
福建云霄秃房野生茶树群体花器官微形态特征研究
2023-06-30 12:50王攀于文涛蔡春平刘财国王泽涵叶乃兴
江苏农业科学 2023年5期
王攀 于文涛 蔡春平 刘财国 王泽涵 叶乃兴
摘要:为对比福建云霄秃房野生茶群体花部结构的微形态性状差异,利用扫描电子显微镜对云霄秃房野生茶树花器官的细胞形状、表皮纹饰和气孔器起伏程度等性状进行观察,继而进行了主坐标分析和聚类分析。研究表明,云霄野生茶树花器官萼片的内表皮中部及边缘存在平滑型茸毛;花瓣表细胞表面分布着条状、波浪状和脊状等3种纹饰,部分种质存在气孔;花丝表皮细胞排列紧密,有波状、丝状、条状3种表皮纹饰;萼片气孔器大小为195.29~539.52 μm2,开度为0.18~0.40,变异系数为23.16%。通过对10份野生茶树种质不同性状进行PCoA比较,结果显示,采用花器官质量性状即可作为鉴别秃房野生茶种质的依据,也可作为探究云霄秃房野生茶种质亲缘关系的佐证之一。
关键词:野生茶树;云霄;秃房;花器官;微形态;扫描电镜
中图分类号:S571.101 文献标志码:A
文章编号:1002-1302(2023)05-0155-07
茶树[Camellia sinensis (L.) O. Kuntze]是我国南方的重要经济作物之一[1],作为一种多年生常绿木本植物,花器官相较于叶、茎、根等营养器官遗传稳定性更强[2]。同时,花部结构形态性状是研究茶树种质演化的重要内容[3],也是茶树品种鉴定分类的重要特征,因此,对茶花部结构的形态特征研究极为重要,其结果有利于茶种质资源的有效保护和利用。
近年来,扫描电镜在动植物物种的鉴定、种质间的分类和品种保护等领域扮演着重要的角色。而植物花器官微形态的研究一直备受关注,卢广英等通过观察待检植物的横切面、叶表面、花粉形态、动物的体表鳞片、毛发和矿物质的晶体结构等微形态特征,从而为判断中药材的真伪提供了一种可靠的方法[4]。谢微微等对我国10省14份茶树种质花粉的极轴长、赤道轴长和萌发沟长等微形态特征进行描述,丰富了茶树花粉研究内容[5]。杨国一等研究表明不同树型的乌龙茶种质具备不同的叶表纹饰[6]。樊晓静等对11份茶树种质花器官进行了较全面的观测,结果表明其微形态的质量性状比叶片气孔数量性状更可靠,可作为种质鉴别、分类的参考依据[7]。迄今为止,扫描电镜对茶树的应用主要集中在叶片和花粉的观察[8-9]。子房茸毛是茶组植物分类的关键性状[10],王泽涵等报道了福建省内的秃房野生茶树群体,并对其子房茸毛等微形态特征展开描述,扩大了秃房野生茶种质资源的地理区域,丰富了该资源的地域资料[11]。本研究旨在對该秃房野生茶树群体的花柄、花托、花萼和子房等花器官的微形态继续深入展开研究,从而为福建省云霄秃房野生茶树群体的亲缘关系提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为2021年11月采摘的野生茶树花朵,取样地点为福建省漳州市云霄县。样品共计30个,取自小帽山1号、龙翔1号和梁山大茶树1号等10份野生茶树种质(详细信息见表1),每份野生种质资源的3份花器官样品均采自单株。
1.2 试验方法
参考樊晓静等的电镜样品预处理试验方法[7]。
1.3 统计分析
用软件Image J进行数据测量,用Microsoft Excel 2021对数据进行计算与汇总,用软件MVSP v3.13n对数据进行主坐标分析(PCoA)和聚类分析。
气孔数量性状量化方法为测量值计算折合数值,质量性状则按形态特点进行赋值。微形态图选取描述特征具有代表性的茶树种质。
2 结果与分析
2.1 茶树花器官微形态特征
本试验样品为茶花的解剖单位,包括花柄、花托和花萼等7个部位。
2.1.1 茶树花柄、花托微形态性状 扫描电镜图像表明,30个样品中花柄和花托外表面的体表纹饰趋于相同(图1-A1、图1-A2),细胞均呈矩形状,体表纹饰主要表现出细的长矩状纹路,10份茶树种质资源花柄表皮上未见气孔;在小帽山2号、小帽山5号、小帽山7号和梁山大茶树1号的花托上均观察到气孔(图1-A3、图1-A4),而其他茶树种质资源花托上未见气孔。
2.1.2 茶树花萼、花瓣微形态性状 扫描电镜下图像表明(表2、表3、表4),样品茶树花萼的内侧体表细胞主要为不规则的多边形,表面光滑。其中,梁山大茶树1号的萼片内表皮存在较多的绒毛(图 1-B1),而小帽山1号、小帽山2号和小帽山5号的萼片内表皮茸毛数量较少(图1-B2);萼片外表细胞主要呈现不规则状,表面较光滑具平展纹饰、皱脊状纹饰,气孔器主要分布于根部或边缘,气孔均凸起于细胞表面。小帽山3号、龙翔2号的花萼内侧体表细胞则呈现起伏不定(图1-B3),小帽山4号、小帽山6号和梁山大茶树1号的花萼内侧体表细胞较平展且饱满(图1-B4)。小帽山2号、小帽山6号的花萼外侧气孔显著凸出于外侧体表(图1-B5),小帽山7号显著低于表皮(图1-B6),其他种质均稍突出于表皮;小帽山2号、小帽山7号、梁山大茶树1号和龙翔2号的花萼外缘角质层呈脊状增宽;小帽山2号的花萼外侧气孔周边较平滑(图 1-B7),其他种质的花萼外侧气孔周边大多分布皱脊状纹饰(图1-B8)。萼片气孔大小为195.29~539.52 μm2,开度为0.18~0.40。
10份种质的花瓣体细胞外形有不规则的多边形、五边形、六边形、近圆形4种类型(图1-C1、图1-C2、图1-C3、图1-C4),其中分布着波浪状、条纹状、皱脊状等纹饰。小帽山4号、龙翔1号、小帽山7号和梁山大茶树1号的花瓣表细胞形态特征表现出非单一性,如小帽山7号茶树的花瓣表细胞形状有不规则形、五边形等,而龙翔1号茶树的花瓣表细胞形状有不规则形、近圆形等。其他种质形状多为不规则或单一多边形;除小帽山6号、梁山大茶树1号、龙翔2号未见气孔外,其余种质的花瓣体表均观察到气孔存在。小帽山1号、小帽山2号和龙翔1号的花瓣气孔略低于体表(图1-C5),其余种质的花瓣气孔器与表皮持平。小帽山1号、小帽山2号、小帽山4号和小帽山7号的花瓣气孔外缘围绕粗条状纹饰(图1-C6),龙翔1号气孔器外拱盖明显增大且外围覆盖辐射状纹饰(图1-C7),其余种质均为环绕皱脊状纹饰(图1-C8)。
2.1.3 茶树雄蕊微形态性状 扫描电镜下图像表明,花丝表皮细胞紧密排列,纹饰为条状、丝状、波状。小帽山5号、小帽山7号种质存在极少数气孔,其他种质未发现茸毛和气孔。龙翔1号、梁山大茶树1号的体表纹饰表现为水波形(图1-D1);小帽山4号、小帽山5号和龙翔2号表现为盘曲且略粗的丝状(图1-D2);小帽山1号、小帽山2号、小帽山3号、小帽山6号和小帽山7号表现为微曲的条形(图1-D3、图1-D4)。
2.1.4 茶树雌蕊微形态性状 运用扫描电镜观察样品花柱,其表细胞突起,花粉粒附着可见,这表明柱头上正进行授粉过程。花柱呈上下底面积差异较小的圆台形(图1-E1),从花柱根部至顶端其横截面渐小,体表细胞紧凑。根据花柱细胞形状可把10份茶树种质分为梭形、长条纹形和不规则的多边形3种类型,其中,小帽山2号、小帽山3号为梭形(图1-E2),小帽山1号、小帽山4号、龙翔1号、小帽山7号和梁山大茶树1号为长条纹形(例图1-E3),小帽山5号、小帽山6号和龙翔2号则为不规则的多边形(图1-E4)。茶树子房根据是否被茸毛可分为2类:小帽山2号和小帽山3号种质的子房
[FK(W41][TPWP11.tif][FK)]
满被茸毛(图1-F1),茸毛纹饰为平滑型。小帽山1号、小帽山4号、小帽山5号、小帽山6号、龙翔1号、小帽山7号、梁山大茶树1号和龙翔2号种质资的子房未被茸毛(图1-F2)。小帽山5号、梁山大茶树1号、龙翔1号和小帽山6号表皮细胞凸起(图1-F3),小帽山1号和小帽山7号子房体表细胞略微内陷(图1-F4),小帽山2号、小帽山3号、小帽山4号和龙翔2号的体表细胞相对平缓。
2.2 茶树花器官气孔数量性状的变异分析
通过对样品所观测到的花萼气孔数量性状进行变异分析,其变异系数结果在2.04%~53.68%之间,平均值为23.16%。种质内气孔数量性状的变异系数平均值为10.87%~29.36%,而种质间气孔数量性状变异系数的平均值为14.78%~32.99%。可见,茶花部的结构气孔数量特征不稳定,不具备明显的可区分性。
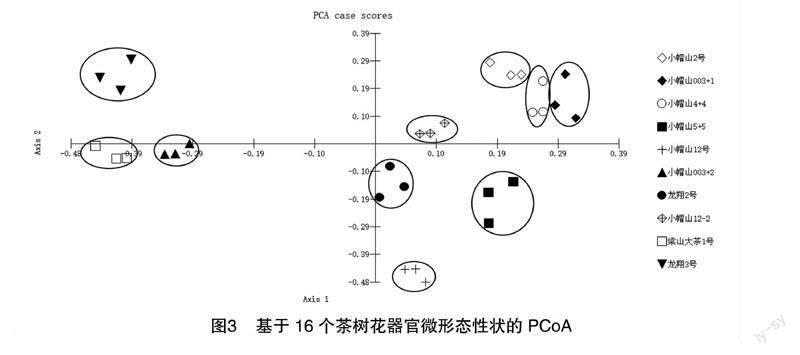
2.3 基于茶树花器官微形态性状的主坐标分析
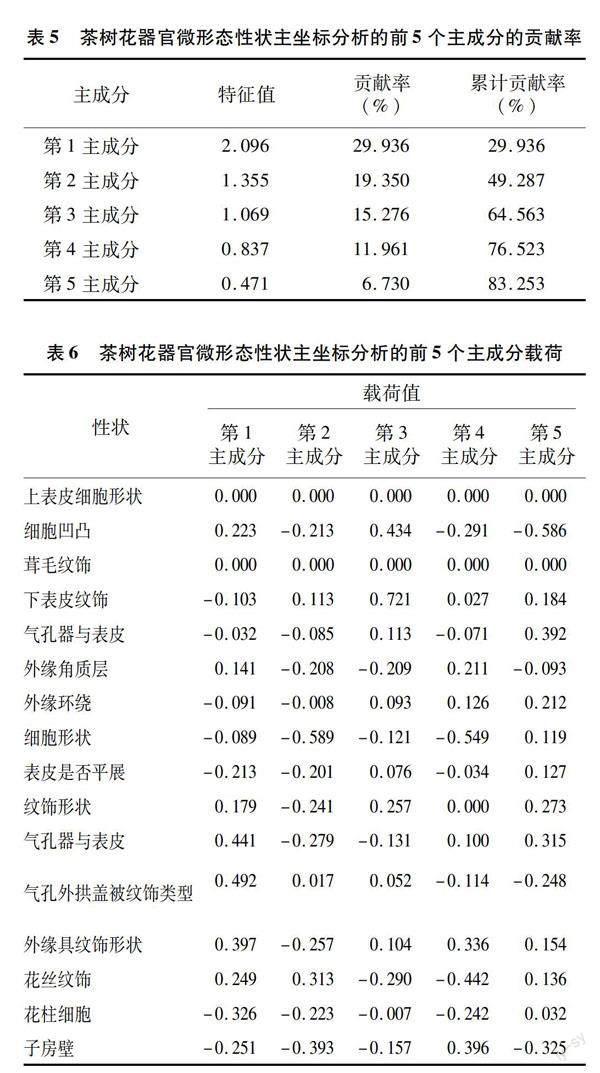
对样品的22个茶花部结构微形态特征(包括6个气孔数量特征在内)进行PCoA,结果表明(图2),试验材料中,梁山大茶树1号、小帽山2号、小帽山6号、龙翔1号和小帽山7号等种质之间存在相互交错现象, 未见明显区分种质。利用样品的质量性状数据展开主坐标分析,结果见图3,30份样品聚成了10个独立的区域,且每个区域各为一种种质。分析结果(表5、表6)的前5个主成分的累计贡献率总和高达83.253%,这意味着这5个主成分可以作为种质区分的关键因素。其中,第1主成分的贡献率为29.936%,主要由花瓣气孔外拱盖所披纹饰类型、花瓣气孔器与表皮的关系所决定;第2主成分的贡献率为19.350%,主要由花瓣细胞形状、子房壁形状决定;第3主成分的贡献率为15.276%,主要由花萼外表纹饰形状、花萼内表细胞决定;第4主成分的贡献率为11.961%,主要由花瓣细胞的外形特征、花丝表面的纹饰决定;第5主成分的贡献率为6.730%,主要由萼片内表细胞起伏、花瓣气孔与体表细胞决定。对22个微形态特征(含6个气孔数量特征)和16个微形态特征(仅含质量特征)的分析显现出,气孔器数量性状的存在将导致聚类效果不佳;而只用质量性状时,种质聚类效果得到显著的改善。
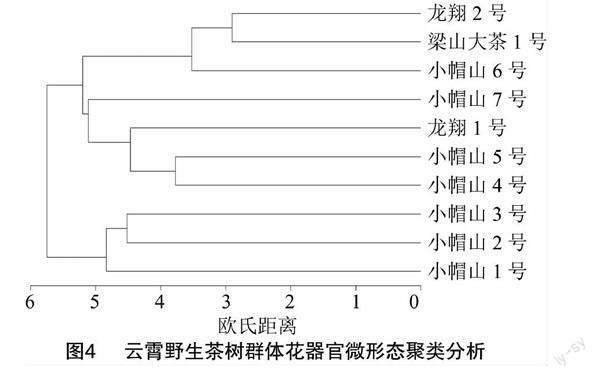
2.4 基于茶树花器官微形态性状的聚类分析
由于气孔数量性状不具备品种特异性,根据10份供试材料的数据,对16个质量性状进行聚类分析,如图4所示。在欧式距离为5.75时,10份种质可分为2支,小帽山1号、小帽山2号和小帽山3号聚为一支, 这3份种质的共同特征是萼片内表细胞不规则,萼片外表细胞形状不规则,气孔器稍凸出表皮;花瓣细胞形状为五边形、不规则多边形,存在明显的皱脊纹饰,小帽山1号、小帽山2号的花瓣气孔器凹陷于表皮,外缘脊状具明显条状纹饰环绕,而小帽山3号的花瓣气孔则持平于表皮,外缘脊状具皱脊状环绕。龙翔1号、梁山大茶树1号和小帽山4号等7份种质聚为另一支,它们的共同特征:茸毛纹饰为平滑型,萼片内表皮形状呈现不规则,萼片外表光滑;花瓣细胞形状为多边形,气孔外拱盖平滑。
3 讨论与结论
3.1 讨论
3.1.1 茶树花器官微形态的质量性状与数量性状 长期以来,植物的形态性状在探究种间或属间分类等方面发挥着不可代替的作用,具有重要的研究意义[12-14]。通过对试验数据整理分析,结果表明不同部位的形态性状往往存在可区别性,如花瓣与萼片在细胞形状、纹饰等方面均存在明顯差异。不同种质的同一部位往往有不同的形态特征,如萼片的气孔器与表皮有凹陷、凸出2种;子房存在被茸毛、未被茸毛2种。植物表皮毛的纹饰特征是研究植物种间或属间关系,从而进行区别分类的重要途径之一[15-16]。此外,本研究观测到花托、萼片、子房的茸毛纹饰均为平滑型,这与樊晓静等的研究结果[7]一致,同时,也表明茶花茸毛纹饰不因部位差异而具备特异性。气孔作为植物进行物质循环和能量供应的场所,在植物生命活动中发挥重要作用,它存在于植物的大多数部位[17]。在茶树中,目前已有众多学者针对叶片气孔进行了各种相关研究,其中包括茶叶加工过程中的气孔变化[9]、栽培环境对气孔的影响等[18]。此外,样品中的萼片上存在较多气孔,花瓣、花丝、花托等样本存在少许气孔。生物性状通常受到基因和环境协同调控,如气孔的类型与物种的遗传物质密切相关,而气孔器的数量特征受众多生态因子调节[19]。同时,本研究对30份样品所观察到的花萼气孔数量性状进行了变异系数分析,结果显示气孔数量性状在种质间或种质内都具有高度的变异,说明气孔数量性状受环境影响较大,不适合作为种质鉴定、种质区分的依据。对22个微形态特征(含6个气孔数量特征)和16个微形态特征(仅含质量特征)进行主PCoA,结果与上述观点一致,茶树花器官的花瓣气孔外缘纹饰、花瓣气孔器与表皮、子房壁形状、花瓣细胞形状、花萼外表皮纹饰形状、花萼内表细胞起伏等质量性状往往在同一种质上能够较稳定的存在,对茶树的品种溯源、亲缘关系和开发应用等研究有积极意义。
3.1.2 基于花器官微形态的秃房野生茶树群体亲缘关系分析 20世纪中叶以来,福建茶叶科技人员开展了野生茶种质调研与考察,在宁德市等[20-22]多地发现野生茶树资源,此后,陈潇敏等陆续又在福建多地发现苦茶野生资源,而秃房茶树(子房无茸毛)仅发现个别单株[23-25]。据已有报道,茶组植物秃房茶主要分布在滇、黔、粤、桂、川。此外,广东凤凰水仙群体后代中也曾发现秃房单株[26]。本试验的10份云霄秃房野生茶群体种质的30份样本在第1和第2主坐标排序中分为3个群体,第1群体包括小帽山1号、小帽山2号和小帽山3号;第2群体包括采自梁山的龙翔1号、小帽山4号、小帽山5号和小帽山7号;第3群体包括采自小帽山的小帽山6号、梁山大茶树1号和龙翔2号。由图3和图4可知,聚类分析和主坐标分析都将10份种质划分成3个群体,但2种方法在种质间亲缘关系上的研究存在差异。主坐标分析显示,第1群体和第2群体亲缘关系更近,而聚类分析第2群体和第3群体亲缘关系更近,这说明第2群体亲缘关系更复杂,该结果与SNP分子标记技术研究的该地区种质亲缘关系可相互印证[27]。
3.2 结论
通过比较30个云霄秃房野生茶树种质资源花器官样品的多个部位微形态的差异,筛选出茶树花器官的花瓣气孔外拱盖所披纹饰类型、花瓣气孔器与表皮、子房壁形状、花瓣细胞形状、花萼外表皮纹饰形状、花萼内表皮细胞起伏等质量性状即可作为秃房野生茶树鉴别的依据,也可作为探究云霄秃房野生茶种质亲缘关系的佐证之一。
致谢:本研究试验样品采集承蒙福建省云霄县茶叶科学研究所王彩云、方德音、蔡捷英、王金焕等老师的大力支持,特此致谢!
参考文献:
[1]叶乃兴. 茶学概论[M]. 2版. 北京:中国农业出版社,2021.
[2]Caseys C. Shy girl gives kiwifruit male flowers [J]. The Plant Cell,2018,30(4):739-740.
[3]陈常颂,余文权. 福建省茶树品种图志[M]. 北京:中国农业科学技术出版社,2016.
[4]卢广英,刘红燕,邢训颜,等. 扫描电镜在中药真伪鉴别中的应用[J]. 电子显微学报,2021,40(4):483-487.
[5]谢微微,于文涛,杨国一,等. 14个茶树品种的花粉微形态观察[J]. 南方农业学报,2018,49(9):1698-1704.
[6]杨国一,于文涛,郑 晶,等. 乌龙茶种质叶片微形态特征的扫描电镜观察[J]. 南方农业学报,2018,49(10):2020-2027.
[7]樊晓静,于文涛,蔡春平,等. 茶树种质资源花器官微形态特征观察[J]. 南方农业学报,2021,52(3):700-710.
[8]樊晓静,于文涛,刘登勇,等. 福建寿宁野生茶树种质资源叶片和花粉微形态观察[J]. 福建农业学报,2019,34(3):298-305.
[9]刘财国,于文涛,樊晓静,等. 白茶萎凋过程叶片微形态动态变化规律[J]. 茶叶学报,2021,62(2):73-77.
[10]张宏达. 茶树的系统分类[J]. 中山大学学报(自然科学版),1981,20(1):89-101.
[11]王泽涵,于文涛,樊晓静,等. 福建秃房野生茶种质资源新纪录及其子房微形态观察[J]. 福建农业学报,2020,35(8):830-836.
[12]林新春,俞志雄. 木兰科植物的叶表皮特征及其分类学意义[J]. 浙江林学院学报,2004,21(1):33-39.
[13]Song J H,Yang S,Choi G. Taxonomic implications of leaf micromorphology using microscopic analysis:a tool for identification and authentication of korean piperales[J]. Plants,2020,9(5):566.
[14]Bhat M A,Shakoor S A,Badgal P,et al. Taxonomic demarcation of Setaria pumila (Poir.) Roem. & Schult.,S. verticillata (L.) P. Beauv.,and S. viridis (L.) P. Beauv. (Cenchrinae,Paniceae,Panicoideae,Poaceae) from phytolith signatures[J]. Frontiers in plant science,2018,6(9):864.
[15]王 虹,王 磊,范林仙,等. 新疆14种青兰属植物叶表皮微形态结构研究[J]. 西北植物学报,2014,34(10):2004-2019.
[16]李 烨,李秉滔. 蜡梅科植物的叶表皮特征及其在分类上的意义[J]. 热带亚热带植物学报,1999,7(3):202-206.
[17]杨 洋,马三梅,王永飞. 植物气孔的类型、分布特点和发育[J]. 生命科学研究,2011,15(6):550-555.
[18]唐荣南,汤兴陆. 松茶间作茶树叶片的解剖构造和气孔活动[J]. 应用生态学报,1991,2(3):201-206.
[19]刘 婧,王宝山,谢先芝. 植物气孔发育及其调控研究[J]. 遗传,2011,33(2):131-137.
[20]郭元超. 武夷类群的地理分布与演化特征[J]. 茶叶科学简报,1994(3):4-10.
[21]詹梓金,林茂锋. 福建野生茶资源的地理分布与生态型[J]. 福建农学院学报,1991,20(4):427-433.
[22]楊如兴,陈芝芝,张 磊,等. 福建野生茶树种质资源保护与利用[J]. 茶叶学报,2017,58(3):96-101.
[23]陈潇敏,王鹏杰,王淑燕,等. 基于转录组挖掘蕉城苦茶苦茶碱合成相关基因[J]. 应用与环境生物学报,2021,27(5):1382-1389.
[24]陈潇敏,赵 峰,王淑燕,等. 福建野生茶树资源嘌呤生物碱构成评价及特异资源筛选[J]. 茶叶科学,2022,42(1):18-28.
[25]陈潇敏,赵 峰,金 珊,等. 福建云霄地方茶树品种资源生化成分特征分析与评价[J]. 西北植物学报,2022,42(1):127-137.
[26]周梦珍,谢曼卫,陈坚升,等. 抗寒凤凰水仙茶树资源性状调查[J]. 中国茶叶,2019,41(6):47-51.
[27]王泽涵,于文涛,方德音,等. 基于EST-SNP的福建云霄茶树种质资源遗传多样性分析[J]. 福建农业学报,2021,36(12):1431-1438.
收稿日期:2022-04-25
基金项目:福建省科技计划引导性项目(编号:2021N0024);福建农林大学科技创新专项(编号:CXZX2020123B);海关总署科研项目(编号:2020HK187);福建张天福茶叶发展基金会科技创新基金(编号:FJZTF01)。
作者简介:王 攀(1996—),男,四川巴中人,硕士研究生,研究方向为茶树栽培育种与茶叶品质。E-mail:1048773934@qq.com。
通信作者:叶乃兴,硕士,教授,主要从事茶树栽培育种与品质化学研究,E-mail:ynxtea@126.com;于文涛,博士,高级农艺师,主要从事植物种质资源及外来入侵植物研究,E-mail:wtyu@foxmail.com。
猜你喜欢
收藏与投资(2023年12期)2024-01-03
中国棉花(2023年9期)2023-12-17
当代作家(2023年3期)2023-04-23
少年漫画(艺术创想)(2019年10期)2019-04-20
智富时代(2018年7期)2018-09-03
智富时代(2018年7期)2018-09-03
军营文化天地(2017年7期)2017-09-25
电线电缆(2017年4期)2017-07-25
岩矿测试(2015年3期)2015-12-21
扬子江诗刊(2015年4期)2015-11-14
