
生物多样性导向的再生水补给型城市河道水生态修复效果
2023-07-29 13:55孟婷婷董月群闻丞薛晓飞冒建华黄越李丹汪洋徐开翔
环境工程技术学报 2023年4期
孟婷婷,董月群,闻丞,薛晓飞,冒建华,*,黄越,李丹,汪洋,徐开翔
1.北控水务(中国)投资有限公司
2.北京镜朗生态科技有限公司
3.清华大学环境学院
4.北京净通水环境治理有限公司
自2015年国务院颁布实施《水污染防治行动计划》(简称“水十条”)至今,我国全国范围内黑臭水体治理任务已基本完成,城市河道水环境质量得到显著提升,但仍面临返黑返臭的风险,有研究证明经生态修复的河道返黑返臭风险明显低于未经生态修复的河道[1]。而且,随着污水处理规模、水资源循环利用规模的提升,再生水补给的河道、湖泊逐渐增多。2021年北京市再生水利用量达12 亿m3,占北京年度水资源配置总量的近30%,使用总量居全国第一。但是再生水中相比于天然水较高的有机物、氮磷浓度及pH 增加了河道爆发藻类水华的风险[2]。
在截污等岸上工程均已完成的情况下,水生态修复成为解决以上问题的必要、有效措施。水生态修复的措施多样,常见的包括曝气复氧、生境修复、生态岸线改造、沉水植物种植等。工程中通常采用2 种或以上措施组合实施[3-7]。目前工程中所采取的生物群落构建措施往往以单纯的沉水植物种植为主,投放水生动物的工程基本以投放鲢、鳙、鲫、螺等物种为主,从而导致系统不够稳定、运行维护成本较高,较少有将水生植物、水生动物、浮游动物等生态系统全营养级恢复的案例。从工程效果持久性、运营维护难度上来看,只有完整的生态系统才能增强生态系统功能性和不良环境抵抗力,更好地发挥生态系统的营养物质吸收转移和净化能力,减轻后期运行维护压力[8]。另一方面,生物监测在水环境治理效果评估中也发挥越来越重要的作用。相比于传统的水体物理或化学指标的测定,生物群落监测不容易受到河道水流等瞬时因素的影响,且具有整合不同时间尺度上化学、物理和生物影响,能够反映多种生态胁迫对水环境的累积效应的优点[9-10]。利用水生生物群落结构、生物多样性指数监测等,可以直观评价工程效果,也可为后期水环境治理和运营提供预警和参考[3,5,11-14]。目前水生态监测多采用浮游动物、底栖动物多样性指数,少有各营养级生物、多指标全面监测评估的案例。
综上,如何在再生水补给型河道中构建包含食物链各营养级的自稳定生态系统,实现水生态快速恢复和生物多样性快速提升,并凭借系统自稳定性实现水质净化效果的持续,是当前水生态修复工程的重点方向和主要挑战。研究水生态修复工程后的水生态指标变化,也可以更好地评价水生态修复工程效果,为后续工程设计提供参考。
萧太后河是北京城市副中心文化旅游功能区的核心河道,串联起北京由西向东的南部城区。萧太后河也是北京市2017年重点整治的 53 条黑臭水体之一,经过综合治理后已于 2017年摘掉“黑臭水体”帽子。但由于北京环球度假区的建设,萧太后河老河道成为断头河,且由于部分补水水源的水质问题河道水质面临返黑返臭的风险。笔者选取北京城市副中心文化旅游功能区的核心河道——萧太后河老河道为研究区域,开展全营养级生态系统修复试验,以探索再生水补给型河道以生物多样性提升为导向的生态修复路径,并尝试根据水生态指标监测评价工程效果。
1 研究区与研究方法
1.1 研究区概况
研究区域位于萧太后河老河道因北京环球度假区建设截断后的断头处,面积约1 万m2(图1)。

图1 研究区域位置示意Fig.1 Position of the study area
本研究开展期间研究区域主要水源为位于西侧上游约500 m 处的生活污水处理站,该处理站设计处理规模800 m3/d,实地监测出水水质低于GB 3838—2002《地表水环境质量标准》[15]中的Ⅴ类水质标准限值,河道平均水位维持在 30~50 cm。上游处理站出水化学需氧量(COD)、总氮(TN)、氨氮(NH3-N)、总磷(TP)监测结果如表1 所示。
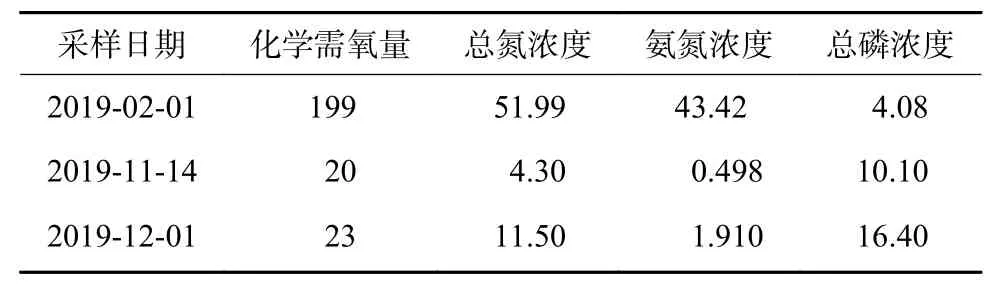
表1 研究区域上游污水处理站出水水质Table 1 Effluent quality of the upstream sewage treatment station of the study area mg/L
研究区域虽已消除黑臭但仍不满足北京城市副中心要求的Ⅳ类水质标准,更无法满足副中心文化旅游核心功能区的生态品质要求,且河道因为断头河而出现返黑返臭趋势。
2020年初开始,由于北京环球度假区外萧太后河上跨河桥梁的修建,南大沟来水被截断无法进入研究区域,上游生活污水处理站出水仅部分进入研究区域,导致2020年10 月—2021年5 月连续8 个月水位严重下降至平均不足15 cm,甚至出现部分河床裸露的情况。2020年10 月—2021年2 月持续5 个月上游生活污水处理站出水进一步恶化,导致研究区内水质持续恶化。因此,本研究区域内的生态修复,遭遇了水质、水量的双重考验。
1.2 生态修复试验
以提高水体生物多样性为导向,结合经典生物操纵,试验区域于2020年7 月31 日—10 月15 日开展水生态修复。具体措施包括在试验布置区(图1)设置里外2 道透水性围网,形成物种修复种源区。首先在其中构建水生植物群落,种植轮叶黑藻( Hydrilla verticillata) 、 金鱼藻(Ceratophyllumdemersum)、穗状狐尾藻(Myriophyllum spicatum);待水生植物系统稳定后,投放浮游动物和底栖动物,浮游动物物种包括隆线溞(Daphnia carinataking)和仙达溞(Limnosida frontosasars),底栖动物包括铜锈环棱螺(Bellamya aeruginosa) 、 中华圆田螺(Cipangopaludina cathayensis)、背角无齿蚌(Anodonta woodianawoodiana)和黑壳虾(Atyoidaesp.);待上述系统稳定后,分批投放鱼类,包括中小型功能鱼类草金鱼(Carassius auratus) 、 餐条(Hemicculter Leuciclus)、鳑鲏(Rhodeinaesp.)和大型食肉鱼类黑鱼(Ophiocephalus argus)、鲶(Silurus asotus)。浮游动物、底栖动物和鱼类均选择幼体种苗,避免使用成体个体。物种投放后定期监测,避免物种大规模流失。本次试验过程中未发生该类情况。
1.3 水质监测与生物调查
1.3.1 水质监测
在图1 试验布置区及上游污水处理站出水口处分别设立水质监测点。2020年7 月—2021年3 月及2021年8 月每月进行1 次水质监测,监测指标包括城市黑臭水体指标氨氮、溶解氧(DO)、氧化还原电位及地表水常规水质指标总磷、总氮、化学需氧量。
1.3.2 水生态调查
对浮游植物、浮游动物、大型底栖动物(不含水生昆虫)、鱼类、鸟类开展监测,方法如表2 所示。对浮游植物、浮游动物、大型底栖动物在现场取样后将样品带回室内开展物种鉴定及密度检测工作。鱼类、鸟类开展现场物种鉴定和相对丰度估算。

表2 生物调查方法Table 2 Methods of biological investigation
1.4 数据处理与方法
数据分析及相关图表制作在Excel 软件中完成。
通过计算藻类、浮游动物、底栖动物、鱼类和鸟类的物种丰度(S),即某一群落包含的物种种类数以及底栖动物、鱼类和鸟类的优势度(Y)、Shannon-Wiener 多样性指数(H)、Pielou 均匀度指数(J)和Margalef 丰富度指数(D),对生态修复区的生物群落结构特征进行分析评价。计算公式如下:
式中:ni为群落中第i个物种的个体数;N为群落中总个体数,二者之比为物种i 的丰富度;fi为该种在各位点中出现的频率。本研究中因每次调查均统计各样点每个物种的总数,或者采用样线法,因此fi均为1。Y>0.02时定义为优势种[16],鱼类、鸟类丰富度柱状图中将非优势种合并统称为其他。
2 结果与讨论
2.1 水质指标的变化
本研究表明,水生态系统逐步建立后,对水质具有很好的改善作用,尤其是对总磷、氨氮、总氮有明显的降低作用。在为期1年的水质监测期内,虽然试验区与上游来水的总磷、氨氮、总氮基本保持同样的变化趋势,但是在2020年10 月生态系统基本建立后试验区内水质总体明显优于上游来水水质,其中总磷浓度平均低35%,氨氮浓度平均低31%,总氮浓度平均低30%(图2)。本研究中重建的水生态系统,不仅发挥了沉水植物通过生物吸附、富集作用去除氮、磷污染物并为底栖动物、小型鱼类提供避难所和栖息地的作用[3,4,17-22],而且通过水生植物—浮游动物—底栖动物—鱼类的序批式投放,能在2 个月内快速建立稳定的生态系统,从而发挥整个生态系统更持久、有效的水质净化作用。

图2 试验区内外水体总磷、氨氮、总氮浓度变化Fig.2 Changes of total phosphorus, ammonia nitrogen and total nitrogen in and out of the experimental area
也有指标表现出相反趋势。试验区内的化学需氧量在2020年10 月—2021年2 月均高于上游来水,平均高15%,2021年3—8 月均低于上游来水,平均低30%(图3),这可能是因为冬季鱼类生命活动不活跃而浮游动物相对较多,水体中出现大量浮游动物尸体而导致有机质含量升高。试验区内水体溶解氧浓度在2020年12 月—2021年5 月均低于上游来水,平均低23%(图3),可能是由于试验区水体水生动物丰富而消耗氧气导致[23]。因此,水生生物群落结构与水环境之间存在双向互馈作用。

图3 试验区内外水体化学需氧量、溶解氧浓度变化Fig.3 Changes of chemical oxygen demand and dissolved oxygen in and out of the experimental area
2.2 物种丰度的变化
水生生物的变化是水环境质量变化的直观体现,具有高敏感性、长期性、富集性、综合性等优势[24]。各生物类群在2020年7 月水生态修复工程开始后物种丰度均有明显上升。生态修复实施后浮游动物、底栖动物、鱼类物种丰度分别为之前的6、2、3 倍。除鸟类外其他营养级物种丰度均在越冬后的春夏季节出现明显的跃升并高于前一个夏季。鸟类物种丰度在工程实施后的秋季迅速增长为实施前的2 倍,虽然冬、春季由于水位下降严重导致鸟类数量减少,但仍在2021年5 月监测到试验开始时的近2 倍丰度(图4)。
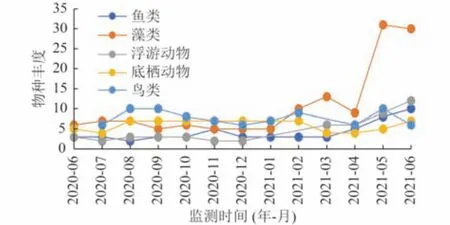
图4 监测期间各类群物种丰度的变化Fig.4 Change of the species richness during the monitoring
浮游动物前期以试验投放的枝角类为主,后期除枝角类外还出现多种轮虫类并成为优势种。冬季浮游动物物种稀少,其中2021年1 月、2 月未监测到浮游动物,3 月后物种丰度迅速上升。总体而言试验后浮游动物物种丰度和多样性均有明显的提升(表3)。
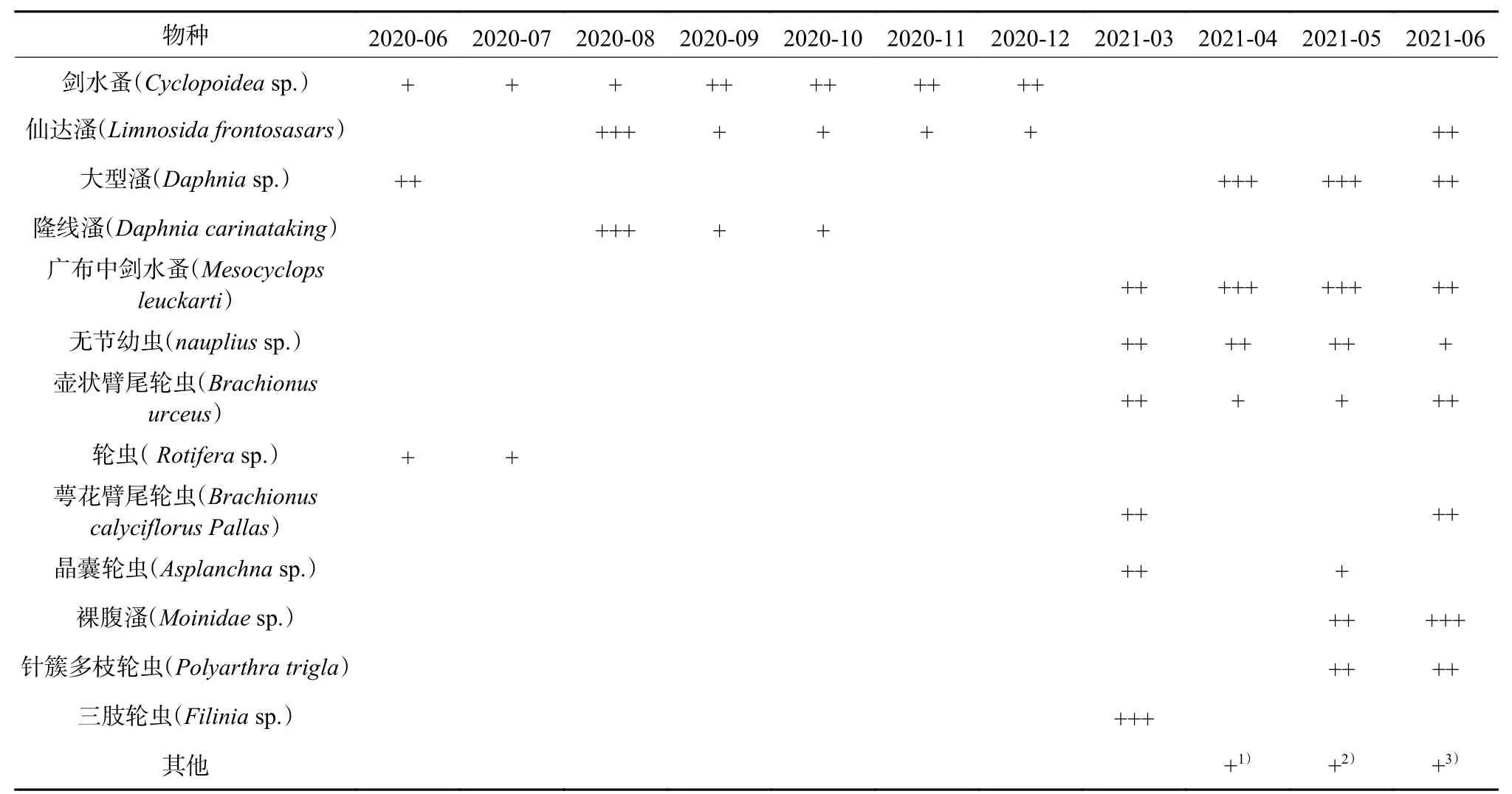
表3 历次监测中的浮游动物物种及相对丰度Table 3 Species and relative abundance of zooplankton during the monitoring
本研究中浮游动物物种丰度和群落结构的变化体现了季节规律性。多项研究表明水温通过影响浮游动物的体温、新陈代谢、生长发育及繁殖周期,进而影响浮游动物物种丰度及群落结构[15,25-27]。有研究表明,温暖水体适宜轮虫生活[28],而高温不利于枝角类部分物种的生长发育[29]。但涡河中的研究[30]得出了相反的结论。本研究中在第二年夏季,轮虫丰度和多样性明显高于枝角类,且在寒冷季节仅监测到枝角类,这与前者结论相符合。浮游动物的群落结构还受到营养盐、有机质、溶解氧等多重因素的影响。本研究中水体总磷、氨氮、总氮浓度及化学需氧量自2021年3 月起均表现出下降趋势,而同期浮游动物丰度和多样性增加。在温瑞塘河的研究中浮游动物密度与COD 呈负相关[31],在妫水河的研究中浮游动物多样性指数随水质变差而降低[27],这与本研究得出结论一致。因此,浮游动物的多样性指数可以在一定程度上反映出水环境的状况,也可作为环境质量评价的依据之一[27,32]。
2.3 生物多样性指数的变化
底栖动物的Shannon-Wiener 多样性指数、Pielou均匀度指数、Margalef 丰富度指数分别最高提升108%、72%、87%,至1年监测期满时分别提升60%、32%、33%(图5)。多样性指数和丰富度指数在试验开始后均明显上升,但是在2021年3—4 月均出现低谷值,这体现了底栖动物群落对2021年2—3 月水体污染物浓度达到峰值的响应,证明底栖动物的多样性指数可随水质和栖息地条件恶化而下降[33]。群落结构也发生了明显变化,优势种明显地由耐污型物种向高水质要求、净化能力强的物种变化。前期以摇蚊幼虫(Chironomidaesp.)、水丝蚓(Limnodrilussp.)为绝对优势种,之后随着生态系统逐渐健全二者优势度呈明显下降趋势。2020年 8 月以后投放的螺类和黑壳虾逐渐成为底栖动物优势种,体现出二者对水质波动的耐受性和净化作用。但螺类优势度在2020年12 月开始出现下降,主要是因为2020年12 月—2021年5 月河道水位严重下降甚至出现部分河底裸露的现象。有研究表明,水生植物可为大型底栖动物群落提供适宜生境,从而改善城市河流中底栖动物的营养层级关系[17]。本研究中底栖动物群落结构的变化也说明了这一点。

图5 底栖动物多样性指数及丰富度变化Fig.5 Changes of diversity indices and abundance of zoobenthos
鱼类的Shannon-Wiener 多样性指数、Margalef丰富度指数均呈上升趋势,尤其是2021年3 月以后更为明显,分别最高提升138%、113% 且均出现在1年监测期满时。两指数的季节差异较小,而且即使在水质较差时期也未表现出明显下降,这说明鱼类的短时耐受性更强一些。Pielou 均匀度指数最高提升43%,至1年监测期满时提升19%(图6)。鱼类群落结构的变化体现了水生态修复的效果,包括优势种向水质健康型转变和优势物种数量增加。原生鱼类主要为水质亚健康指示物种鲫、麦穗鱼。自试验投放后除2021年3 月外每次监测都能采集到草金鱼,说明草金鱼已稳定繁殖。2021年起逐渐出现鳑鲏、鰕虎鱼、青鳉、棒花鱼等并且成为优势种(图6),其中鳑鲏、青鳉为水质健康指示物种。虽然鱼类能有效反映河流的综合生态条件,但已有研究中较少采用鱼类作为生态系统状态单一评价指标,而是常用作综合指标系统的一项[34-36]。有研究表明,采用单一类群的生物多样性指数进行生态系统健康评价可能存在偏差,采用多类群的综合指数则能够更好地反映客观情况[37]。

图6 鱼类多样性指数及丰富度变化Fig.6 Changes of diversity indices and abundance of fish
鸟类作为水域生态系统食物链的顶端生物,其多样性受到食物、季节和栖息地环境等多种因素的综合影响。本研究中鸟类的Shannon-Wiener 多样性指数、Pielou 均匀度指数、Margalef 丰富度指数均呈先下降后上升趋势,分别最高提升26%、73%,至1年监测期满时分别降低7%、2%(图7)。2020年8 月、9 月投放底栖动物和部分鱼类后鸟类多样性、丰富度有所上升,但随即进入下降趋势直至2021年5 月重新回升,这可能与家燕等候鸟迁徙有关。从优势种来看,除麻雀外其他优势种呈现出比较明显的季节变化。2020年秋、冬以黑水鸡、喜鹊、珠颈斑鸠、小䴙䴘为优势种,2021年春以珠颈斑鸠、喜鹊、家燕、白鹡鸰等为优势种,优势种数量基本持平。白腰草鹬、红隼、短耳鸮、绿翅鸭仅出现在了冬季。

图7 鸟类多样性指数及丰富度变化Fig.7 Changes of diversity indices and abundance of birds
由分析可知,鸟类的丰富度、多样性指数与鱼类的丰富度和多样性指数并未表现出相同趋势,但与水环境质量表现出明显的相同趋势,即与水体污染程度呈负相关,这与罗莎等[38]的研究成果一致。此外,项目区自2020年10 月起受到跨河桥梁施工的影响,河道水位下降、岸坡多处被破坏,导致了鸟类的食物及生境的变化。喜鹊、珠颈斑鸠逐渐成为优势种可能与滨水地带大量土地裸露有关。综合来看,鸟类对生态修复效果具有一定的综合指示性作用[39]。本研究中还在项目水域观测到猛禽短耳鸮、大鵟、红隼,说明区域生物多样性较高,生物链已接近顶端。
2.4 水生态系统的抗冲击能力及效果持续性
水生态系统的健康持续性是水生态系统健康水平评判的最高层级,一个健康的水生态系统应该具有抵抗干扰、恢复自身结构和功能持续性的能力[40]。
试验期间虽然经历了长达7 个月的上游来水水质水量的波动,甚至出现几近干涸的情况,但是从监测结果来看,所构建的生态系统经受住了水质水量波动的冲击,表现为物种丰度未出现明显下降、生物多样性指数在投放当年的冬季仍维持较高水平,且春季水质水量冲击缓解后均有明显回升,并且持续发挥水质净化效果。这证明了本试验构建的水生态系统具有较高的健康水平,可以抵抗一定程度的持续污染输入或水量波动[41]。从各类群的抗冲击能力差别来看,浮游植物、浮游动物、鱼类的抗冲击性比底栖动物、鸟类更强,表现为波动更小甚至未出现降低趋势,鱼类生物多样性指数还出现了较大幅度的提升。底栖动物的波动主要是因为水量水位的急剧下降,尤其是2021年4—5 月出现部分河床裸露导致大量螺类死亡,但是从优势种的丰富度对比来看,黑壳虾、螺类仍然具有较强的抗冲击性,丰富度占比呈波动上升趋势,也证明了本项目投放物种的适宜性。因具有较强的迁徙能力,鸟类的波动则是食物、栖息地的综合作用,对系统抗冲击能力的体现作用较小。
根据项目区生物多样性指数分析可知,本研究中构建的水生态系统不仅在初期发挥了快速恢复水体生物、提高生物多样性的作用,而且效果可持续1年甚至会继续上升。根据项目区水质变化分析可知,本研究中构建的生态系统不仅在1年内发挥了较强的水质净化作用而且在1年后仍保持较高的污染物削减率。
以上综合说明本试验构建的生态系统可耐北方严寒季节考验、可在一定时期内持续发挥提升水生态系统健康的作用。根据已有研究,河流生态修复工程对施工河段生物多样性提升、水质改善的有效作用期为5年左右[5,11],因此本项目中生态修复措施的效果持续性还有待持续监测。
3 结论
(1)在再生水补给为主要水源的河道中,以生物多样性提升为导向的水生态修复具有较好的水质改善效果,尤其对氮磷削减效果明显,总磷削减率可达35%,氨氮削减率可达31%,总氮削减率可达30%。也有一些指标,如化学需氧量、溶解氧可能因水生生物的生长出现阶段性降低现象。总体来看生态修复对再生水水质的改善效果在1年内保持较高且稳定的水平。
(2)通过构建完整生态系统可快速提升生物多样性并建立稳定、可持续系统。物种丰度提升2~6 倍,底栖动物、鱼类、鸟类的Shannon-Wiener 多样性指数分别最高提升108%、138%、26%,Margalef丰富度指数分别最高提升87%、112%、73%;底栖动物、鱼类的Pielou 均匀度指数分别最高提升72%、43%,鸟类的Pielou 均匀度指数则下降7%。鸟类生物多样性指数的变化体现出季节、食物、生境等因素的综合指示效果。底栖动物、鱼类群落的优势种呈现数量增多且由耐污型向健康指示型的转变。生物多样性指数结合优势种和群落结构可以更好地判断生态系统结构的变化。多个生物多样性指数的综合表征效果更全面、更客观。
(3)完整的生态系统具有一定时期内抵抗水质、水量波动的韧性,有助于系统的水质恢复和生物系统重建。浮游植物、浮游动物、鱼类具有更强的抗水质、水量波动的能力,底栖动物受水位影响更明显。
猜你喜欢
学与玩(2022年9期)2022-10-31
海洋信息技术与应用(2022年1期)2022-06-05
儿童时代·幸福宝宝(2020年9期)2020-09-08
文苑(2020年12期)2020-04-13
小太阳画报(2019年1期)2019-06-11
作文周刊·小学二年级版(2018年21期)2018-09-06
现代园艺(2018年1期)2018-03-15
中国资源综合利用(2017年4期)2018-01-22
小学生必读(低年级版)(2017年5期)2017-08-12
金色少年(奇趣科普)(2016年8期)2016-09-21
