
光质调控对豆科和禾本科牧草生长影响的研究进展
2023-07-31 05:36:42李怡彤李羽超祁兆奔李润谢菲刘权
中国农学通报 2023年16期
李怡彤,李羽超,祁兆奔,李润,谢菲,刘权
(1兰州大学草地农业科技学院/红三叶高值化开发利用创新创业示范基地,兰州 730020;2阿拉善盟林业草原和种苗工作站,内蒙古阿拉善盟 750300)
0 引言
光是植物生长发育的基本环境因素,是植物进行光合作用的直接能量来源,调控植物生长。太阳光谱可以粗分为紫外辐射、可见光或光合有效辐射和红外辐射(又称远红光等)3 个部分(图1)。植物的生长发育不仅受光量或光强(光子通量密度,photonflux density,PFD)、光周期等的制约,而且受光质即波长不同的光与辐射及它们不同组成比例的影响[1]。不同光质的辐射对植物有不同的生物学效应,对植物光合作用、光形态建成、器官生长发育、生理学特性、营养成分组成产生影响。

图1 太阳光谱波长分布示意图
在蔬菜作物生产方面光调控技术已经得到大量应用。有研究表明,增加蓝光可以显著提高温室内苋菜叶片的光合能力,从而提高其产量[2]。此外,不同比例组合的光质对作物的影响也不同。研究表明,生菜采前连续红蓝光照射中红光比例越高,地上部和根的鲜重及干重越高[3],复合光比单色光更能促进生菜生物量积累[4]。光强为100 μmol/(m2·s)且红蓝光比例为5:1的光照条件对辣椒幼苗的生长和光合作用最有利[5]。光质调控还会抑制农作物的生长发育,周成波等[6]研究发现,对叶用莴苣补充比例为1:1 的红蓝光不利于其地上部分干物质积累与光能利用。
目前关于光调控对于牧草生长影响的研究相对较少(图2),且年份跨度大,缺少系统性结论观点,光调控生长的机制尚不明确。随着人们物质生活水平的提高,膳食结构发生了显著变化,肉蛋奶产品的刚性需求量上涨;1982—2018年中国居民人均畜产品消费量处于快速增长状态,导致肉蛋奶缺口进一步增加[7-9]。因此,养殖业得到了空前发展,这造成了饲草短缺、过度放牧和草场破坏等问题,不合理的草原利用以及全球气候的频繁变化,国内草原品质及生态环境出现了严重退化[10],导致畜牧业存在饲料粮短缺、农田土壤退化、毒化严重、奶制品产量低质量差等问题[11]。此外,由于国内牧草种子和饲草产品存在生产水平低、质量差、机械配套不足等问题,饲草生产尚不能满足高品质畜牧业产品的供给需求。中国牧草资源种类繁多,但饲喂以禾本科和豆科牧草为主[12]。通过光质调节提高牧草生物产量,提升牧草品质,有助于缓解饲草缺口、人畜争粮等问题。
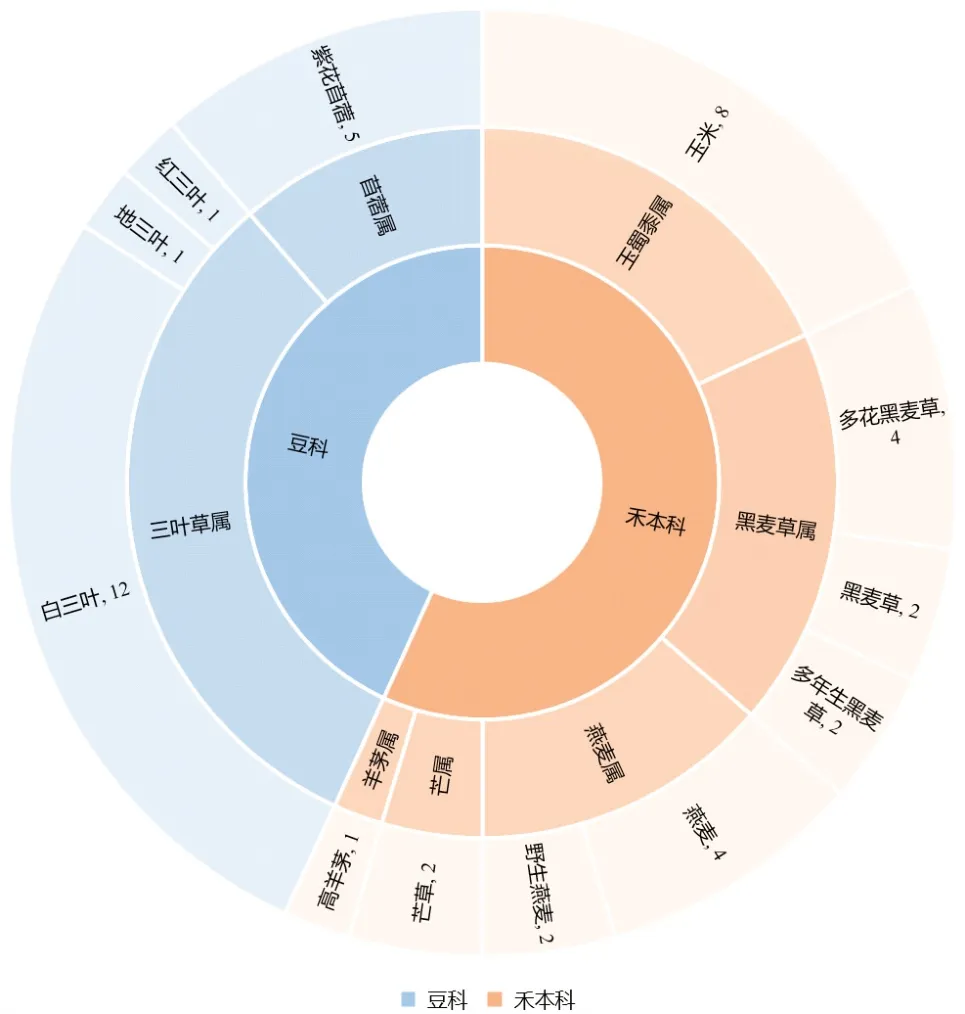
图2 1983—2022年豆科、禾本科牧草光质调控相关文献统计
关于光质调控对牧草的影响主要集中于红光、蓝光、紫外光和远红光,其中以豆科、禾本科牧草的研究居多。笔者综述光调控对禾本科(黑麦草属、羊茅属、燕麦属、芒属、玉蜀黍属)和豆科(三叶草属、苜蓿属)的研究现状,以期为牧草高质量生产管理和应用开发提供参考。
1 光质对牧草生长发育的影响
1.1 对生长发育的影响
不同光质对牧草器官根、茎、叶发育的促进作用不同。豆科牧草白三叶在蓝、绿、红荧光灯分别照射下,叶面积在蓝光下增长最快,绿光下最慢;叶柄在红光下最长,绿光下最短;茎根长度比在红光下最大,蓝光下最小[13]。光照强度的改变也会影响其促进作用。0.02 μmol/(m2·s)光照强度蓝光下的白三叶较83 μmol/(m2·s)蓝光的,叶柄长度、叶柄与水平面的角度及冠层上层的叶片高度增加,主轴叶柄的叶面积和生物量也有增加。而对于禾本科牧草,低光照强度21 μmol/(m2·s)的蓝光照射下黑麦草和高羊茅叶伸长率和叶伸长时间增加,叶片长度增长[14-16]。
匍匐茎的生长与匍匐茎的延伸、分枝率有关,其对环境中光条件改变会产生塑性反应,尤其是蓝光的调控,在豆科牧草白三叶上的研究颇多。Gautier 等[17]研究发现,低光照强度的蓝光可以提升白三叶地面以上匍匐茎的离地高度,促进白三叶在水平方向的扩展。此外,对于不同级别的植物轴,照射低强度的蓝光可以触发它们的差异反应。0.02 μmol/(m2·s)低光强的蓝光照射在主轴上较83 μmol/(m2·s)高光强的蓝光对节间长度或匍匐茎长度没有影响;但照射在较老的一级枝上,表现为低光强照射下匍匐茎的长度减少;在二级枝上影响更明显,导致匍匐茎长度显著减少,对植物的横向延伸有抑制性[18]。同时,38 μmol/(m2·s)低光强的蓝光较83 μmol/(m2·s)光强的蓝光可以增加白三叶个体发育的相对速率,主轴上叶片出现率增加,平均叶柄长度增加,但总叶面积减少[19]。而远红光主要通过改变叶片形态来改善三叶草叶片对光的捕获,相对较低光照强度下发育的叶片比较高光强度下发育的叶片叶面积更大,叶柄长度更长。研究表明远红光处理能增加白三叶的茎根比,减少植株匍匐茎的分枝,促进叶柄和节间的伸展[20-21]。而红光照射与自然光相比,可以促进黑麦草总生物量的快速累积,利于其地下部分根系的生长[22]。对于地三叶而言,Aud 等[23]指出白光或红光可以加重对其光周期的负面影响,调控活性吸收光敏色素形式Pfr,参与促进臭氧引起的可见叶面损伤。
红光、远红光对种子发芽起到一定的调节作用。远红光对种子发芽有抑制作用,而红光对种子发芽有促进作用[24]。研究表明红光和远红光照射下,野生燕麦颖果胚胎盾片组织的吸水率存在差异,造成休眠种子和非休眠种子萌发情况不同。与远红光处理相比,红光处理后18 h检测到胚壳组织水分含量显著增加,表明红光对种子发芽的刺激比远红光更强[25]。休眠种子在光照或黑暗条件下水合时不会发芽,但在黑暗中完全水合储存(以下称为“种子分层”)2~3周会使得种子对光敏感,使它们处于休眠状态,并在之后暴露于光时发芽。研究表明远红光可以促进一年生黑麦草种子分层期间的休眠释放,程度与黑暗相似,红光在抑制休眠释放方面与白光相似[26]。
不同比例的光质组合对于植物器官的生长发育也存在不同的影响,其中红光-远红光(R-FR)组合是影响植物生长发育不可缺少的混合光质。白三叶在低R:FR光照处理下叶面积会增加[21],且两者光质比例适当下调会增加匍匐茎的分枝量,从而促进生长。研究表明在光强为175 μmol/(m2·s)R:FR(1.12)光照处理下,对白三叶主匍匐茎分枝减少不显著[27],而低R:FR(0.27)光照处理下会发生植物生长延迟,最终导致分枝量较少[28],更低的R:FR(0.25)光照处理会导致植株每个主匍匐茎的主枝数变得更少[29]。黑麦草分蘖延伸和叶鞘长度随着远红光的增加而增加,在光强为107 μmol/(m2·s)R:FR(0.84)光照处理促进了多花黑麦草的生长发育,并增加了每株可育分蘖数,还增长了叶鞘、叶片和生殖芽,增强了节间伸展生长[30-31]。
1.2 对营养物质的影响
植物利用光合作用产生营养物质,但每个物种对光的需求略有不同,不同光质产生的营养物质响应也存在显著差异。对于豆科牧草白三叶而言,蓝光处理下的总植物干重、根瘤干重和总固氮累积率最大,而在绿光下最少[13]。苜蓿则是在RGB 光(蓝绿红光)下芽苗产量最高,同时总酚类化合物含量最高[32]。而对于禾本科燕麦幼苗来说,具有较高光强度和较高蓝/红光比的光源(157 μmol/(m2·s)光强和1.27 蓝/红光比的Spectrolight-2 LED 照明器)最适合其营养物质的积累[33]。
单色光处理有利于提高植株的营养品质。冯娜娜[34]在不同光质照射豆科牧草紫花苜蓿的试验中发现,白光、蓝光、红光与绿光及黑暗环境相比,可提高幼苗的品质及抗氧化能力,其中红光可以显著提高苜蓿的可溶性糖含量。红光还可以增加燕麦抗坏血酸含量[35]。蓝光可以显著提高苜蓿的可溶性蛋白、花青素含量,从而提高DPPH 自由基清除活性及抗氧化能力[34];(30±3)μmol/(m2·s)光强的蓝光还可以增加苜蓿芽苗菜中黄酮类化合物、总酚、游离氨基酸及抗坏血酸含量[36],14 μmol/(m2·s)光强的蓝光能有效增加燕麦中抗坏血酸含量[37]。此外,白光也能提高苜蓿中总酚和黄酮类化合物含量[34],(30±3)μmol/(m2·s)光强的白光可以增加苜蓿芽苗菜干物质含量,黄光处理可以显著提高芽苗菜槲皮素含量和苯丙氨酸解氨酶活性[36]。UV-B辐射与其他光质处理相比,更利于苜蓿的营养品质和抗氧化性的提高,体现在其能够有效提高可溶性糖含量,提高可溶性蛋白含量、总酚、花青素、黄酮类化合物等紫外吸收物质含量[34]。
光质与光照强度对植株的营养生长的影响是相辅相成的。研究表明低剂量的UV-B光照可促进植物合成多种紫外线可吸收的化合物,激活多种酶的催化活性,上调核基因表达,对植物生长发育有积极的生物学效应[37]。王小飞[38]通过试验证明了该结论,表明UV-B照射下紫花苜蓿幼苗组织中山奈酚、异甘草素、芹菜素和大豆苷元等黄酮类化合物的含量均明显升高,且在较低水平UV-B照射下黄酮类化合物的含量明显高于较高水平UV-B 照射下的含量。与之结论相反,研究表明1.6 倍UV-B 辐射比1 倍UV-B 能诱导白三叶更多的黄酮类化合物产生[39],且UV-B辐射的增加同样导致饲用玉米叶片中黄酮类化合物的积累,赖氨酸水平上升,而种子的蛋白质、糖和淀粉水平则降低[40]。燕麦叶片在蓝光下比在白光照射下幼苗的生长受到的抑制效果更显著,降低生物量40%。同时也会导致叶片乙烯合成差异,蓝光照射燕麦叶片后,1-氨基环丙烷-1-羧酸(ACC)氧化酶活性较黑暗处理降低,从而导致ACC 依赖性乙烯含量降低。研究表明20 μmol/(m2·s)蓝光和红光较白光和黑暗处理对乙烯的生成抑制效果明显,而2 μmol/(m2·s)远红光和1.5 μmol/(m2·s)UV 可以有效刺激ACC转化为乙烯[41]。此外,远红光照射下的燕麦幼苗与黑暗条件下的幼苗相比,表现出不同的生长和向地性响应,880 nm的远红光照射下幼苗表现出更明显的垂直生长趋势,随着辐射波长接近可见光,重力性响应略有增加[42]。
光诱导离子通量的响应与光质相关。光诱导的K+通量动力学在饲用玉米生长(完整叶片)和非生长组织(叶肉组织)之间是不同的。研究表明,与红光相比,蓝光在饲用玉米叶片刺激生长的基底区域吸收K+方面要更多;而与蓝光相比,红光在叶片生长过程中Ca2+信号传导中十分重要[43]。
2 光质对牧草光合作用的影响
光质对牧草叶片光合作用的调控是一个复杂的分子调控过程,其中包括光形态的建成,光信号的感知、接受和转导,光控基因的表达,光系统的能量转换等机制层面调控。此外,参与光合作用的酶和主要色素会对不同光质产生一定的响应,光合参数也会发生相应的变化。
2.1 对光合作用调控机制的影响
植物对光环境极为敏感,光质的改变会引起生物体内的一系列细胞分化、结构和功能变化并最终影响组织和器官形成的过程[44],包括种子的休眠和发芽、开花结果及生长方向(向光性)等[45]。例如,低剂量的UV-B辐射可以促进植物的光形态建成[37],而高剂量的UV-B光则会造成植物体光合系统的损伤,如光合色素合成受阻、类囊体完整性改变、Rubisco酶活性降低,破坏细胞结构和功能等[46]。
2.1.1 对光信号的接受和转导的影响植物对光质作出响应的前提是感知和传递不同光质的信号。植物光信号通路涉及级联反应,包括多种光感受器与信号转导基因或转录因子的共同参与,它们之间相互作用共同影响植物生长发育[47-48]。研究已发现的光信号接受体有光敏色素(红光/远红光受体)、隐花色素和向光素(蓝光受体)、UVR8蛋白(紫外光受体)等。
光敏色素(PHY)是植物感受红光和远红光的光感受器[49],可调节种子萌发、幼苗光形态发生、避荫和叶片衰老等过程[50]。研究表明,野生燕麦种子处于休眠状态时,对光没有反应,而在种子发育的第Ⅱ阶段吸水过程,红光远红光混合处理后的种子发芽,证实光敏色素介导参与种子的吸水过程[24]。隐花色素(CRY)是主要感受蓝光的光受体蛋白。有研究表明蓝光激活的GmCRY2a隐花色素基因通过抑制GmCIB1与GmWRKY53启动子区的结合,下调GmWRKY53基因的表达,从而延迟大豆叶片衰老[51]。1995 年Liscum 和Briggs[50]从拟南芥的突变体nphl 中首次分离得到了调控向光性的蛋白NPH1。在蓝光处理下,向光蛋白可形成共价蛋白-黄素单核苷酸,启动光信号的传导[51]。UVR8 蛋白为紫外光受体,王小飞[38]构建紫花苜蓿UVR8 超表达载体,并通过转基因烟草表达的方法表明,经UV-B 光辐射后,UVR8 蛋白向核内转移,证明UVR8蛋白参与紫外光调控,但具体机制仍不明确。
2.1.2 对光合基因的表达和光系统的影响植物可以通过调整光合器官的结构来增强对光的吸收和利用[53],适应其光合生理反应。光质可以通过影响气孔指数和密度[54-55]来影响植物进行光合作用。例如,通过光质调控饲用玉米和芒草的气孔基因表达可以增加叶片气孔密度,显著提高光合速率[54]。研究表明,小麦幼苗在双光束试验中以红色光为背景照射低强度蓝光,可以恢复对气孔导度的敏感性响应,诱导气孔打开[56]。
饲用玉米作为一种对光高度敏感的C4植物,其光合基因-气孔因子的表达受到光照的调节。光诱导的气孔发育是通过隐花色素(CRY)、光敏色素(PHY)和组成型光形态1(CRY-PHY-COP1)信号系统之间的串扰来调节的[57]。红光和蓝光都控制着气孔的发育,蓝光抑制CRYs的表达并通过控制COP1 和CRY直接调节气孔形成,而红光作为CRY和PHY 的激活剂,通过细胞内和细胞间信号传导过程调节气孔发育。红光反应与保卫细胞中渗透势的变化相关,保卫细胞电子传递产生ATP 为糖酵解和线粒体呼吸产生ATP 提供额外能量,可用于渗透调节的过程,响应红光和蓝光诱导的气孔开放[58]。
ZmCOP1是光形态发生、开花、气孔开放的关键负调控因子[57],而ZmSTOMAGEN是近年在玉米中发现的气孔发育正调控基因,研究表明蓝光可以促进玉米ZmSTOMAGEN基因的表达,促进气孔蛋白的表达和无机元素的吸收,提高气孔密度和气孔指数,有助于植物进行光合作用[59],而相对应的蓝光减少,红光增加会对ZmCOP1进行正向调节,促进其表达,抑制气孔功能。红光在玉米幼苗气孔形成过程中主要参与并正向调节基因功能,在气孔发育过程中蓝光占主导作用[60]。
光能转换为化学能的反应是由位于类囊体膜上的光系统I(PSI)和光系统II(PSII)协同作用而完成的。由于两者结合的色素组成不同,不同波长的光对2 个光系统的激发常不均衡[61]。Kenneth等[62]研究表明,与白光和红光照射相比,远红光加上低红光抑制了PSI(CP1+CP1a)反应中心蛋白的形成并增强了束鞘和叶肉细胞中的PSII 蛋白合成。同时,(110±40)μmol/(m2·s)的远红光可以降低PSⅡ的光化学量子产率;红光远红光混合下,低R:FR(0.4)可以使白三叶叶片的PSⅡ中心光化学量子产率显著降低[63];红光处理导致玉米叶绿体中PSⅡ组分的Deg1 依赖性裂解,而远红光不诱导Deg1介导的降解[64]。此外,Kenneth等[65]研究表明红光比蓝光对玉米PSⅡ相关的叶绿素a、b 和光捕获蛋白(LHC-2)的合成更有促进作用;而蓝光对PSⅠ相关的捕光蛋白(LHC-1)的合成更有效,红光中添加少量蓝光会抑制红光对LHC-2蛋白合成的促进作用,也会造成mRNA的平衡水平降低[66]。Romualdo等[67]通过测量燕麦植株叶片切片发出的叶绿素荧光发现,与适应黑暗的叶片相比,单独白光和红光降低了叶片发出的最大荧光。单独绿光或蓝光不会改变叶片发出的最大荧光。此外,研究表明,蓝光和绿光以一种独立于远红光的方式介导植物休眠维持。绿光和蓝光多种感受器参与一年生黑麦草种子休眠的维持,其中没有光敏色素的参与[68]。
2.2 对叶片光合色素含量的影响
高等植物的光合色素分为叶绿素和类胡萝卜素2类。光合色素吸收、传递和转化光能,是植物进行光合作用的基础,其含量与组成影响植物叶片光合速率,是评价植物生长发育状况和光合作用能力的重要指标[69]。
光合色素的形成受到光质的调控作用,不同牧草的光合色素含量及组成对不同光质的响应规律存在一定差异。唐丽等[36]证明30 μmol/(m2·s)白光处理显著提高了苜蓿芽苗菜类胡萝卜素含量;而苜蓿子叶在21.6 μmol/(m2·s)的RGB 光和冷白光处理下叶绿素和类胡萝卜素含量达到最高[32]。Gałązka-Czarnecka 等[70]通过不同光源比较发现,UV-A 和蓝光是最能促进红三叶芽中类胡萝卜素含量提高的光源。此外,光质也会对光合色素的产生起到抑制作用。UV-B 辐射条件下,紫花苜蓿幼苗叶片和黑麦草叶片的光合色素合成受到明显抑制[38,66],并且随辐射的增加,光合色素含量降低的幅度更大[40]。低比例的红光远红光(0.4)混合照射处理使白三叶刚展开的新叶叶绿素含量降低[63],成熟期叶片并没有改变,对总生物量积累和干物质在不同器官分配方面没有影响。
2.3 对叶片中酶活性、光合参数的影响
植物的各项生理活动都离不开酶的参与和调节。研究表明,蓝光可以提高苜蓿苯丙氨酸解氨酶活性,促进苯丙烷代谢,促进植物次生代谢物质的生物合成[34]。而Shang 等[55]研究表明红光可以导致黑麦草叶片光氧化反应中的过氧化氢酶失活。此外,光质对植物净光合速率、气孔导度、胞间CO2浓度、蒸腾速率、CO2同化率等光合参数的变化中,也少不了酶的参与。Sun等[54,71]通过蓝光、红光、绿光处理,发现芒草和玉米在蓝光下的光合速率、CO2同化的量子产率和最大磷酸烯醇丙酮酸羧化酶活性均显著低于白光、红光和绿光,蓝光下的能量限制影响了C4和C3循环,导致净CO2同化率最低。红光蓝光混合处理比单独红蓝光处理能表现出更高的净光合速率、气孔导度、胞间CO2浓度、蒸腾速率等,在150 μmol/(m2·s)的红光蓝光各处理4 d 的情况下,玉米幼苗表现出最高的气体交换能力,较红光和蓝光单独处理8 d效果更佳[72]。
3 光质调控技术的研究前沿
相对而言,果蔬、园艺作物等的光质调控技术应用更为全面和成熟,而在牧草方面的应用与研究局限于产量、品质及营养物质层面,光质对光合作用调控机制方面研究较为缺乏。
近些年来,光质调控技术在果蔬、园艺作物等的应用愈加关注功能性次级代谢物的积累[73-74]。研究表明,紫外线可以提高蔬菜次生代谢物含量[75],同时,紫外线和蓝光影响水果果实花青素的生物合成调控[76]。不同光谱波段还可以向园艺植物的光感受器传递特定信号,从而诱导与生理和代谢功能相关基因的表达[77]。例如果蔬生长过程中,关注光质对红肉桃[78-79]、红葡萄[80]、血橙[81]等果肉果皮色泽的调控,对黄瓜[82]嫁接苗愈合生长的调控,对辣椒[83]丛枝菌根共生及磷肥吸收的调控,对银耳子[84]、香菇[85]等农艺性状的影响,与栽培技术相结合产出符合人们需求的高质果蔬。此外,具有观赏性的园艺植物通过光质调控可以实现更快开花及着色[86]。对光质的调整也已运用于提高药用植物性能的方面,获得最大的次级代谢物产量,从而服务于人类健康[87]。
在设施应用方面,光调控技术在果蔬及园艺作物种植领域已广泛运用于受控环境农业、多层垂直农业及果蔬采后保鲜等诸多方面[88],采取温室大棚高架进行多方向补光处理,根据品种类型进行光源布置和光质光强调控,实现温室增产[89]。研究表明,在番茄的栽培中,计算机功能结构模型的建立可以量化植物结构对植物光吸收和光合作用的影响,可有效模拟植物之间的相互作用及光的三维分布[90],模拟光质调控的作用机制,节省大量的人力物力,为更好地探究最佳光质组合提供新的可能。同时,随着LED 技术的精进,对光质光量控制趋于精细化,通过精准控制红蓝光百分比已实现对生菜、菠菜、羽衣甘蓝、罗勒和甜椒的生长特性、色素含量和抗氧化能力的调控[91],还可通过调控不同时期或照射不同部位的光质提高果蔬品质。
此外,计算机模拟及分析技术的应用提高了对光质调控机理的认识和理解。通过电子层析成像技术分析的低分辨率数据与单个蛋白质复合体和超复合体的高分辨率信息,结合计算机模拟可以揭示堆叠的类囊体细胞结构域中的光系统II(PSII)和复合体的蛋白质网络建立光捕获的机制[92],为全面了解光质如何影响叶绿体不同结构层次(全膜、介观、单一蛋白质)连接、相互作用和共同作用奠定了基础[93]。除研究成熟的叶绿体外,其余独特细胞器的生物过程发生和降解也是新的研究目标。
4 研究展望
植物对光质的响应是一个复杂的过程,种属差异、生长差异使植物对各种光质的响应不同。就牧草生产而言,牧草种类和品种对光质需求具有差异性,同时生长发育阶段对光质需求也具有明显的阶段性。例如种子萌发时期需要大量的水分补充,红光调控可以参与到种子的吸水过程,促进种子发育[24-25];幼苗时期,光质的不同组合尤其是红蓝光的组合有利于营养物质的积累,提升牧草品质和产量,远红光、红光和蓝光的调控可以在气孔基因层面促进牧草进行光合作用。但整体而言,豆科禾本科牧草对同一种光质的响应具有相似性,研究结果具有一致性。例如蓝光可以促进白三叶和黑麦草的叶片生长[13-16],红光和远红光的组合可以促进它们两者的分蘖生长[27-31]。
通过文献梳理发现,国内外目前针对光质对牧草光形态建成和生长发育调控存在研究内容较少、年限长、研究范围局限、光质类型少、机制机理尚不明确等问题。对于许多牧草作物来说,不同物种在每个物候期为优化产量和产品质量所需的最佳光谱和强度仍然未知。建议在今后的研究中,在如下方面进行深入研究:(1)针对性提升某些功能性代谢产物的开发利用水平,提高对农艺性状的调控,利于牧草营养物质积累、资源优化利用;(2)光调控技术不应局限于光强、光质及光周期的调节,应与其他调控措施(温度、气体和水肥)开展耦合研究,寻求提高牧草品质和产量的最佳农艺措施组合,尽最大可能提高光合效率,提升利用效率;(3)人工补光光源类型种类更加丰富的同时,设施应更加先进、精准;(4)加强机制机理研究,利用多组学方法,如基因组学、蛋白质组学、代谢组学和结构生物学相结合,对常见牧草的光合作用分子调控机理及叶绿体内电子转移机制进行全面研究,为牧草生产提供理论基础;(5)结合信息通信技术、生物学及物联网最新科技,计算模拟调控模型,整合出有关牧草的大型生产系统,为增质增产、品种优化提供新的发展思路。
虽然目前光质调控还未大规模应用于牧草生产中,存在一定的差距,但相信通过开展光质调控牧草光合作用代谢过程机理的深入研究,开展耦合学科及多种因素综合处理,实现最新科技的利用及设施完善,将会为牧草有效成分的高效利用和高营养价值的快速累加提供强有力的理论支撑,为克服季节、温度、地域等自然因素的限制,实现器官发育、生理学特性及营养成分多种指标的定向提升奠定基础,同时也为更高水平的牧草工厂、品种选育铺平道路,对于保障食物安全也具有重要意义。
猜你喜欢
学与玩(2022年6期)2022-10-28 09:18:52
四川林业科技(2022年1期)2022-03-22 07:48:32
上海农业学报(2022年1期)2022-03-08 00:09:12
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
——记嘉荫县红光乡燎原村党支部
奋斗(2020年24期)2021-01-16 06:23:36
装备制造技术(2019年12期)2019-12-25 03:06:52
科技创新与品牌(2019年12期)2019-02-06 04:03:29
浙江农业学报(2017年1期)2017-05-17 06:13:36
中国铸造装备与技术(2015年5期)2015-12-10 10:23:33
中国果业信息(2015年12期)2015-01-24 07:31:03
