
棉秆固体发酵中木质素降解与酶活性变化的关联分析
2023-07-13 03:55李佩琪孙庆培王志慧秦新政樊永红
新疆农业科学 2023年6期
李佩琪,孙庆培,王志慧,秦新政,樊永红
(1.新疆大学生命科学与技术学院/新疆生物资源基因工程重点实验室,乌鲁木齐 830046;2.新疆农业科学院微生物应用研究所/新疆特殊环境微生物实验室,乌鲁木齐 830091)
0 引 言
【研究意义】2020年新疆棉花产量达516.1×104t,占全国棉花总产量87.3%,棉花秸秆作为主要的棉花副产品,总量很大[1]。当前处理棉秆的方法主要是直接焚烧后还田和就地粉碎掩埋于棉田[2]。棉秆有机物含量高,同时富含氮、磷、钾等营养元素,通过固体发酵,可以转化为有机基质[3],其中木质素和纤维素的降解程度代表了有机基质的腐熟程度[4]。木质素作为一种物理屏障嵌合在纤维素和半纤维素中,形成致密的基质,阻碍了秸秆的生物降解与利用[5-6]。秸秆腐熟过程实质是微生物反应过程,而微生物反应主要依赖于其分泌的相关降解酶作用。木质素降解酶系统决定着木质素降解的途径、速度和程度。目前已知的木质素降解酶主要包括:木质素过氧化物酶(Lignin peroxidases/LiP)、锰过氧化物酶(Manganese perxidases/MnP)和漆酶(Laccase/Lac)[7-8]。分析棉秆固体发酵中木质素的降解与木质素降解酶活性变化关联性,对研究木质素降解机理有重要意义。【前人研究进展】目前,其他类型的秸秆用于生产有机基质已经有了很多成功的实例[9-11],在稻草秸秆堆肥过程中,纤维素和半纤维素均在高温期降解明显,堆肥结束时分别降解10.07%和3.20%[12]。纤维素和半纤维素在发酵初期降解最强烈,木质素的降解率在发酵9 d后才略微升高,15 d后半纤维素、纤维素、木质素含量分别下降82.49%、97.20%和20.12%[13]。在含有稻草和鸡粪的堆肥研究中发现,LiP活性在堆肥的最初7 d逐渐升高,随后开始下降[14]。在小麦秸秆堆肥中添加腐生真菌使Lac、MnP活性显著提高至2 000和220 U/L,且Lac活性和MnP活性之间存在明显的相关性[15],关于木质素降解酶的研究更多集中在木质素降解酶相关基因结构[16-17]、表达调控机理[18-19]、以及酶学性质[20-21]等。【本研究切入点】由于棉秆理化结构有其特殊性,实际应用中有关棉秆木质素降解关键酶作用规律及其与木质素降解的关联研究较少。需分析棉花秸秆经固体发酵转化为有机基质过程中的木质素降解与木质素降解酶活性的关联性。【拟解决的关键问题】研究木质素在基质化过程中降解的生物学特性,对优化棉秆生产有机基质工艺和提高木质素降解效率提供重要依据和参考。
1 材料与方法
1.1 材 料
棉花秸秆来源于新疆乌鲁木齐市,将棉秆自然晾干粉碎成长度约2~3 cm。
1.2 方 法
1.2.1 棉秆固体发酵
取5 kg粉碎干燥的棉秆,加入4 kg的水,搅拌均匀,在40目透气尼龙袋中进行腐熟发酵,根据温度变化,分别在第1、3、5、14、18、25 d时从堆体不同部位取样后混合,低温粉碎后用于测定各种指标。在堆体内外放置温度计,记录环境温度和发酵温度变化。
1.2.2 固体发酵不同阶段棉秆失重率
棉秆失重率以质量分数ωx计,数值以%表示。
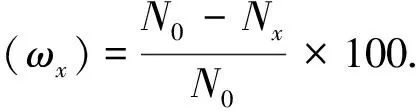
式中,N0为发酵前棉秆干重;NX为某一发酵时间的棉秆干重。
1.2.3 固体发酵不同阶段棉秆纤维素、半纤维素的含量
纤维素和半纤维素含量均按照NY/T3494-2019[22]测定。棉秆发酵样品干燥、粉碎,过40目筛,通过索氏抽提,先后用去离子水和95%无水乙醇充分去除样品中的水溶性、醇溶性游离组分。经过索氏抽提和干燥处理的样品进行硫酸水解,经浓硫酸酸化将样品中高聚合度的纤维素和半纤维素降解为低聚合度的寡糖,再通过稀硫酸将低聚寡糖进一步水解成单糖。样品的酸解溶液过砂芯坩埚滤器后,采用高效液相色谱方法测定其中单糖的含量,确定样品中的纤维素、半纤维素的含量。纤维素、半纤维素含量均以质量百分数(%)表示。

纤维素含量(Cel)=Z1.
半纤维素含量(Hem)=Z2+Z3+Z4+Z5.
式中,Ci为第i种单糖的浓度;V为滤液体积;F为脱水校正因子;木聚糖、阿拉伯聚糖用0.88校正,葡聚糖、甘露聚糖和半乳聚糖用0.9校正;Ri为单糖回收率;ω0为抽提前样品质量;ω1为抽提后样品质量;ωef为称取的不含抽提物的试样质量;Z1为试样中葡萄聚糖的含量;Z2为试样中木聚糖的含量;Z3为试样中半乳聚糖的含量;Z4为试样中阿拉伯聚糖的含量;Z5为试样中甘露聚糖的含量。
1.2.4 固体发酵不同阶段棉秆木质素含量
木质素含量按照NY/T3494-2019测定。总木质素含量为酸溶性木质素和酸不溶性木质素之和。样品的酸解滤液经过320 nm紫外检测吸光度,计算出酸溶性木质素含量。酸不溶性木质素则通过高温灼烧法,在马弗炉中575 ℃充分灼烧砂芯坩埚滤器收集的残渣,计算出酸不溶性木质素含量。以上二者之和即为样品中的总木质素含量。酸溶性木质素、酸不溶性木质素含量均以质量百分数(%)表示。
式中,m1为坩埚的质量;m2为坩埚和酸不溶性残渣的质量;m3为坩埚和灰分的质量;A为滤液在320 nm处的吸光值;N为滤液稀释倍数;L为比色皿厚度;ε为酸溶木质素在320 nm处吸收率;值为25 L/(g·cm)。
1.2.5 固体发酵不同阶段木质素降解酶活性
粗酶液的提取:取5 g发酵棉秆,加入50 mL缓冲溶液(0.1 mol/L酒石酸钠缓冲液(pH 4.5)和0.1 mol/L乙酸-乙酸钠缓冲液(pH 4.5),分别用于不同的木质素酶活分析),220 mg/kg振摇浸提1 h,在4℃下10 000 r/min离心10 min,取上清作为待测粗酶液。
锰过氧化物酶(MnP)的测定[23]:反应体系3 mL,0.1 mol/L酒石酸钠缓冲液(pH 4.5)2.52 mL,10 mmol/L的硫酸锰0.15 mL,10 mmol/L的2,6-DMP (2,6-二甲氧基苯酚) 0.15 mL,粗酶液0.15 mL,混匀,37℃水浴3 min,加入10 mmol/L过氧化氢0.03 mL启动反应,测定最初3 min内469 nm处吸光值的变化,469 nm处2,6-DMP氧化产物的摩尔消光系数为49 600 L/(mol·cm)。酶活定义为每分钟氧化1 μmol底物所需的酶量为一个酶活力单位(U)。
木质素过氧化物酶(LiP)的测定[24]:反应体系3 mL,0.1 mol/L酒石酸钠缓冲液(pH 3)1.5 mL,10 mmol/L的藜芦醇1 mL,粗酶液0.4 mL,混匀,25 ℃水浴3 min,加入10 mmol/L的过氧化氢0.1 mL启动反应,测定最初3 min内310 nm处吸光值变化,310 nm处藜芦醇氧化产物的摩尔消光系数为9 300 L/(mol·cm)酶活定义为每分钟氧化1 μmol底物所需的酶量为一个酶活力单位(U)。
漆酶(Lac)的测定[25]:反应体系3 mL,0.1 mol/L乙酸-乙酸钠缓冲液(pH 4.5)1.8 mL,0.5 mmol/L的ABTS(2,2-联氮-二(3-乙基-苯并噻唑-6-磺酸))0.2 mL,粗酶液1 mL,混匀,28 ℃水浴3 min,测定最初3 min内420 nm处吸光值变化,420 nm处ABTS氧化产物的摩尔消光系数为36 000 L/(mol·cm)酶活定义为分钟氧化1 μmol底物所需的酶量为一个酶活力单位(U)。

式中,n为测定时酶液的稀释倍数;ε为摩尔消光系数;V总为反应总体积;V酶为反应中加入的粗酶液体积。
1.3 数据处理
采用Microsoft Excel 2019、Orgin 2021软件进行数据整理、绘图。在SPSS 26.0软件中利用Pearson相关性分析法、主成分分析法对数据进行分析。
2 结果与分析
2.1 固体发酵不同阶段温度变化
研究表明,整个发酵过程中,发酵温度都呈现出先上升后下降的趋势,发酵前期堆体快速升温,在温度达到50℃后进入高温期,最高温度达到62℃,高温维持2~3 d后开始降温,降至室温后进行翻堆,翻堆后温度再次上升至54℃,之后温度再次下降;经多次翻堆后发酵温度不再上升,静置发酵直至结束。将整个发酵阶段分为升温期(0~3 d),高温期(4~6 d),熟化期(8~25 d)。取发酵第1 d(初始阶段)、3 d(升温期)、5 d(高温期)、18 d(熟化期)、25 d(末期)发酵样品作为研究材料。图1

图1 棉秆固体发酵过程中堆体温度变化
2.2 固体发酵不同阶段棉秆木质素、纤维素及半纤维素含量测定
研究表明,整个固体发酵过程中棉秆失重明显,尤其是进入熟化期后,棉秆失重率显著提高(P<0.05)。发酵之前棉秆木质素、半纤维素和半纤维素含量分别为31.51%、29.32%和15.17%,升温期时其含量几乎没有改变,高温期才逐渐开始减少,此时木质素、半纤维素和半纤维素含量分别下降至28.36%、11.48%和30.60%。在发酵结束后木质素含量下降至25.38%,但在整个发酵过程中含量总体变化较小;纤维素和半纤维素含量变化显著(P<0.05),分别下降至18.18%和7.75%。表1

表1 固体发酵棉秆中纤维素、半纤维素和木质素含量
木质素降解速率在高温期过后显著升高,在熟化期持续升高,直到发酵结束时达到最大13.84 g/d。半纤维素和纤维素降解速率分别在升温期和高温期达到最高,为26和23.95 g/d。图2

注:IS:初始;WP:升温期:HP:高温期;MP:熟化期;FS:末期,下同
2.3 固体发酵不同阶段木质素降解酶活性测定
研究表明,发酵初期堆体温度逐渐升高,MnP活性在发酵第3 d上升至4.24 U/g;进入高温期后酶活下降至3.45 U/g,随着堆体温度下降,MnP活性逐步上升;进入熟化期后,MnP活性持续上升,在发酵结束时酶活达到最高,为12.91 U/g。图3

图3 棉秆发酵过程中锰过氧化物酶活性变化
LiP活性在发酵过程中呈先升后降再上升的趋势,与MnP活性变化趋势一致。LiP活性在发酵第3 d由初始的3.81 U/g上升至22.14 U/g,进入高温期后酶活下降至14.65 U/g;随着堆体温度下降,LiP活性逐步上升;进入熟化期,LiP活性持续上升,在发酵结束时酶活达到最高,为55.28 U/g。图4

图4 棉秆发酵过程中木质素过氧化物酶活性变化
在发酵初期未检测到Lac活性,直至高温期才出现Lac活性,此时Lac活性为0.02 U/g;随后Lac活性持续上升,在发酵结束时酶活达到最高,为0.19 U/g。图5

图5 棉秆发酵过程中漆酶活性变化
2.4 固体发酵过程中木质素降解酶活性与木质素降解相关性
研究表明,LiP、MnP、Lac活性与木质素降解速率呈极显著正相关(P<0.01),随着这几种酶活性升高,堆体中木质素降解速率不断加快,木质素降解酶对木质素的降解影响极大。3种木质素降解酶活性与纤维素降解速率也呈显著相关(P<0.01),与半纤维素降解速率相关性不显著,木质素降解酶活性对纤维素的降解影响较大,对半纤维素的降解影响较小。图6

注:A锰过氧化物酶;B木质素过氧化物酶;C漆酶
发酵过程中3种木质素降解酶活性之间也存在着一定的相关性。在LiP、MnP、Lac中,两两之间均呈极显著正相关(P<0.01),在发酵过程中3种木质素降解酶呈现相互促进作用。但LiP、MnP、Lac活性与温度波动相关性差异不显著,3种酶活性的变化与温度有关,但受温度的影响较小。 表2

表2 木质素降解酶活性变化与木质素降解相关性
2.5 棉秆固体发酵过程中木质素降解的主成分
研究表明,前2个主成分所包含的信息量分别为62.591%、22.800%,累积占总信息量的85.391%,可以代表所有被测变量的绝大多数信息,主成分1和主成分2对棉秆固体发酵过程中木质素降解的影响最大,提取这2种主成分进一步做负荷矩阵分析。
主成分1反映了与固体发酵中木质素降解酶相关的指标,其中MnP的因子载荷为0.957、LiP的因子载荷为0.931和Lac的因子载荷为0.909,3种木质素降解酶活性对主成分1的影响最大。随着酶活性的增加,固体发酵中木质素降解速率升高,木质素含量逐渐减少。主成分2反映了与固体发酵中木质素降解程度有关的指标,其中半纤维素降解速率的因子载荷为0.918、温度的因子载荷为0.898、纤维素降解速率的因子载荷为0.690,木质素降解程度与发酵温度、纤维素和半纤维素降解速率密切相关。由于主成分1对固体发酵中木质素降解的方差贡献率显著高于其他主成分,而MnP活性在主成分1中因子载荷最高,MnP活性是影响棉秆固体发酵过程中木质素降解的最主要因子。表3~4,图7

表3 总方差的解释

表4 主成分因子负荷矩阵

图7 主成分分析碎石图
3 讨 论
3.1 棉秆固体发酵中木质素的降解
温度作为有机基质腐熟的重要参数,影响着秸秆腐熟过程中微生物的代谢活动强弱[26]。作为典型的秸秆固体发酵过程,研究中棉秆固体发酵过程也可根据温度分为3个阶段:升温期,高温期,熟化期[27]。在升温期时,简单有机物如单糖、蛋白质和淀粉等首先被微生物分解,所产生的热量使堆体温度升高[28];随后进入高温期,在此期间堆温升高到50℃以上,耐热微生物占主导地位,可将纤维素、半纤维素和果胶分解为微生物容易利用的碳水化合物[29]。与纤维素和半纤维素不同,木质素作为难降解的成分,其含量的变化主要发生在温度低于50℃的发酵后期[30-32],即熟化期。在此阶段木质素开始被中温微生物降解,还有许多耐高温的微生物,如芽孢杆菌等在堆体温度开始下降时就能降解木质素,木质素主要在高温阶段后期和熟化期时被降解,期间没有出现明显的降解减缓过程[33]。研究显示,木质素降解速率在高温期后显著提升,发酵结束时达到最高,为13.84 g/d,经过固体发酵棉秆中木质素含量下降6.13%,纤维素和半纤维素含量分别下降11.13%和7.42%,与前人研究的水稻秸秆和玉米秸秆腐解规律相似[34]。
3.2 棉秆固体发酵中木质素降解酶的活性变化
木质素、纤维素和半纤维素降解酶活性的动态变化是秸秆发酵的过程中重要的生物学指标[35]。研究中LiP和MnP活性表现出相同的变化趋势,在发酵初期温度上升,微生物代谢活力和代谢水平开始逐渐升高,此时微生物主要利用含量有限的、易降解的有机物进行生长繁殖,无法产生大量酶类[36]。进入高温期后,由于有机质降解酶活性与微生物代谢密切相关,高温期微生物数量减少,酶活性下降。与纤维素和半纤维素相比,木质素更难被微生物分解利用,进入熟化期后,堆体温度下降,嗜温微生物重新活跃起来,此时开始分解转化堆料中残余的难降解有机物,木质素降解酶活性显著升高[37]。在含有松树皮和麦秆的堆肥中,LiP活性在较长堆肥时间后才会显著升高[38],而在木材堆肥过程中,也检测出LiP活性在发酵后期升高[39]。不同于LiP和MnP的活性变化,Lac活性在发酵初期很低,在发酵进入高温期后才表现明显,这可能与过氧化物酶作用下木质素降解产物诱导其生物合成有关[40]。研究表明,在整个发酵过程中,MnP、LiP、Lac活性均随着发酵时间的延长逐步上升,在熟化期酶活明显提高,在发酵结束时木质素降解酶活性达到最高。3种木质素降解酶中LiP的活性最高,由于LiP的底物主要是芳香环化合物,棉秆固体发酵过程中易降解的木质素可能以芳香环化合物为主。
3.3 棉秆固体发酵中木质素降解及酶活性变化的关联分析
秸秆发酵中木质素降解主要依靠木质素降解酶,它能够使木质素结构单体间之间的C-O键或C-C键断裂,并对侧链进行一定的修饰,使木质素最终分解为低分子物质[41]。Lac、MnP、LiP都是直接作用于木质素的酶,在木质素降解过程中发挥着关键作用[42]。其他类型的酶也对木质素的降解具有协同作用,例如葡萄糖氧化酶氧化葡萄糖的同时为LiP提供过氧化氢,诱导木质素降解,而在LiP作用下的木质素降解产物能够诱导Lac的生物合成[43]。碳源浓度的降低能够诱导菌株产生木质素降解酶[44],Lac和LiP活性也会随着秸秆腐熟过程中碳氮比的降低而增加[45]。研究的木质素降解及酶活性变化的关联分析可以看出,木质素降解趋势与固体发酵不同时期的降解酶活性的变化趋势基本相同,3种木质素降解酶活性之间显著性相关,3种木质素降解酶在发酵过程中相互协同降解木质素。而温度和MnP、LiP、Lac活性没有显著的相关性,可能是因为木质素降解酶活性受其他因子的影响程度更大。MnP活性对棉秆固体发酵的木质素降解的影响最大。可以通过固体发酵的过程控制,例如添加MnP的生产菌株,有针对性的提高发酵后期MnP的活性,以提高棉秆木质素的降解速率,促进棉秆固体发酵转化的效率。
4 结 论
在棉秆固体发酵过程中,棉秆木质素含量由31.51%显著降低至25.38%(P<0.05),其含量的变化主要发生在熟化期。木质素降解速率也在高温期后显著升高,熟化期持续升高直至发酵结束时达到最高13.84 g/d。在整个发酵过程中,MnP、LiP、Lac活性均随着发酵时间的延长逐步上升,在熟化期酶活明显提高,发酵结束时活性达到最高,分别为12.91、55.28和0.19 U/g。3种木质素降解酶活性均与木质素降解速率呈显著正相关(P<0.01),木质素降解酶活性的升高有利于木质素的降解。3种木质素降解酶活性之间也呈显著正相关(P<0.01),在发酵过程中相互协同降解木质素,其中MnP占据主要的地位。
猜你喜欢
安徽农业科学(2022年14期)2022-08-04
中国农机化学报(2021年7期)2021-08-13
纺织科技进展(2021年3期)2021-06-09
陶瓷学报(2021年1期)2021-04-13
新疆农机化(2020年1期)2020-03-26
上海包装(2019年8期)2019-11-11
天津造纸(2016年1期)2017-01-15
中国造纸学报(2015年1期)2015-12-16
中国塑料(2014年4期)2014-10-17
应用化工(2014年11期)2014-08-16
