
玉米ZmCDPK22基因在干旱胁迫下的表达分析
2023-07-13 03:53邵盘霞邵武奎郝晓燕高升旗李建平胡文冉黄全生
新疆农业科学 2023年6期
邵盘霞,赵 准,邵武奎,郝晓燕,高升旗,李建平,胡文冉,黄全生
(新疆农业科学院核技术生物技术研究所/新疆农作物生物技术重点实验室,乌鲁木齐 830091)
0 引 言
【研究意义】在植物生长过程中会遇到干旱、盐碱、低温等非生物逆境胁迫,严重影响植物的生长发育和制约农业生产。干旱是制约作物生产的主要非生物胁迫之一,利用分子生物学方法和手段探索作物的耐旱机制,改善作物本身的抗旱能力,对作物抗旱品种的培育具有重要意义。【前人研究进展】应对逆境,植物在生长过程中会调节生理生化代谢途径,激素信号转导及调控基因表达等来抵御胁迫,提高逆境胁迫的耐受性[1, 2]。Ca2+在介导生物和非生物胁迫信号转导中起着重要作用,细胞中的钙信号通过钙敏感蛋白,如钙调蛋白,钙调磷酸酶B蛋白(Calcinerium-B Like,CBL)和钙依赖蛋白激酶(Calcium-dependent protein kinase,CDPK)等调节相关基因的表达,从而应对环境胁迫[3, 4]。CDPK作为钙结合蛋白之一,其家族的许多成员在植物抵抗干旱胁迫的调控反应中发挥着重要作用。前人研究报道CDPK基因具有提高作物抗旱性的作用。拟南芥CDPK相关基因通过调节气孔参与干旱胁迫响应,过表达AtCPK4、AtCPK8、AtCPK10和AtCPK11可以显著提高转基因植物的抗旱性[5-7]。OsCDPK4在水稻的耐盐性和干旱胁迫中发挥着重要作用,过表达显著增强水稻对盐和干旱胁迫的耐受性[8];OsCDPK9通过调节气孔关闭和渗透调剂提高水稻干旱胁迫的耐受性[9]。ZoCDPK1在参与生姜响应干旱胁迫的信号通路中发挥正向调节作用,在烟草中过表达该基因,提高了烟草对干旱胁迫的耐受性[10]。玉米是我国重要的作物之一,是重要的饲料和工业原料,其产量和品质受干旱胁迫的制约。在玉米中,已发现了40个CDPKs基因[11]。【本研究切入点】ZmCDPK1参与玉米低温胁迫响应,在低温信号转导中期负调节作用[12];ZmCPK11由机械损伤诱导表达,在酶促反应和转录水平受亚麻酸和茉莉酸甲酯调节[13];虽然已经克隆了ZmCDPK、ZmCDPK17、ZmCDPK22、ZmCDPK28和ZmCDPK34等基因,但是在植物中的功能尚未研究[12]。因此探索ZmCDPK22是否响应干旱、盐碱等非生物胁迫及它们发挥的作用很有必要。【拟解决的关键问题】以ZmCDPK22作为研究对象,从生物信息学和干旱胁迫表达两方面对该基因的功能进行判断,为研究该基因的功能提供一定的参考依据。
1 材料与方法
1.1 材 料
以玉米自交系B73(实验室保存及繁种)为材料,以叶片含水量模拟干旱胁迫条件,在玉米四叶时期测定玉米含水量,采用烘干称重法对相应叶片测定叶片含水量。用精度为0.001 g的天平对鲜叶称重,称重后将叶片放入105℃恒温箱杀青15 min,于80℃下烘干至恒重后称其干重,计算各叶片含水量,叶片含水量=(鲜重-干重)/鲜×100%。
1.2 方 法
1.2.1ZmCDPK1生物信息学分析
通过ExPASy的工具ProtParam(https://web.expasy.org/protparam/)预测蛋白理化性质;应用在线工具DeepTMHMM(https://services.healthtech.dtu.dk/service.php/DeepTMHMM)预测跨膜结构;SOPMA(http://npsa-prabi.ibcp.fr/)用于预测ZmCDPK22二级结构;应用SWISS-MODEL((https://swissmodel.expasy.org/))预测其三级结构;Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)用于预测亚细胞定位;SignalP3.0 Server (http://www.cbs.dtu.dk/services/SignalP/)用于分析信号肽;通过NCBI CDD(https://www.ncbi.nlm.nih.gov/cdd/)分析该基因的保守结构域;MEME(http://meme-suite.org/)预测保守基序;使用STRING(https://string-db.org/)在玉米数据库中进行ZmCDPK22互作蛋白预测;应用MEGA-X进行多序列比对和构建进化树;通过NCBI网站下载ZmCDPK22基因起始密码子上游2 000 bp序列在PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)网站进行启动子区顺式作用元件分析。
1.2.2ZmCDPK22表达模式
按照植物总RNA提取试剂盒说明提取玉米叶片RNA,逆转录后得到的cDNA。以cDNA为模板进行实时荧光定量PCR分析。根据ZmCDPK22基因的CDS(coding sequence)序列设计引物(ZmCDPK22-qF:AGGTCATATTGACTTCGCATCT;ZmCDPK22-qR:GCAACCTTCTTAAGCTTGTTCA)。以ZmUBI(ZmUBI-qF:TGGTTGTGGCTTCGTTGGTT;ZmUBI-qR:GCTGCAGAAGAGTTTTGGGTACA)作为内参基因,使用BIO-RAD CFX96 Real-Time System qPCR仪进行实时荧光扩增。应用2-ΔΔCT方法分析ZmCDPK22基因的干旱胁迫表达模式。
2 结果与分析
2.1 ZmCDPK22基因序列及蛋白理化性质分析
研究表明,ZmCDPK22基因全长为2 216 bp仅含有一个外显子,CDS序列长1 620 bp,编码539个氨基酸。ZmCDPK22分子式为C2675H4217N779O798S21,分子量为60 731.91,等电点为6.44;该蛋白中丙氨酸(Ala,A)、谷氨酸(Glu,E)和精氨酸(Arg,R)的含量较高,分别为45个(8.3%)、43个(8.0%)和41个(7.6%),不含吡咯赖氨酸(Pyl,O)和硒代半胱氨酸(Sec,U);亲水性系数为-0.602,不稳定性指数为50.15,说明该蛋白为亲水性不稳定蛋白。蛋白二级结构显示该蛋白由ɑ-螺旋(41.93%)、无规则卷曲(39.89%)、延伸链(9.28%)和β-折叠(8.91%)组成(图1A);蛋白三级结构预测显示与该蛋白同源基因三级结构最高相似度为37.53%。亚细胞定位预测该蛋白定位于细胞核,无信号肽和跨膜结构域。图1
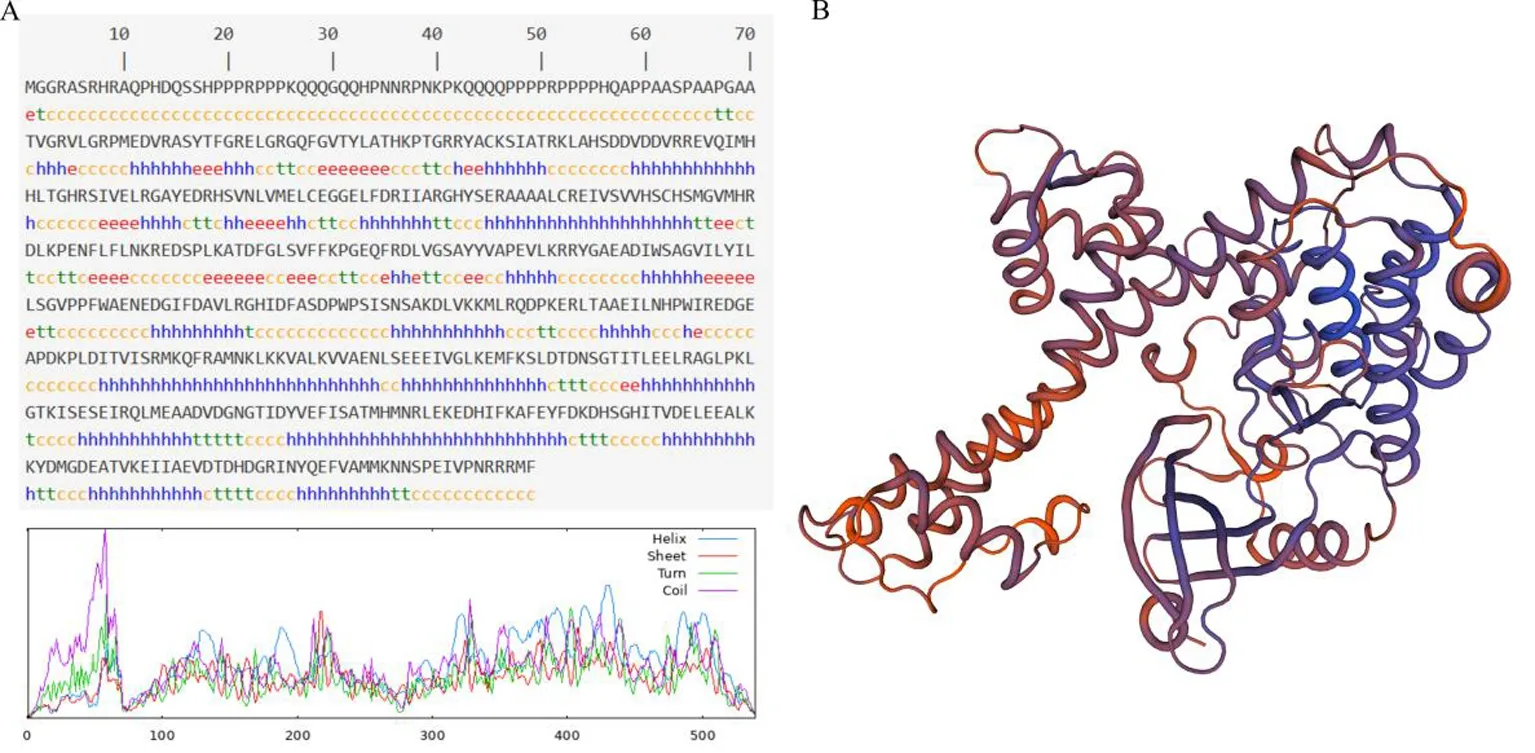
图1 ZmCDPK22蛋白质二级结构(A)与三级结构(B)
2.2 ZmCDPK22蛋白结构域和基序预测
研究表明,ZmCDPK22含STK_CAMK结构域,属于PK_like亚家族。有18种保守基序,分别为Motif1~Motif18;ZmCDPK22、ZmCDPK15和DoCDPK3不含Motif18基序且先后顺序都相同。图2,图3
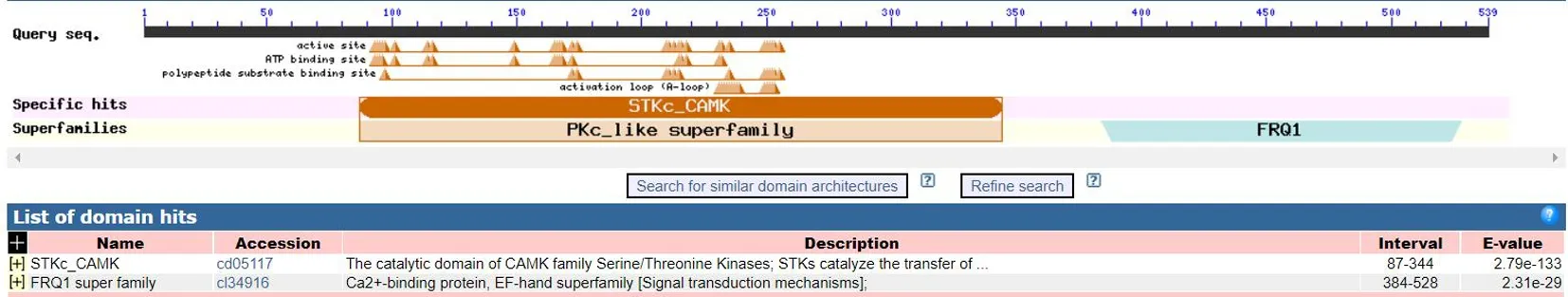
图2 ZmCDPK22的蛋白结构域
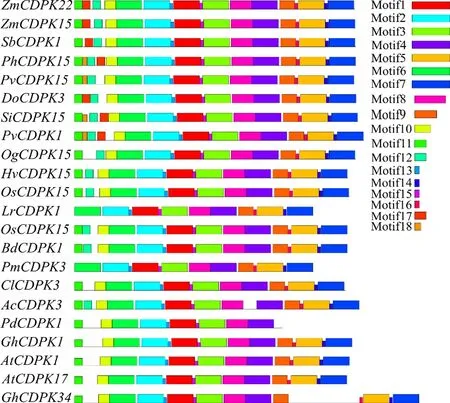
图3 ZmCDPK22的保守基序
2.3 蛋白互作预测
研究表明,ZmCDPK22与10个蛋白互作,与丝氨酸/苏氨酸蛋白磷酸酶(GRMZM2G148539_P02)的互作最强,互作系数为0.618;与呼吸爆发氧化酶同源蛋白相关基因(RbohC和rboh2)互作较弱,互作系数为0.600。图4
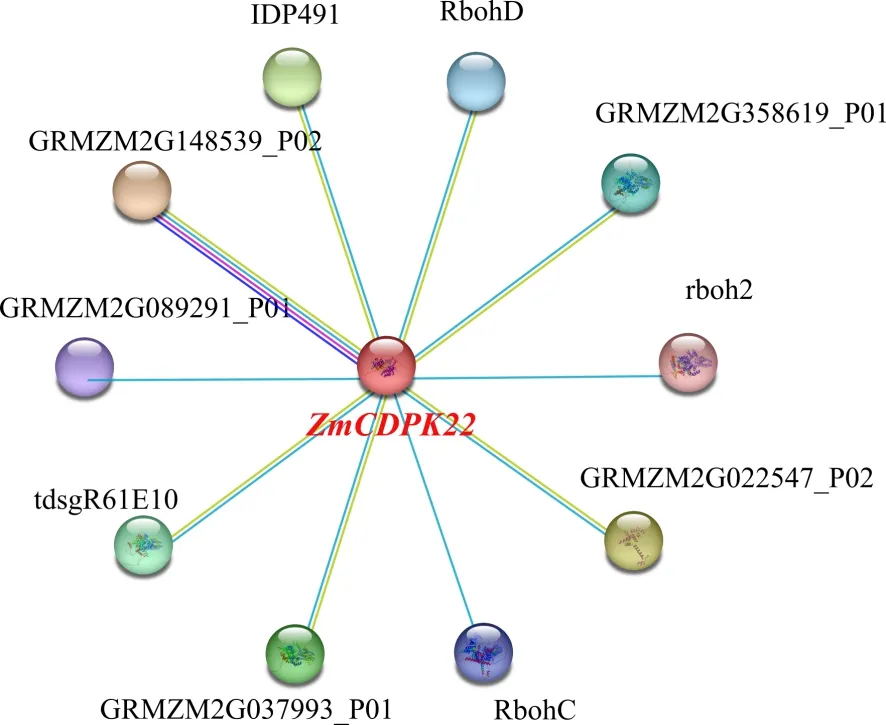
图4 ZmCDPK22蛋白互作网络模型
2.4 进化树与启动子区域顺式作用元件
研究表明,ZmCDPK22与ZmCDPK15的蛋白序列相似度最高,为98.89%,与拟南芥AtCDP17和棉花GhCDPK34的蛋白序列相似度比较低,分别为69.25%和64.62%。利用MEAG-X的邻接法构建玉米、拟南芥、棉花、小麦和水稻等植物相关CDPK基因的进化树,ZmCDPK22与玉米ZmCDPK15和高粱SbCDPK1的亲缘关系最近,与拟南芥AtCDPK17和棉花GhCDPK34的亲缘关系较远。图5

图5 ZmCDPK22系统进化树
ZmCDPK22基因启动子区含有31种顺式作用元件,除含有TATA-box和CAAT-box等核心启动元件之外,还含有激素相关响应元件、光响应元件、厌氧诱导元件、低温响应元件及分生组织表达等相关元件。在激素响应相关元件中,茉莉酸甲酯响应元件共有6个,脱落酸响应元件有2个。表1

表1 ZmCDPK22启动子顺式作用元件
2.5 ZmCDPK22对干旱胁迫的响应
研究表明,叶片含水量为87%时ZmCDPK22的表达量记为1,在不同含水量情况下,ZmCDPK22的表达量不同。与叶片含水量87%相比,ZmCDPK22的相对表达量随着叶片含水量的降低而逐渐降低,在叶片含水量为78%时相对表达量最高,在含水量为33%时相对表达量最低。图6
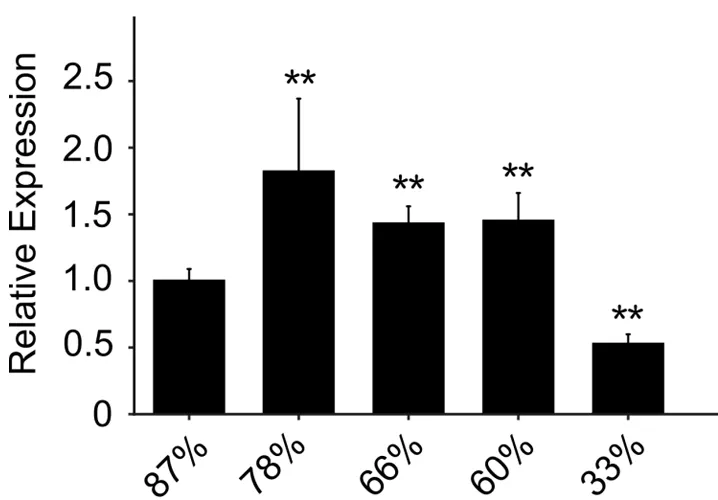
图6 ZmCDPK22在叶片不同含水量的相对表达量
3 讨 论
研究对ZmCDPK22的启动子区顺式作用元件分析表明,该基因包含2个关键的启动子元件,将转录起始位点与RNA聚合酶连接的TATA-box和调节基因转录效率的CAAT-box,ZmCDPK22可以正常转录。在对拟南芥和蒺藜苜蓿相关CDPKs基因的启动子区顺式作用元件的研究中发现一些AtCPKs和MtCDPKs参与植物激素和非生物胁迫信号的调节[14-17]。ZmCDPK22启动子区还含有多种激素及逆境响应元件,推测该基因可能在多种逆境胁迫响应中发挥着重要作用。
ZmCDPK22可能参与对生物和非生物胁迫的反应,尤其是对病原体的防御和干旱胁迫,研究中通过蛋白互作网络预测分析,发现ZmCDPK22与RBOH有关。RBOHs是植物体内活性氧生成的主要途径,在植物生长发育和逆境响应中发挥重要的作用[18]。在葡萄中,盐和干旱胁迫处理显著增强了VvRBOHA、VvRBOHB和VvRBOHC1的表达,外源ABA处理显著上调VvRBOHB的表达[19]。油菜RbohA、RbohD基因在响应低温、盐、PEG-6000模拟干旱胁迫过程中发挥作用[20]。研究中ZmCDPK22与多个Rboh蛋白互作,猜测其可能与Rboh基因有相似或相同的功能。
研究发现CDPK参与植物非生物胁迫反应,干旱[21]、盐[22]、光[23]等多种环境因子以及生长素[24]等都能引起CDPK基因的特异性表达。ZmCDPK9是CDPKs基因家族中一个新的成员并对干旱胁迫具有一定的应答反应[25]。ZmCDPK38基因响应干旱胁迫与盐胁迫,可能在干旱应答与盐胁迫应答反应中发挥一定的功能[26]。叶片含水量能直接反映作物生长发育的实际状况,是机理研究中反映作物水分盈亏程度的最佳指标,是作物耐旱能力综合作用的体现和监测作物受胁迫的重要参考[27, 28]。研究测定了ZmCDPK22基因在不同含水量叶片中的相对表达量,在含水量为78%时相对表达量最高,在含水量为33%时相对表达量最低,ZmCDPK22对干旱胁迫有一定响应。
4 结 论
从玉米中克隆得到ZmCDPK22基因,该基因编码539个氨基酸,相对分子量为60 731.91,等电点为6.44,无跨膜结构以及信号肽,该蛋白定位于细胞核;ZmCDPK22与10个蛋白存在互作,属于PK_like亚家族。其启动区含有大量的光响应和多种激素响应等元件。在不同含水量情况下,ZmCDPK22的表达量不同。与叶片含水量87%相比,ZmCDPK22的相对表达量随着叶片含水量的降低而逐渐降低,该基因对干旱胁迫具有一定响应。
猜你喜欢
今日农业(2020年20期)2020-12-15
今日农业(2020年17期)2020-12-15
世界农药(2019年4期)2019-12-30
森林工程(2018年4期)2018-08-04
时代农机(2018年11期)2018-03-17
上海农业学报(2016年2期)2016-10-27
现代工业经济和信息化(2016年2期)2016-05-17
电源技术(2016年9期)2016-02-27
电子工业专用设备(2015年4期)2015-05-26
汽车维修与保养(2015年8期)2015-04-17
