
中国水华蓝藻新记录属
——泽鞘丝藻属(Limnoraphis)
2023-07-13 02:28:32谢艳张荣臻张葵东吕兴菊张和肖鹏李仁辉程耀耿若真
河南师范大学学报(自然科学版) 2023年4期
谢艳,张荣臻,张葵东,吕兴菊,张和,肖鹏,李仁辉,程耀,耿若真
(1.温州大学 生命与环境科学学院,浙江 温州 325035;2.温州市珊溪水利枢纽管理中心,浙江 温州 325000;3.大理州洱海湖泊研究院,云南 大理 671000)
蓝藻分类学是认识和评价蓝藻生物多样性的重要方法,对于了解自然生物群落中蓝藻种群的功能非常重要.作为地球上最古老的生物类群之一,蓝藻具有漫长且复杂的进化历史,部分蓝藻化石的形态与现在的物种非常相似,是一类非常具有挑战性的分类群体[1-2].在蓝藻的原核特性被发现之前,由于蓝藻形态多样,传统的蓝藻分类系统主要以形态学为基础,遵循植物学命名法则进行命名和描述[3].随着分子生物学的发展及系统发育分析的出现,利用分子手段解决蓝藻的分类学问题已被越来越多的分类学家所接受,蓝藻的整个分类阶层(目、科、属、种)经历了广泛的重组和修订[4-5].在这个过程中,基于形态学、分子系统发育学和生态学等特征相结合的多相分析方法,逐渐成为研究蓝藻分类学的最佳方法[6-8].蓝藻分类学研究中多相方法的应用导致大量属种的修订和新属种的建立,为解决系统发育的单系分类问题提供了方法[5,9].
泽鞘丝藻属(Limnoraphis)隶属于颤藻目(Oscillatoriales),Sirenicapillariaceae,是一种能形成水华的浮游性丝状蓝藻,于2013年由KOMREK等人从鞘丝藻属(Lyngbya)分离出来而成立为新属,模式种为希罗泽鞘丝藻(Limnoraphishieronymusii)[10].该属在形态学上与鞘丝藻类(Lyngbya-like)蓝藻相似,均为丝状,不分枝;藻丝单列,具鞘,横壁处不收缢或稍收缢,细胞圆盘状;无异细胞和厚壁孢子.其中,泽鞘丝藻属的种类因能够在细胞中形成气囊从而在水体中自由漂浮,这是区分泽鞘丝藻属和不含气囊而非浮游性的鞘丝藻属(Lyngbya)最主要的形态学特征.目前泽鞘丝藻属中的4个物种均为淡水种类,在生态学上也能与海洋习性的鞘丝藻属区分开来.此外,二者的系统发育关系也相距较远,拥有独立的系统发育位置,彼此之间的16S rRNA基因相似度也低于细菌学和现代蓝藻分类学中属的界限[11-13].因此,KOMREK等将包含模式种在内的4个原来归属于鞘丝藻属的种类(LyngbyahieronymusiiLemmermann 1905、LyngbyabirgeiG.M.Smith 1916、LyngbyacryptovaginataSchkorbatov 1923以及Lyngbyahieronymusiif.robustaParukutty 1940)从鞘丝藻属中分离并成立了新型浮游性蓝藻属——泽鞘丝藻属.
在本研究中,从中国云南省大理市的洱海北部湖湾采集并分离出一株浮游丝状蓝藻藻株,在形态学上与鞘丝藻属相似,基于16S rRNA基因的系统发育分析发现分离的藻株与泽鞘丝藻属的藻种聚为紧密的一个系统进化分支,并显示出最高的序列相似度(>99.84%).进一步通过形态学观察和比较,确定该藻株为泽鞘丝藻属的模式物种希罗泽鞘丝藻.这也是我国对泽鞘丝藻属的新记录属的报道.
1 材料与方法
1.1 样品的采集、藻种分离及培养
2007年7月,使用25#浮游生物网在云南省大理市洱海北部湖湾表层水体的0.5 m处呈“∞”字形缓慢拖动采集浮游植物.首先对聚集的群落进行简单解聚和分散,待群落里面的藻体或者藻丝分散开来以后使用毛细管分离藻种,即在40倍的倒置显微镜(Olympus CKX31,Japan)下用巴斯德吸管制作成的毛细管挑取单根藻丝或单个细胞,在干净的无菌培养基中清洗数次后转入含有无菌CT培养基的24孔板中培养.一定时间后,将镜检为单种蓝藻的孔中的培养物转移到含有10 mL无菌CT培养基的螺口管中,放置在培养条件为温度25 ℃,光暗周期12 L:12 D,光照强度2 000 lx的培养箱中保存,每隔1~2月进行藻种转接.
1.2 藻株的形态观察及形态特征计量
光学显微镜观察:藻株生长至一定生物量时,无菌操作取1 mL藻液,适当稀释后用移液枪吸取适量藻液置于载玻片上,于Nikon eclipse 80i(Nikon,Japan)光学显微镜下进行形态观察,显微镜配备DS-Ri1用于显微照片的拍摄,NIS-Elements 3.2 D用于形态特征的测量(Nikon,Japan).对不同藻株的每个形态特征(如藻丝和营养细胞的长与宽等)均进行100次以上的测量.
1.3 DNA的提取,PCR扩增,克隆及测序
分离得到的藻株采用改良后的十六烷基三甲基溴化铵(CTAB)法提取藻株DNA.将16S rRNA作为 PCR 扩增的目标基因和序列测定,扩增引物为PA(5′-AGAGTTTGATCCTGGCTCAG-3′)[14]和B23S(5′-CTTCGCCTCTGTGTGCCT AGG T-3′)[15].PCR扩增体系为20 μL体系:由1 μL基因组DNA(100 ng/μL),0.5 μL每个引物(10 μmol/L),8 μL无菌水和10 μL 2×PCR TSINGKE Taq聚合酶(Beijing Tsingke Biotech Co.,Ltd.,Beijing,China)组成.扩增程序为:95 ℃预变性3 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸90 s共35个循环;72 ℃延伸5 min.PCR产物通过质量浓度1%琼脂糖凝胶电泳分析,切取的目的条带使用TSINGKE DNA凝胶提取试剂盒(Beijing Tsingke Biotech Co.,Ltd.,Beijing,China)进行纯化回收,然后取5 μL纯化产物在pMD18-T载体(TaKaRa,TaKaRa BioInc.,Otsu,Japan)进行连接,并转化到大肠杆菌Trans5α(TransGen Biotechnology,China)细胞中轻轻混匀,冰浴30 min后42 ℃热击30 s,冰上放置5 min后,在超净台中加入无氨苄青霉素的LB液体培养基200 μL,于37 ℃转速设置为190 r/min的摇床振荡培养45 min后,取70 μL菌液均匀涂布在含Amp+的LB固体培养基上,于37 ℃的恒温培养箱过夜培养12 h.挑取单克隆子菌落加入800 μL LB液体培养,37 ℃,190 r/min振荡培养4~5 h后对菌液进行PCR检测,提取阳性克隆子重组质粒进行Sanger双向测序,测序平台为ABI 3730XL自动测序仪(PerkinElmer,Waltham,Massachusetts USA).每个目标片段至少获得3个阳性克隆序列.
1.4 序列比对分析及系统进化树的构建
测序获得的序列首先在NCBI 网站(http://www.ncbi.nlm.nih.gov/)上进行BLAST比对分析,检测序列是否正确扩增.目的序列与从NCBI网站上下载的相关序列利用BioEdit version 7.2.5.0软件进行比对分析,手工检查并校正,校正后的序列用于系统发育树的构建.系统发育树的构建采用最大似然法(Maximum Likelihood,ML)和贝叶斯推理法(Bayesian Inference,BI),其中最大似然法基于GTR+I+G核酸替代模型,使用PhyML 3.0[16]软件构建ML系统发育树,步展值设为10 000.贝叶斯推理法使用软件MrBayes v3.2.6[17]构建贝叶斯系统进化树,共运行160万代,每100代采样一次,舍弃前25%样本树.软件Fig Tree version 1.4.3(http://tree.bio.ed.ac.uk/software/figtree/)用于系统进化树可视化编辑.
2 结 果
2.1 形态学观察
泽鞘丝藻属(Sirenicapillariaceae,Oscillatoriales)
希罗泽鞘丝藻(图1)
Limnoraphishieronymusii(Lemmermann)J.Komárek,E.Zapomelová,J.Smarda,J.Kopecky,E.Rejmánková,J.Woodhouse,B.A.Neilan &J.Komárková,2013
生境:在淡水水体中自由漂浮.
分布:云南省大理市洱海.
标本号:WZUH-ZLYN200701.
参考藻株:CHAB 3367.

丝状体单生或以小的聚集体生长,不分枝,直或稍弯曲,自由漂浮,具明显鞘.鞘坚固,无色,透明,薄或稍加宽,在末端开放.藻丝单列,圆柱形,蓝绿色到棕绿色,衰老时变黄,在横壁处不收缢或稍收缢,向端部不渐狭.藻丝中部宽12~14 μm,顶端细胞圆形.细胞短柱形或盘状,长1.00~1.84~3.54 μm,宽8.75~10.60~12.90 μm,长短于宽,含颗粒状内容物,具有气囊,细胞壁无色.无异形胞和厚壁孢子.通过藻殖段进行繁殖.
从形态学特征来看,分离的藻株CHAB 3367为丝状,不分枝,具鞘等特点与鞘丝藻相似,但因该种类具有气囊,能在水体中自由漂浮,因此属于从鞘丝藻中分出来的一类具有兼性气囊的泽鞘丝藻类群.从细胞大小来看,CHAB 3367丝状体和藻丝的宽度都比KOMREK等人于2013年描述的希罗泽鞘丝藻略窄,除此之外与该物种具有非常相似的形态学特征,因此,我们从洱海分离的藻株CHAB 3367为中国新记录属泽鞘丝藻属的模式新记录种,即希罗泽鞘丝藻(Limnoraphishieronymusii).
2.2 分子系统学分析
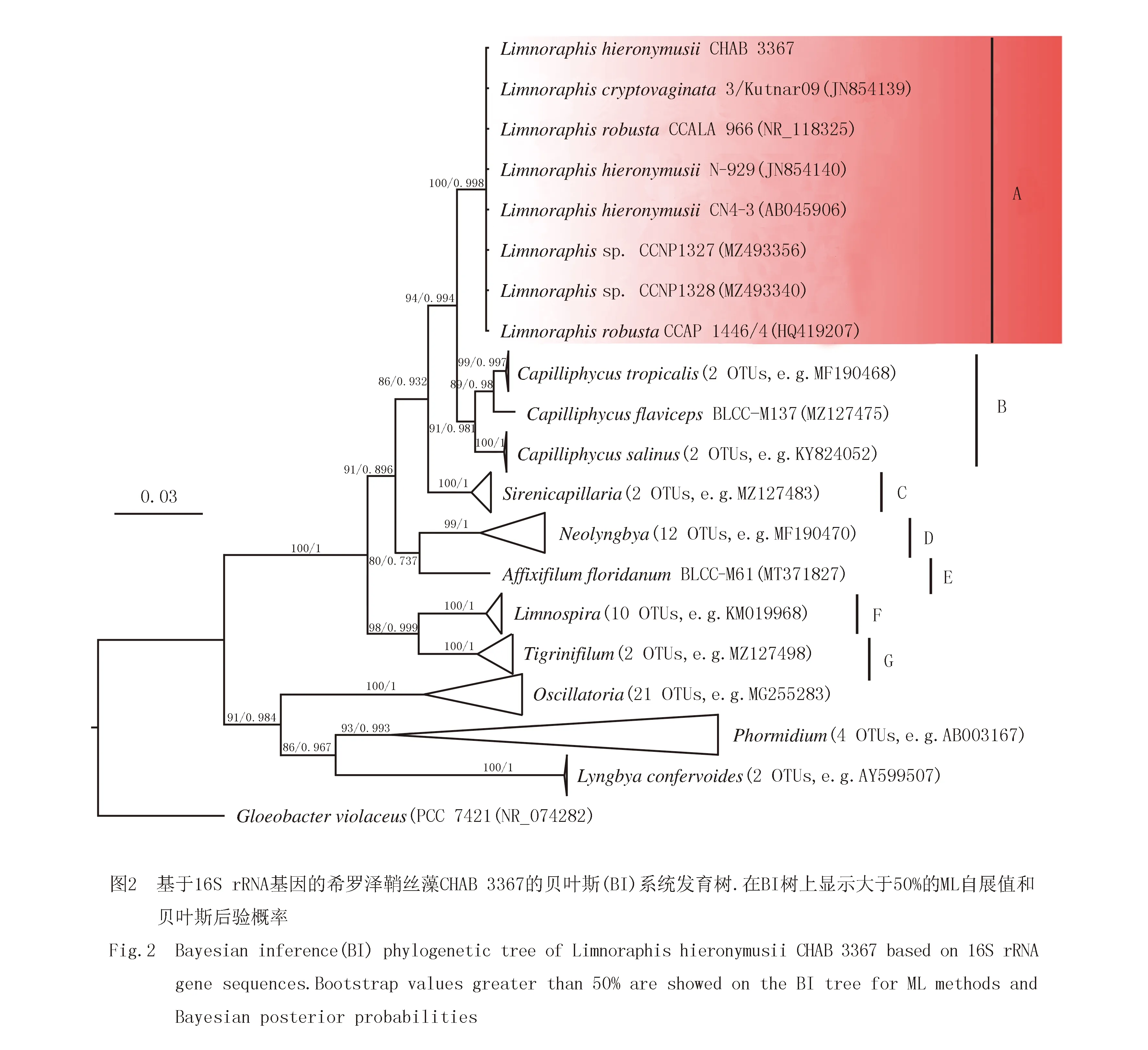
通过一代测序我们获得了CHAB 3367藻株的16S rRNA基因序列(1 463 bp),其与泽鞘丝藻属藻株16S rRNA的序列相似度为99.84%~99.92%(表1),高于细菌学中物种划分的界线.基于ML和BI方法,本研究以黏菌藻目的藻株(GloeobacterviolaceusPCC 7421)为外类群,使用颤藻科和Sirenicapillariaceae下的68条序列来构建系统发育树.从图2可知,CHAB 3367跟泽鞘丝藻属的序列聚为一个紧密的系统进化分支(clade A),ML步展值和贝叶斯后验概率分别为100%和0.998,与Sirenicapillariaceae中其他的6个属分离(clade B~clade G),很大程度上表明分离的藻株CHAB 3367归属于泽鞘丝藻属(图2).
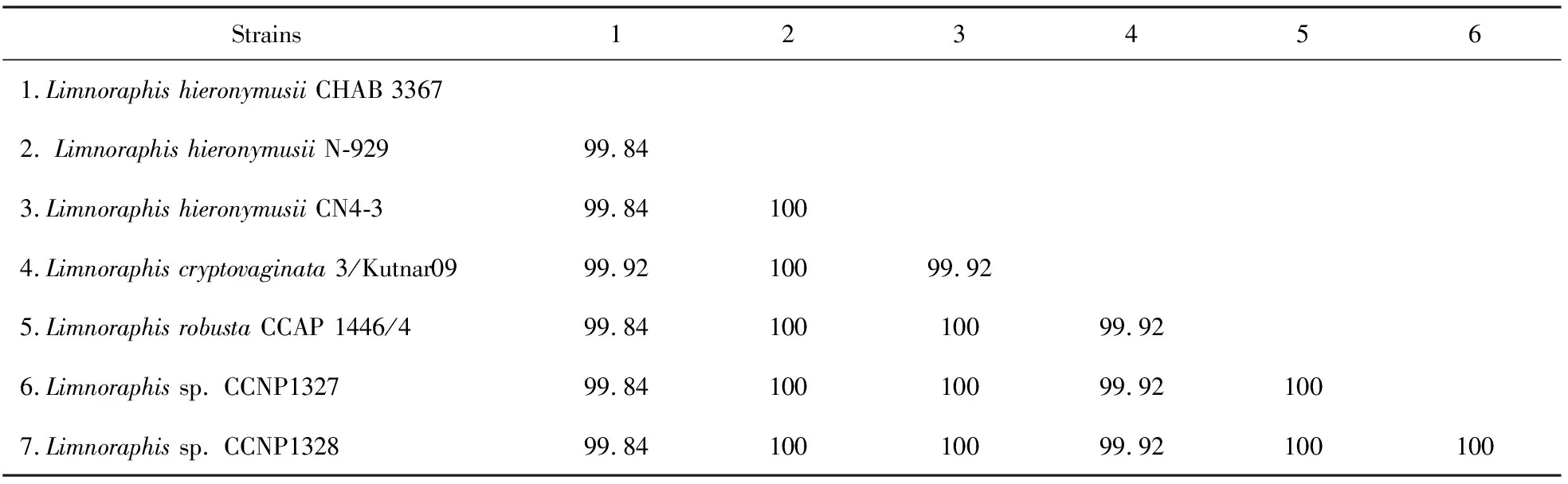
表1 CHAB 3367与泽鞘丝藻属藻株16S rRNA基因序列相似度比较
3 讨 论
随着大量DNA序列的出现和分子生物学的发展,蓝藻的分类学系统经历了几番变化,多相特征方法在蓝藻分类学中的大量应用导致许多新属和新种的出现,并使蓝藻属向狭义单系的“小属”方向推进[5,9,18].在过去几十年里,颤藻目中具有大型盘状细胞的无异形胞分化的丝状蓝藻已被广泛报道[18].其中,鞘丝藻类蓝藻是一类广泛存在于多种生境中的无异形胞分化的丝状蓝藻.根据国际权威藻类数据库Algaebase统计,目前已有952个淡水/海洋鞘丝藻属的种类被记录描述,只有218个物种被现代分类系统有效接受.众多的分子生物学证据表明鞘丝藻属是一个高度多系属,系统发育关系上由几个不同的进化分支组成,一些与模式物种相距较远的类群如Moorea、泽鞘丝藻属(Limnoraphis)、Okeania和微线藻属(Microseira),都是近年来从该属中分离出来而成立的新属[10,20-22].鉴于鞘丝藻属的模式物种为海洋种类,预计这一拥有众多物种的丝状蓝藻属将在未来得到更多的修订和关注.
泽鞘丝藻属因其具有气囊而浮游的特性,区别于传统的附着类的鞘丝藻类,是一类可以形成水华的无异形胞分化的丝状蓝藻[10].泽鞘丝藻属最初被描述为Lyngbyahieronymusii[23],2013年,KOMREK等[10]基于形态学与分子生物学的差异,将具有气囊能漂浮生长的4个鞘丝藻物种Lyngbyahieronymusii、Lyngbyabirgei、Lyngbyacryptovaginata以及Lyngbyahieronymusiif.robusta从鞘丝藻属中分离出来成立为新属,即泽鞘丝藻属,并在次年提出的8目系统中的颤藻目颤藻科中占据一席之地,该属也是当时颤藻科下唯一一个细胞具气囊能形成水华的丝状蓝藻属[5,10].2022年,BERTHOLD等[24]基于16S rRNA基因序列及p-距离的系统发育分析表明,具有高支持度的7个丝状蓝藻属,即Sirenicapillaria、Tigrinifilum、Affixifilum、Capilliphycus、Limnoraphis、Limnospira和Neolyngbya在系统发育树上聚为一个大的紧密的单系系统进化支,因而将该分支重新定义为一个蓝藻新科Sirenicapillariaceae.至此,泽鞘丝藻属的分类地位较为明确地确定下来.
由于泽鞘丝藻属物种间基因型的高度相似性,该属内的种类主要依据形态特征作为进一步的分类依据.当前泽鞘丝藻属中包括4个物种,基于丝状体和藻丝宽度,这4个种类由宽到窄依次为L.birgei、L.robusta、L.hieronymusii及L.cryptovaginata[10].本研究中从洱海分离得到的藻株具有与泽鞘丝藻属相似的形态学特征,其中该藻株的丝状体宽12~14 μm,藻丝宽8.75~10.60~12.90 μm,与KOMREK等[10]2013年描述的L.hieronymusii相比略小一些(丝状体宽12~16 μm;藻丝宽11~14 μm),这可能是由于不同水体分离的藻株具有一定的生态表型差异.由于分离的藻种仍处于该种类的形态特征的大致范围内,我们仍将CHAB 3367定种为泽鞘丝藻属的模式物种,即L.hieronymusiiCHAB 3367.从16S rRNA基因构建的系统发育树来看,CHAB 3367和包括模式种在内的泽鞘丝藻属的序列聚为一个紧密的进化分支(图2 clade A),拥有极高的ML步展值和BI后验概率(100/0.998).根据p-距离计算的16S rRNA基因相似度可知,CHAB 3367与该属所有序列的相似度>99.84%(表1),远高于细菌学与蓝藻分类学中物种的分界线[10-12],进一步证实了形态学的鉴定结果.
此外,泽鞘丝藻属在世界范围分布广泛,目前已知的有欧洲的德国[25]、捷克[26]、乌克兰[27],中东的土耳其[28]、以色列[29],亚洲的萨哈(雅库特)共和国[30]及塔吉克斯坦[31]等.本研究则是泽鞘丝藻属在中国(洱海)的首次报道,也显示出我国无异形胞丝状蓝藻的物种多样性.同时,泽鞘丝藻属作为一种能产生水华的丝状蓝藻,与浮丝藻(Planktothrix)/拟浮丝藻(Planktothricoides)[32],气丝藻(Aerosakkonema)[33]等无异形胞分化的颤藻目蓝藻相似,其形成的蓝藻水华的危害及生理生态学的研究仍需要更多的藻株来证实.未来我们期待更多具有气囊能形成蓝藻水华的无异形胞的丝状蓝藻被发现、分离和鉴定,利用更加先进的宏基因组和高通量测序技术等探索不同气候、不同生境(湖泊、河流、温泉等)下的蓝藻多样性和丰富度信息.
猜你喜欢
中国海洋大学学报(自然科学版)(2023年12期)2023-11-25 13:45:08
生物化工(2022年4期)2022-09-20 09:18:08
法医学杂志(2022年1期)2022-06-21 01:23:34
中国环境科学(2021年8期)2021-09-03 07:14:46
河北渔业(2020年11期)2020-12-21 03:48:44
化学与生物工程(2020年4期)2020-05-08 12:02:10
中国环境科学(2018年7期)2018-07-26 09:03:50
法医学杂志(2016年5期)2016-11-21 01:44:43
中外医疗(2015年5期)2016-01-04 03:57:57
中国环境科学(2015年6期)2015-11-19 08:39:58
