
miR-146b-3p 通过靶向结合LCN1 抑制肺鳞癌生物学行为
2023-07-20 02:12丁晓霞刘亚群于小纬张悦琪河北北方学院附属第一医院呼吸与危重症监护室河北张家口075000
局解手术学杂志 2023年7期
丁晓霞,刘亚群,于小纬,杨 楠,张悦琪(河北北方学院附属第一医院呼吸与危重症监护室,河北 张家口 075000)
肺癌是导致癌症患者死亡的重要原因,肺鳞癌是常见的肺癌亚型之一,其化疗和放疗敏感性较低[1-2]。由于肺鳞癌容易发生转移,并且目前缺乏有效的诊断性生物标志物,确诊时患者往往已处于中晚期,导致患者预后较差[3-4]。因此,积极寻找用于肺鳞癌诊断和预后预测的生物标志物至关重要。微小RNA(microRNA,miRNA)是一类不具有编码能力的小分子RNA,在基因转录中发挥重要作用,参与细胞增殖、迁移、侵袭和凋亡等生物行为学过程[5-6]。脂质运载蛋白1(lipocalin-1,LCN1) 能够间接导致肺组织基质降解增加,造成肺组织结构重塑,使病情加重。miRNA 可长期稳定存在,且其异常表达会影响肿瘤的发生发展[7-8]。miRNA 既可以发挥抑癌作用,也可以发挥促癌作用,并且在多数人类恶性肿瘤中异常表达[9-10]。有研究显示,miR-146b-3p 可作为诊断非小细胞肺癌的重要标志物[11]。还有研究显示,miR-146a 在肺鳞癌组织中表达下调,其异常表达能抑制肺鳞癌细胞的迁移和侵袭[12]。基于这些研究结果,我们推测miR-146b-3p可能在肺癌中发挥重要作用。本研究探讨miR-146b-3p差异表达对肺鳞癌细胞生物学行为的影响,并初步分析其可能的作用机制,以期为肺鳞癌的治疗提供理论依据。
1 材料与方法
1.1 细胞、试剂与仪器
NCI-H226、NCI-H2170 细胞株购于中国科学院上海生命科学研究院细胞资源中心;RPMI 1640培养基、AnnexinV-FITC 凋亡检测试剂盒购于上海经科化学科技有限公司;胎牛血清购于爱必信(上海)生物科技有限公司;细胞计数仪购于Invitrogen 有限公司;RNA 提取试剂TRIzol 及转染试剂Lipofectamine 2000 均购自南京森贝伽生物科技有限公司;逆转录试剂盒Prime-ScriptTMRT Master Mix 购自上海妍琦生物科技有限公司,PCR 试剂盒Quanti Fast SYBR Green PCR Kit 购自武汉极捷生物科技有限公司;BCA 蛋白浓度测定试剂盒、SDS-PAGE 凝胶配制试剂盒购自北京伊塔生物科技有限公司;CCK-8 试剂盒购自上海吉至生化科技有限公司;BCL2、BAX、CCNB1、CDK2 多克隆抗体、LCN1多克隆抗体以及HRP 标记的羊抗兔免疫球蛋白G(H+L)二抗购自上海科敏生物科技有限公司;双荧光素酶报告基因系统试剂盒购自成都安森盛源科技有限公司;ST-360 型酶标仪购自上海晚成医疗器械有限公司;细胞周期试剂盒购于杭州联科生物技术股份有限公司;ELx808 酶标仪购自北京泽平科技有限责任公司;FACS VantageTM流式细胞仪购自南京华奥生物医药技术有限公司。
1.2 生物信息学分析
通过TCGA 在线数据库下载肺鳞癌临床样本数据,比较miR-146b-3p、LCN1在癌旁组织与肺鳞癌组织中的表达情况,并分析LCN1 与肺鳞癌患者临床表型的关系。使用Kaplan-Meier Plotter 数据库评估LCN1 mRNA水平对肺鳞癌患者总生存期的影响。利用Star-Base 数据库在线预测靶基因数据库预测miR-146b-3p的靶基因。
1.3 细胞培养
将NCI-H226、NCI-H2170 细胞接种至含10%胎牛血清的RPMI 1640 培养基,在37 ℃、5%CO2饱和湿度条件下培养,待细胞贴壁生长,胰酶消化液消化后传代培养,实验取对数生长期细胞。
1.4 RT-PCR检测miR-146b-3p表达
采用TRIzol 法提取细胞或组织中的总RNA,采用紫外分光光度计法测定RNA 的浓度。按TaqMan©MicroRNA Reverse Transcription kit 逆转录试剂盒说明书以RNA 为模板进行逆转录获得cDNA 并置于4 ℃保存,进行荧光定量PCR 扩增,反应条件:95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火45 s,65 ℃延伸1 min,4 ℃ 10 min,扩增40个循环。以β-actin 为内参,miR-146b-3p 上 游 引 物 序 列 :5′-CCTGGCA CTGAGAACTGAAT-3′,下游序列:5′-GCACCAGAA CTGAGTCCACA-3′;β-actin 上游引物序列:5′-CATC CCTTCTCCCCACACAC-3′,下游序列:5′-AGTCCCAG GGCTTTGATTTG-3′。以2-ΔΔCt法计算miR-146b-3p 的相对表达水平。
1.5 构建miR-146b-3p过表达细胞模型
将对数生长期的NCI-H2170 和NCI-H226 细胞分为miR-NC 组和miR-146b-3p 组,根据Lipofectamine 2000 说明书分别转染miR-146b-3p NC 和miR-146b-3p质粒,24 h后更换完全培养基。采用G418筛选出能够稳定过表达miR-146b-3p的NCI-H2170和NCI-H226细胞系。采用RT-PCR 检测miR-146b-3p 水平以评估转染效率。
1.6 构建LCN1敲低细胞模型
将对数生长期的NCI-H2170 和NCI-H226 细胞分为shNC 组、shLCN1 1#组、shLCN1 2#组和miR-146b-3p+LCN1 组,并根据Lipofectamine 2000 说明书分别转染shNC、shLCN1 1#、shLCN1 2#质粒,24 h 后更换完全培养基。用嘌呤霉素筛选出稳定敲低LCN1 的NCI-H2170 和NCI-H226 细胞系。采用Western blot 检测LCN1 蛋白水平以评估转染效率,筛选稳定质粒构建miR-146b-3p+LCN1 组(共转染miR-146b-3p+LCN1 2#质粒)。
1.7 细胞增殖实验
将miR-NC 组、miR-146b-3p 组、shNC 组、shLCN1 1#组、shLCN1 2#组和miR-146b-3p+LCN1 组细胞置于CO2培养箱中培养24 h,分别于0、1、2、3、4、5 d 时对细胞增殖活性进行检测。每孔加入CKK-8 10 μL,在37 ℃、5%CO2的培养箱中继续培养1 h,随后用酶标仪测定450 nm波长处的光密度(optical density,OD)值。
1.8 细胞克隆实验
将miR-NC 组、miR-146b-3p 组、shNC 组、shLCN1 1#组、shLCN1 2#组和miR-146b-3p+LCN1 组细胞分别接种到6 孔板中,并轻轻转动,使细胞分散均匀。用PBS 清洗3 次,再用40 g/L 多聚甲醛固定20 min。结晶紫染色液染色,光学显微镜下观察细胞集落。
1.9 细胞周期实验
将miR-NC 组、miR-146b-3p 组、shNC 组、shLCN1 1#组、shLCN1 2#组和miR-146b-3p+LCN1 组细胞以1×105个/孔的密度接种于6 孔板,消化离心,弃去上清液,收集细胞团块,PBS 重悬细胞,离心,清洗1 次,振荡,使细胞沉淀充分分散,先加入300 μL、4 ℃预冷PBS,再将上样管置于涡旋振荡器上振荡,加700 μL、4 ℃预冷的纯乙醇沉悬,4 ℃固定过夜。第2天,离心弃去上清液,PBS洗1次,加500 μL PI染液,室温避光孵育15 min,上流式细胞仪检测,用Flowjo 软件分析细胞周期时相分布。
1.10 细胞凋亡实验
取miR-NC 组、miR-146b-3p 组、shNC 组、shLCN1 1#组、shLCN1 2#组和miR-146b-3p+LCN1 组细胞进行消化、离心、重悬。将细胞接种于6 孔板内并置于37 ℃、5%CO2的培养箱中培养24 h。以1 500 r/min 离心10 min 去除上清液。加入5 μL AnnexinV-FITC 混匀,再加入5 μL PI混匀,随后室温下避光孵育15 min。采用流式细胞仪检测细胞凋亡情况。
1.11 双荧光素酶报告实验
构建野生型报告基因载体LCN1-WT 与突变型报告基因载体LCN1-MT,将载体分别与miR-NC 或miR-146b-3p 共转染到肺鳞癌细胞中。置于37 ℃、5%CO2培养箱继续培养48 h,用双荧光素酶检测试剂盒测量各组细胞中的荧光素酶活性。
1.12 裸鼠移植瘤实验
16 只裸鼠购自成都达硕实验动物有限公司,动物生产许可证号:SCXK(川)2020-030,在标准环境中饲养,自由采食饮水。将小鼠分为对照组和观察组,每组8 只,对照组裸鼠于右侧腹股沟皮下注射0.2 m L(1×107/mL)转染miR-NC 的NCI-H2170 细胞悬液,观察组裸鼠于右侧腹股沟皮下注射0.2 mL(1×107/mL)已转染miR-146b-3 的NCI-H2170 细胞悬液。待成瘤后,每2 d 用微米卡尺测量肿瘤体积,于第2 周末处死小鼠,剥离移植瘤并称重,RT-PCP 检测移植瘤组织中miR-146b-3p的表达。
1.13 Western blot检测
收集miR-NC组、miR-146b-3p组、shNC组、shLCN1 1#组、shLCN1 2#组、miR-146b-3p+LCN1组对数生长期的NCI-H2170 和NCI-H226 细胞,加入SDS Loading buffer,沸水浴中煮沸10 min,电泳后转至PVDF 膜,用5%脱脂牛奶室温封闭1 h。加入用封闭液稀释的抗体(1∶5 000),4 ℃孵育过夜。次日PBS 洗涤6 min,重复3 次,然后加入封闭液稀释的HRP 标记的羊抗兔二抗(1∶1 000),室温孵育60 min,PBS 洗涤6 min,重复3 次,ECL 化学发光液进行显影,以β-actin 作为内参,利用BCA 蛋白检测试剂盒检测蛋白浓度,具体操作遵照试剂盒说明书进行。
1.14 统计学处理
数据采用SPSS 17.0 软件进行统计学处理,计量资料以均数±标准差(±s)表示,2 组间的表达差异采用t检验分析,多组间比较采用单因素方差分析。Kaplan-Meier生存曲线分析患者总生存期。P<0.05 表示差异具有统计学意义。
2 结果
2.1 miR-146b-3p 过表达抑制肺鳞癌细胞增殖、促进凋亡
GEO数据库分析显示,肺鳞癌组织中miR-146b-3p表达水平明显低于癌旁组织(P<0.05)。RT-PCR 实验结果显示,miR-146b-3p 组中miR-146b-3p 表达明显高于miR-NC 组(P<0.05)。细胞增殖实验结果显示,与miR-NC 组相比,miR-146b-3p 组增殖能力明显降低(P<0.05)。细胞克隆实验结果显示,与miR-NC 组相比,miR-146b-3p 组克隆细胞数明显减少(P<0.05)。细胞周期实验结果显示,与miR-NC组相比,miR-146b-3p组G0/G1 期细胞数明显增加(P<0.05),S 期细胞数明显减少(P<0.05)。细胞凋亡实验结果显示,与miRNC 组相比,miR-146b-3p 组细胞凋亡率明显增高(P<0.05)。Western blot结果显示,与miR-NC组相比,miR-146b-3p 组细胞BCL2、CCNB1 和CDK2 蛋白减少(P<0.05),BAX蛋白增加(P<0.05),见图1。
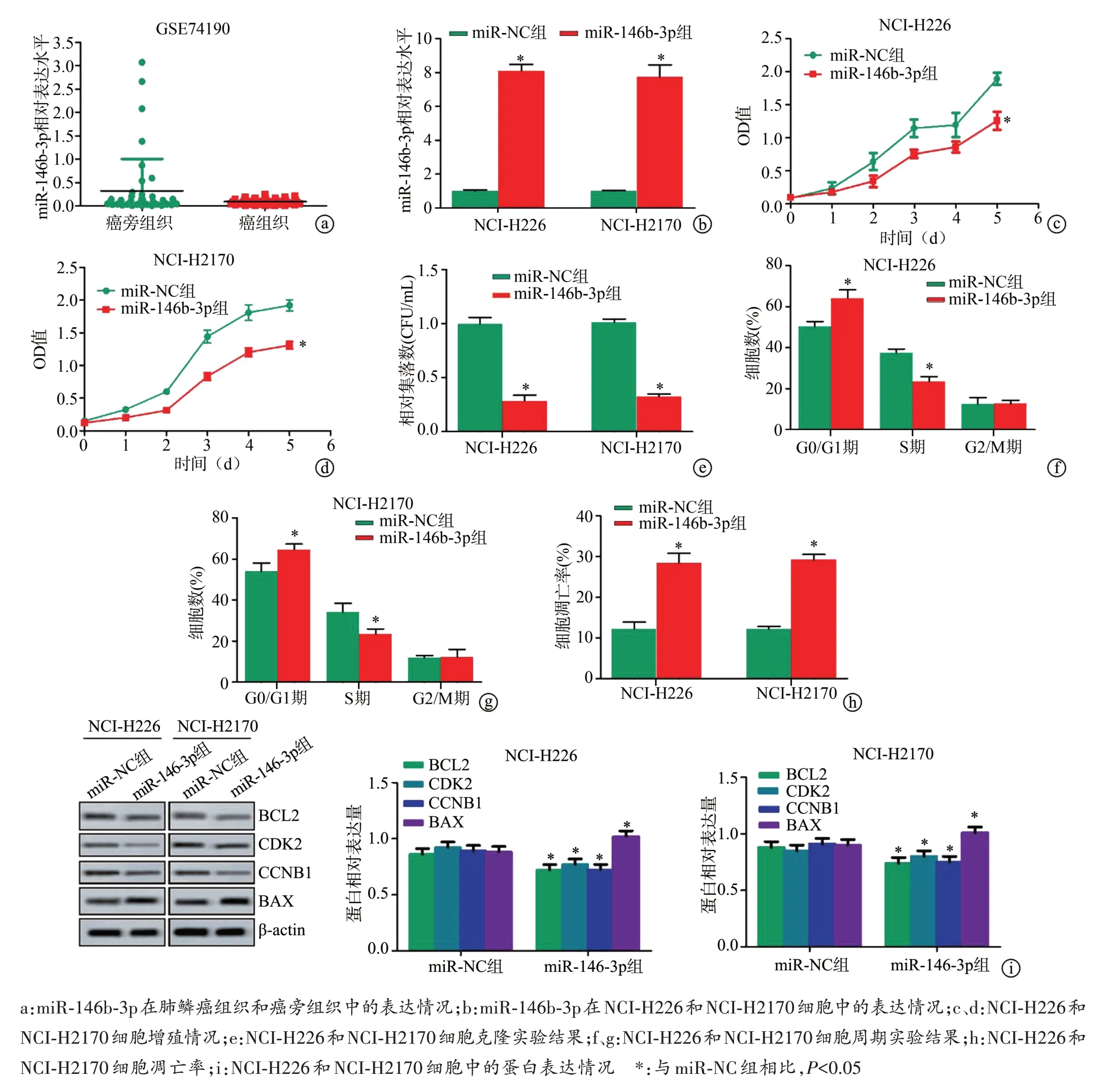
图1 miR-146b-3p抑制肺鳞癌细胞增殖并促进其凋亡
2.2 miR-146b-3p过表达抑制移植瘤生长
裸鼠移植瘤实验结果显示,观察组移植瘤体积(468.57±50.63)mm3和体质量(0.38±0.04)g明显小于对照组的体积(287.91±50.63)mm3和体质量(0.19±0.02)g(P<0.05)。随着时间的推移,移植瘤体积不断增大,但是观察组的增长速度明显慢于对照组(P<0.05)。RT-PCR 结果显示,在裸鼠移植瘤组织中,观察组中miR-146b-3p mRNA 相对表达量明显高于对照组(P<0.05),见图2。
2.3 miR-146b-3p与LCN1的靶向关系
生物信息学分析结果表明,miR-146b-3p 序列3′-UTR 区与LCN1 存在结合位点。双荧光素酶报告实验显示,miR-146b-3p mimic 与LCN1-WT 共转染致荧光素酶活性降低(P<0.05)。Western blot 和RTPCR实验结果显示,与miR-NC组相比,miR-146b-3p组中LCN1 mRNA 和蛋白表达水平明显降低(P<0.05),见图3。

图3 miR-146b-3p与LCN1的靶向关系
2.4 LCN1 在肺鳞癌不同临床特征患者癌组织中的表达水平
TCGA 数据库分析显示,肺鳞癌组织中LCN1 表达高于癌旁组织(P<0.05),M1 期患者LCN1 表达高于M0 期患者(P<0.05),预后不良患者LCN1 表达高于预后良好患者(P<0.05),LCN1 高表达患者总生存率明显低于LCN1低表达患者(P<0.05),见图4。
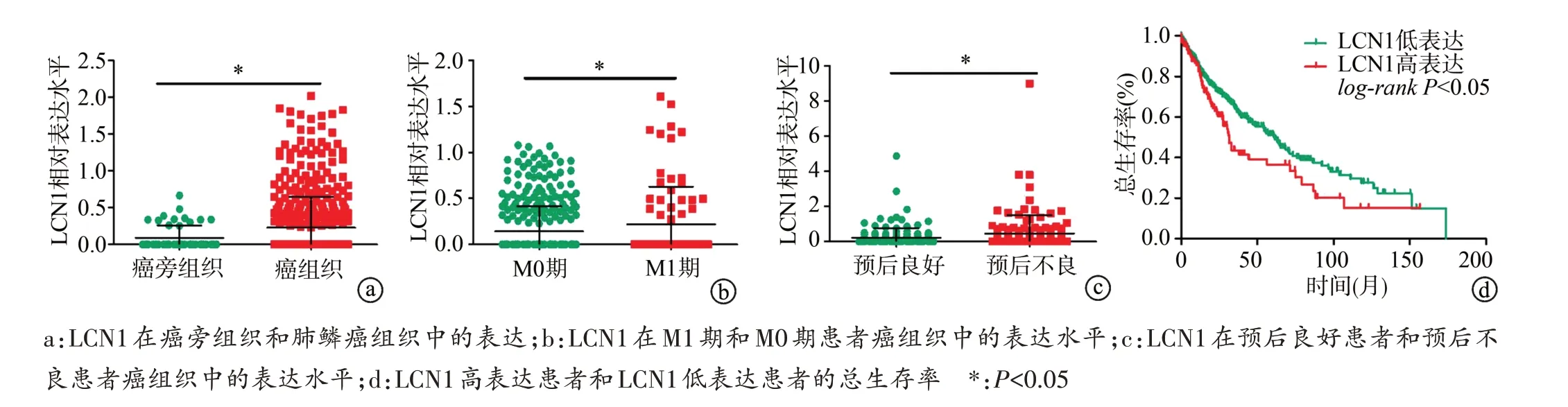
图4 LCN1在肺鳞癌不同临床特征患者癌组织中的表达水平
2.5 敲低LCN1表达抑制肺鳞癌细胞增殖并促进其凋亡
Western blot 实验结果显示,与shNC 组相比,shLCN1 1#组和shLCN1 2#组中LCN1 蛋白表达明显降低(P<0.05)。细胞增殖实验结果显示,与shNC组相比,shLCN1 1#组和shLCN1 2#组中细胞增殖能力降低(P<0.05)。细胞克隆实验结果显示,与shNC 组相比,shLCN1 1#组和shLCN1 2#组中克隆细胞数明显减少(P<0.05)。细胞周期实验结果显示,与shNC组相比,shLCN1 1#组和shLCN1 2#组G0/G1 期细胞数明显增加,S 期细胞数明显减少(P<0.05)。细胞凋亡实验结果显示,与shNC 组相比,shLCN1 1#组和shLCN1 2#组中凋亡率明显增高(P<0.05),见图5。

图5 敲低LCN1表达抑制肺鳞癌细胞增殖并促进其凋亡
2.6 miR-146b-3p 靶向LCN1 抑制肺鳞癌增殖并促进其凋亡
Western blot 实验结果显示,与miR-NC 组及miR-146b-3p+LCN1组相比,miR-146b-3p组中LCN1蛋白相对表达水平下降(P<0.05)。细胞增殖实验结果表明,与miR-NC 组及miR-146b-3p+LCN1 组相比,miR-146b-3p 组中增殖能力降低(P<0.05)。细胞克隆实验结果显示,与miR-NC 组及miR-146b-3p+LCN1 组相比,miR-146b-3p组中克隆细胞数明显减少(P<0.05)。细胞周期实验结果显示,与miR-NC 组及miR-146b-3p+LCN1 组相比,miR-146-3p 组中G0/G1 期细胞数量明显增加,S 期细胞数量明显减少(P<0.05)。细胞凋亡实验结果显示,与miR-NC 组及miR-146b-3p+LCN1组相比,miR-146b-3p组中凋亡细胞数明显增加(P<0.05),见图6。
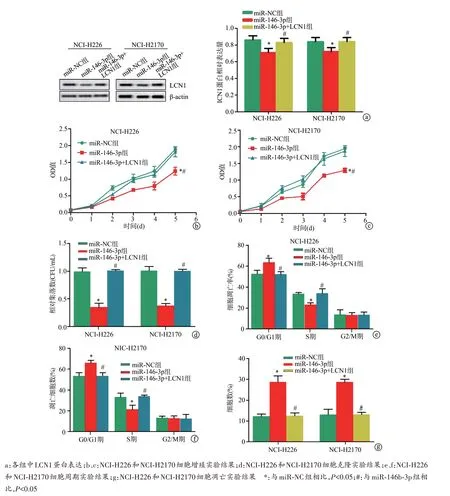
图6 miR-146b-3p靶向LCN1抑制肺鳞癌细胞增殖并促进其凋亡
3 讨论
肺鳞癌是非小细胞肺癌的常见类型,早期并无明显特异性,确诊时大多已处于中晚期,错过了最佳手术时机,因而通常采取化疗等保守治疗,但长期使用各种类型的抗癌药物会导致机体出现耐药性,从而影响治疗效果[13-17]。随着肿瘤精准治疗的快速发展,对肿瘤生物学行为和基因的调控成为了研究重点。其中miRNA 在肿瘤细胞增殖、迁移、侵袭、分化、凋亡、化疗耐药等多种生理过程中起到关键作用[18-19]。本研究旨在寻找影响肺鳞癌生物学行为的潜在标志物,为临床治疗提供理论依据。
本研究通过生物信息学分析发现,miR-146b-3p在肺鳞癌组织中低表达;LCN1 在肺鳞癌组织中高表达,且LCN1 差异表达与肺鳞癌临床预后不良及M 分期有关。构建miR-146b-3p 过表达细胞模型进行一系列体外功能试验,结果显示,肺鳞癌细胞过表达miR-146b-3p后,miR-146b-3p表达水平增加,细胞增殖和克隆能力降低,细胞周期受到阻滞,细胞凋亡明显增加,提示miR-146b-3p 过表达可抑制肺鳞癌细胞增殖,阻滞细胞周期,并诱导肿瘤细胞凋亡。进一步行Western blot分析显示,肺鳞癌细胞过表达miR-146b-3p后,细胞BCL2、CCNB1和CDK2蛋白减少,BAX 蛋白增加,表明过表达miR-146b-3p 可调节肺鳞癌细胞凋亡相关蛋白表达。促凋亡蛋白BAX 和抗凋亡蛋白BCL2在凋亡信号介导的凋亡过程中起关键作用,BAX 过表达可拮抗BCL2 表达,促进细胞凋亡。因此,上调BCL2 表达、下调BAX 表达可抑制细胞凋亡。众所周知,miRNA 通过抑制或降解靶基因mRNA 发挥生物学功能[14]。既往研究显示,miR-146b-5p 可通过靶向调节ZNRF2 表达诱导乳腺癌细胞增殖、迁移和侵袭[20]。因此,结合本研究结果推测miR-146b-3p 可抑制肺鳞癌细胞增殖,调节细胞周期相关蛋白,进而阻滞细胞周期,并通过上调BAX 和下调BCL2蛋白表达诱导细胞凋亡。本研究通过裸鼠移植瘤实验进一步发现,miR-146b-3p 过表达可抑制裸鼠移植瘤的生长,证实了miR-146b-3p 过表达在肺鳞癌中发挥抑制作用。
LCN1 是脂质运载蛋白家族的成员,是泪液中重要的脂质结合蛋白,分布在多种组织和腺体中,并与多种疾病的发生有关[21-22]。既往研究显示,LCN1 在乳腺癌中高表达,且其异常表达与临床病理特征和生存率较低有关[23]。本研究通过miRNA 靶基因数据库预测miR-146b-3p 调控肺鳞癌细胞的靶基因,结果显示,miR-146b-3p 存在与LCN1 靶向结合的位点,LCN1 是miR-146b-3p 的靶基因。Western blot 结果显示,miR-146b-3p 组中LCN1 蛋白表达水平明显低于miR-NC组,推测miR146b-3p 可以调控LCN1 蛋白表达。本研究发现,肺鳞癌组织中LCN1 表达高于癌旁组织,且与肺鳞癌患者M 分期存在一定关联。为进一步分析LCN1 在肺鳞癌中发挥的生物学功能,本研究通过构建LCN1 敲低细胞模型进行体外功能实验,结果显示,LCN1 敲低可以抑制肺鳞癌细胞生长、增殖,阻滞细胞周期,并诱导细胞凋亡。提示LCN1 在肺鳞癌中高表达,敲低LCN1表达可影响肺鳞癌生物学行为。
本研究结果显示,miR-146b-3p 过表达降低了肺鳞癌细胞增殖能力,减少了克隆细胞数,阻滞了细胞生长周期,增加了凋亡细胞数量;而miR-146b-3p 和LCN1 共转染后,逆转了miR-146b-3p 对肺鳞癌的抑制作用。故我们推测,miR-146b-3p 在肺鳞癌中发挥促癌作用,其可能通过靶向抑制LCN1 表达调控肺鳞癌生物学行为。
综上,本研究发现miR-146b-3p 在肺鳞癌细胞中低表达,LCN1在肺鳞癌细胞中高表达,且miR-146b-3p与LCN1 存在靶向结合位点。上调miR-146b-3p 可以抑制肺鳞癌细胞增殖,诱导细胞凋亡,并影响细胞生长周期。而miR-146b-3p 过表达可以抑制移植瘤生长,其作用机制可能是通过靶向LCN1实现的。
猜你喜欢
云南医药(2021年3期)2021-07-21
四川文理学院学报(2020年5期)2020-02-12
国际呼吸杂志(2019年4期)2019-03-12
中国报道(2018年2期)2018-04-20
绿色中国(2016年1期)2016-06-05
中华老年多器官疾病杂志(2016年7期)2016-04-28
癌症进展(2016年10期)2016-03-20
医学研究杂志(2015年5期)2015-06-10
癌变·畸变·突变(2015年3期)2015-02-27
中国中西医结合外科杂志(2013年3期)2013-03-11
