
水稻亮绿叶突变体bgl-2的表型特征与遗传分析
2023-07-20 05:11:38林秋云谢振宇胡伟周玉杰龙开意贺治洲
热带作物学报 2023年6期
林秋云 谢振宇 胡伟 周玉杰 龙开意 贺治洲


关键词:水稻;叶片亮绿;突变体;bgl-2;表型;遗传
中图分类号:S31 文献标识码:A
粮食安全是“国之大者”,悠悠万事,吃饭为大。我国是水稻生产大国,2011—2021 年种植面积稳定在3000 万hm2 左右,以占世界9%的耕地、6%的淡水资源,养育了世界近1/5 的人口[1]。优良高产品种的推广应用发挥了极其重要的作用。光合效率是决定作物产量的重要前提,也是育种家们的改良目标,而叶片是水稻进行光能转化的主要组织[2]。水稻叶色突变是一种发生频率高、易于被发现的突变现象,但多数叶色突变体对光合作用有影响,在一定程度上间接影响水稻产量,因此在早期常被认为是无意义突变体。随着生物技术的发展,大量研究表明叶色突变体不仅是开展光合作用、光形态建成、质-核基因互作、叶绿素合成和叶绿体发育等基础研究的理想材料,同时可作为一种明显且易于识别的标记性状,简化良种繁育和杂交种生产[3-5]。此外,叶色突变体还可以作为观赏稻,致力于打造乡村休闲农业观光旅游点,带动农民增收,促进乡村振兴[6]。
水稻资源辐射诱变创制是利用60Co-γ 射线照射种子,引起种子内染色体重组、畸变,产生遗传变异,从而形成水稻新种质、新材料,进而配制水稻新品种的一種高科技技术。60Co-γ 射线辐照诱变的频率高,可形成自然界没有的性状和类型,打破性状间的紧密连锁,促进基因重组,有些变异性状稳定快,可以在较短的时间内获得罕见的种质材料和基因资源。来自西非地区尼日利亚的农家栽培品种SIPI 具有耐瘠薄、分蘖强、耐高温等优良特性,但其稻穗较小,株型松散,限制了该品种的育种应用。本课题组在前期研究中利用60Co-γ 射线辐照处理SIPI 干种子,辐射剂量为300 Gy,随后连续自交多代筛选,最终获得一个稳定遗传的水稻亮绿叶突变体bgl-2。本研究通过对突变体bgl-2 进行田间表型和细胞学结构观察,并进行温度敏感试验,同时构建F2 分离遗传群体进行BGL-2 基因遗传模式分析,以期为后续BGL-2 基因的克隆和功能分析奠定基础。
1 材料与方法
1.1 材料
突变体bgl-2 是利用60Co-γ 射线300 Gy 辐照来自尼日利亚的籼稻品种SIPI 干种子,在M2 代,田间鉴定出亮绿叶突变表型植株,随后对突变植株连续自交多代获得稳定遗传的一个水稻亮绿叶突变体材料。2020 年春季种植野生型SIPI 和突变体bgl-2,地点位于海南省儋州市中国热带农业科学院水稻综合试验基地。种植规格为20 cm×20 cm,单株插秧,田间水肥、病虫管理参照当地常规大田管理方法。
1.2 方法
1.2.1 F2分离群体构建 2020 年春季种植野生型SIPI 和突变体bgl-2,抽穗期以突变体bgl-2 为母本,人工去雄,授以野生型SIPI 花粉,授粉25 d后收获杂交种F1。2020 年夏季分别种植野生型SIPI、突变体bgl-2 和F1,观察F1 植株表型,排除杂株后自交收获F2 种子。2021 年春季分别种植野生型SIPI、突变体bgl-2 和F2,获得分离遗传群体,田间调查F2 群体单株表型,计算分离比例。
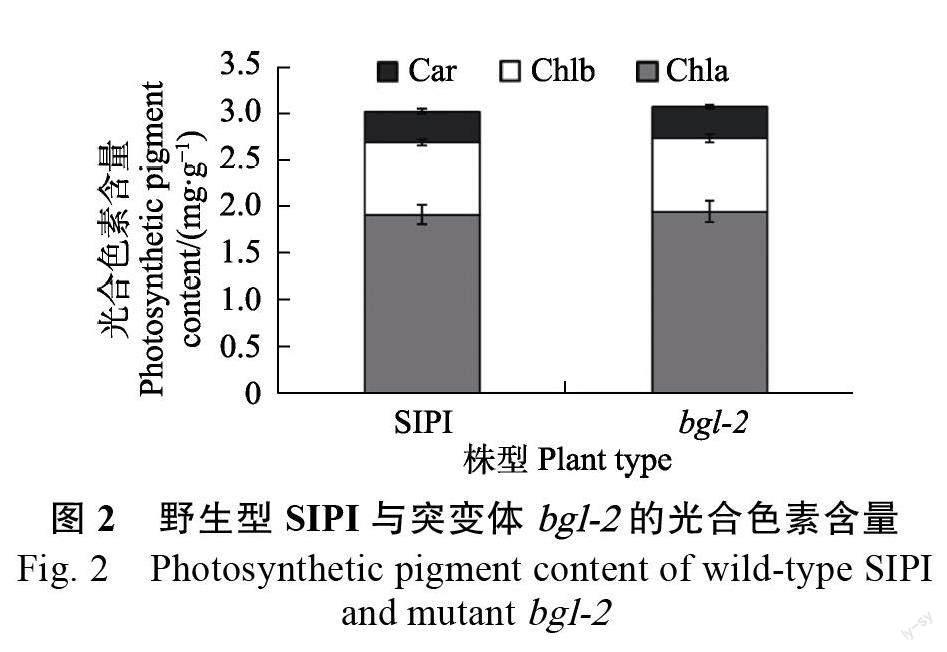
1.2.2 光合色素含量测定 分别取野生型SIPI 和突变体bgl-2 分蘖期剑叶叶片,测定光合色素含量,测定方法参照LI 等[7]的方法。光合色素提取完毕,利用DU800 紫外可见分光光度计,在黑暗条件下,测定上清液的吸光值(波长为470、649、665 nm)。并根据以下公式计算光合色素含量:
叶绿素a 含量:Chla(mg/g)=13.95D665–6.88D649
叶绿素b 含量:Chlb(mg/g)=24.96D649–7.32D665
类胡萝卜素含量:Car(mg/g)=(1000D470–2.05Chla–114Chlb)/245
细胞色素含量(mg/g)=(叶绿素浓度×提取液体积×稀释倍数)/样品鲜重
1.2.3 叶绿体超微结构的透射电镜观察 分别取分蘖期野生型SIPI 和突变体bgl-2 剑叶,横切成数段,每段约2 mm,置于3%的戊二醛固定液,抽气,室温固定12 h,4 ℃保存。将样品送至中国热带农业科学院环境与植物保护研究所电镜实验中心进行样品制备和透射电镜观察。
2 结果与分析
2.1 水稻亮绿叶突变体bgl-2 的表型鉴定
与野生型SIPI 比较,突变体bgl-2 在田间分蘖期表现为明显的叶片亮绿色(图1),此外还伴随着株高变矮,突变体bgl-2 株高为(91.10±3.13)cm,而野生型SIPI 的株高为(107.30±3.93)cm,二者之间存在显著差异(表1)。调查叶片形态发现,突变体bgl-2 的剑叶和倒二叶的叶尖处均表现出弯折,分别测定野生型和突变体bgl-2 的剑叶长度,结果发现野生型SIPI 的剑叶长度为(25.70±0.87)cm,而突变体bgl-2 的剑叶长度为(20.90±0.53)cm,显著短于野生型。拷种发现,突变体bgl-2的千粒重、粒长和粒宽均显著小于野生型SIPI,但长宽比在二者之间无显著差异(表1)。这些结果表明BGL-2 基因可能存在一因多效的效应。
2.2 突变体bgl-2 的光合色素含量测定
大多数造成叶色突变的基因直接或间接影响叶绿素的代谢过程,从而致使叶片叶绿素含量发生变化。为了了解突变体bgl-2 的叶绿素含量是否发生变化,在田间分蘖期分别取野生型SIPI 和突变体bgl-2 剑叶叶片,测定光合色素含量,结果如图2所示,二者之间无显著差异,说明突变体bgl-2 的叶色亮绿表型不是因为光合色素含量变化导致的,可能是BGL-2 基因突变引起其他机制变化导致的。
2.3 突变体bgl-2 的叶绿体超微结构观察
为了了解BGL-2 基因的突变是否影响到叶绿体的发育,本研究通过透射电镜观察分蘖期野生型SIPI 和突变体bgl-2 剑叶叶绿体的超微结构。如图3A 与图3D 所示,突变体bgl-2 的叶绿体数目与野生型SIPI 相比无显著差异。然而突变体bgl-2 的细胞结构出现异常。野生型SIPI 细胞壁厚度为(222.20±6.26)mm,而突变体bgl-2 的胞壁厚度增厚至(280.20±14.20)mm,但其细胞外膜厚度显著减小至(31.30±5.29)mm,且内膜几乎退化,而野生型SIPI 的细胞外膜厚度为(102.80±5.94)mm(表2)。此外突变体bgl-2 的类囊体片層结构显著减少(图3B、图3C、图3E 和图3F),说明突变体bgl-2 的细胞结构不正常,BGL-2 基因的突变影响其细胞结构和叶绿体的正常发育。
2.4 突变体bgl-2 的遗传模式分析
突变体bgl-2 与野生型SIPI 杂交获得的F1 代植株叶片表现正常绿色,剑叶和倒二叶形态正常,而自交后的F2代群体分离出叶色差异明显的正常绿叶苗和亮绿叶苗。卡方测验结果表明,F2 代群体中正常苗与亮绿叶苗的分离比例符合3∶1(表3),说明该突变性状受1 对隐性单基因控制。
3 讨论
水稻叶色突变体是一种出现频率较高的突变体类型,常见的有黄绿、浅绿、条纹、黄化、白化、白化转绿、黄化转绿等类型[8-12]。然而水稻叶片亮绿突变体比较少见,已报道的只有2 个[4]。水稻叶色表型多由一对隐性单基因控制,遗传行为简单。目前已报道的与叶绿素含量相关的基因有150 多个,其中已被克隆的叶色基因至少有30 个,这些基因主要分为叶绿素合成和降解途径的基因、叶绿体发育途径的基因以及其他途径相关基因等[13-16].
YOO 等[17]最先报道的突变体bgl 由于近轴和远轴叶表面均无乳头状结构的小角质乳突(smallpapillae, SP)导致绿光更多的直接反射而减少扩散,所以叶片呈现出亮绿色。但突变体bgl 的叶绿素含量和叶绿体结构与野生型相比,并无显著差异。图位克隆结果表明,bgl 位点是一个编码由11 个成员组成的水稻OsRopGEFs 家族之一OsRopGEF10。与SP 在叶表皮上启动的时间一致,OsRopGEF10 在叶鞘出苗前的新发育叶片中表达最强。随后WANG 等[18]报道另外一个新的水稻亮绿突变体bgl11,不同于突变体bgl1,其叶绿素含量显著低于野生型,且在成熟期,其单株有效穗数和结实率也显著低于野生型,但其他主要农艺性状与野生型相比无差异。基因图位克隆和测序结果表明,BGL11 是一个位于水稻第11 号染色体长臂上的基因LOC_Os11g38040。突变体bgl11 在该基因的编码区存在9 个bp 片段缺失。将LOC_Os11g38040 基因转到突变体bgl11 中,其叶片颜色恢复正常绿色。本研究的bgl-2 是一个辐射诱变获得的水稻亮绿叶突变体,但表型不同于bgl和bgl11,其不仅在叶片颜色上呈亮绿色,叶片形态也发生了变化,剑叶和倒二叶叶尖处均表现出弯折。此外在细胞结构上也发生了变化,细胞壁增厚,细胞膜变薄甚至退化,说明BGL-2 位点有可能不同于OsRopGEF10 和BGL11,是一个新的叶色亮绿基因。遗传模式分析表明突变体bgl-2由一对隐性单基因控制,下一步试验将进一步对BGL-2 进行基因定位及功能分析。
猜你喜欢
中国水稻科学(2021年6期)2021-11-18 12:06:30
语文世界(小学版)(2021年5期)2021-06-04 16:30:11
山东林业科技(2019年2期)2019-06-03 10:10:58
浙江农业科学(2019年1期)2019-01-25 05:25:34
天然产物研究与开发(2018年4期)2018-05-07 06:47:55
山东农业科学(2018年3期)2018-03-29 10:27:08
扬子江(2016年1期)2016-05-19 22:06:26
广西林业科学(2016年1期)2016-03-20 05:33:01
东北农业大学学报(2014年1期)2014-01-14 09:00:42
食品科学(2013年6期)2013-03-11 18:20:13
