技大学学报,2021,41(12):87-97.
YANG X H, LIN W S, LIU X, et al. The dynamics of sapling community structure of typical broad-leaved Korean pine forest in the Lesser Khingan mountains[J]. Journal of Central South University of Forestry & Technology, 2021, 41(12):87-97.
[22]陈玉凯,杨琦,莫燕妮,等.海南岛霸王岭国家重点保护植物的生态位研究[J].植物生态学报,2014,38(6):576-584.
CHEN Y K, YANG Q, MO Y N, et al. A study on the niches of the state's key protected plants in Bawangling, Hainan Island[J]. Chinese Journal of Plant Ecology, 2014, 38(6): 576-584.
[23]杨新东,陈流保,曾献兴,等.广东翁源青云山自然保护区森林群落优势树种的生态位特征[J].热带亚热带植物学报,2020,28(6):565-573.
YANG X D, CHEN L B, ZENG X X, et al. Niche characteristics of dominant species of forest community in Qingyunshan Natural Reserve, Wengyuan, Guangdong Province[J]. Journal of Tropical and Subtropical Botany, 2020, 28(6): 565-573.
[24]刘加珍,陈亚宁,张元明.塔里木河中游植物种群在四种环境梯度上的生态位特征[J].应用生态学报,2004,15(4):549-555.
LIU J Z, CHEN Y N, ZHANG Y M. Niche characteristics of plants on four environmental gradients in middle reaches of Tarim River[J]. Chinese Journal of Applied Ecology, 2004, 15(4): 549-555.
[25]董冬,许小天,周志翔,等.安徽九华山风景区古树群落主要种群生态位的动态变化[J].生态学杂志,2019,38(5):1292-1304.
DONG D, XU X T, ZHOU Z X, et al. Niche dynamics of main populations of old-tree communities in Jiuhua Mountain Scenic area of Anhui Province[J]. Chinese Journal of Ecology, 2019, 38(5): 1292-1304.
[26]周梅妹,高纯,苑景淇,等.山楂海棠种群分布格局与生态位特征[J].北华大学学报(自然科学版),2021,22(3):299-307.
ZHOU M M, GAO C, YUAN J Q, et al. Population distribution pattern and niche characteristics of Malus komarovii (Sarg.) Rehd.[J]. Journal of Beihua University (Natural Science), 2021, 22(3): 299-307.
[27]ROBERT M, CHRISTOPH N, WILHELM B, et al. Biodiversity and endemism mapping as a tool for regional conservation planning case study of the Pleurothallidinae (Orchidaceae) of the Andean rain forests in Bolivia [J]. Biodiversity and Conservation, 2003, 12: 2005-2024.
[28]何忠伟,胡仁传,黄日波,等.广西银杉林主要树种种群生态位分析[J].林业科学研究,2012,25(6):761-766.
HE Z W, HU R C, HUANG R B, et al. Niche of the major tree species populations in Cathaya argyrophylla forest in Guangxi[J]. Forest Research, 2012, 25(6): 761-766.
猜你喜欢
蜜蜂杂志(2022年8期)2022-09-14
北华大学学报(自然科学版)(2021年2期)2021-05-07
传奇·传记文学选刊(2020年12期)2020-12-23
防护林科技(2019年1期)2019-03-13
藏天下(2018年7期)2018-08-20
中老年健康(2017年11期)2017-12-13
文化学刊(2017年9期)2017-03-11
中老年健康(2016年11期)2017-02-05
现代家庭(2016年9期)2016-09-12
中国林副特产(2013年4期)2013-08-07
