
八角林不同郁闭度对鳞尾木生长发育及光合特性的影响
2023-06-29 09:55:20魏秋兰杨卓颖杨日升梁文汇安家成朱昌叁
南方农业 2023年7期
魏秋兰,杨卓颖,杨日升,梁文汇,安家成,朱昌叁
(广西林业科学研究院/广西特色经济林培育与利用重点实验室,广西南宁 530002)
鳞尾木[Champereia manillanavar.longistaminea]是山柚子科茎花山柚属多年生常绿小乔木,又名茎花山柚[1],别名龙须菜(广西田东)、甜菜树(云南富宁)、雷公菜(广西田林)、侧本或满菜(广西靖西、德保),常与假苹婆(Sterculia lanceolata)、苦槠栲(Castanopsis sclerophylla)等伴生[2]。鳞尾木幼嫩茎、芽叶具有丰富的营养成分,是一种极具特色的森林蔬菜[3-4],因其含有艾杜醇、多糖、黄酮及多种药效氨基酸而具有一定的药用开发价值[5-7]。对鳞尾木的研究涉及分类[8]、资源调查、育苗[9]及仿生栽培[10]等,但林下套种时郁闭度大小对鳞尾木生长的影响未见报道。广西是八角(Illicium verumHook.f.)原产地,有“世界八角之乡”的美称[11],栽培面积广大,在八角林下套种森林蔬菜等其他作物,能提高林地的利用率,还可改善林中的光温和土壤结构,提高林业的综合经济效益。光是植物生长的必需环境因子之一,不但对植物的生长发育和生理变化起着重要的作用,也是植物进行光合作用的重要能源和基础[12]。不同郁闭度下光照、水分和湿度等因素差异必然影响林下套种作物生长发育,探讨林分不同郁闭度对鳞尾木生长及生理指标的影响,以期为八角林下套种鳞尾木提供理论依据。
1 材料与方法
1.1 试验材料
供试鳞尾木种子采自广西田林,2018 年6 月采种育苗,2019 年3 月16 日选用生长健壮、长势一致的容器苗出圃造林。
1.2 试验方法
1.2.1 试验地点
试验地点位于广西林科院老虎岭试验林场,海拔为90~130 m,属南亚热带季风气候,年均温21.7 ℃,年平均降水量1 347.2 mm,干湿季节明显,土壤大部分是由砂页岩发育而成的红壤,pH 值5~6,肥力均匀。试验地八角林龄13年,种植密度为2.5 m×2.5 m,因种源与管理措施差异等原因形成不同郁闭度。
1.2.2 试验方法
八角林分的郁闭度通过目测法结合CI-110植物冠层分析仪进行测量[13],并每隔3 个月进行1 次抚育修枝,调控试验区域的郁闭度。鳞尾木栽培试验为完全随机区组设计,林分郁闭度设置4 个处理:全光照(CK)、郁闭度0.3(B1)、郁闭度0.5(B2)、郁闭度0.8(B3),每个处理3次重复,小区样地面积为5.0 m×5.0 m,造林株行距为1.0 m×1.0 m,即每个样地种植25株。
1.3 测定指标及方法
鳞尾木栽培1 年后,于2020 年3 月进行各项生长及生理指标的测定。
1.3.1 株高、地径测定及保存率的统计
以直尺测量株高,游标卡尺测量地径。在试验开始及结束时测定株高和地径,计算各处理株高或地径的增量。调查鳞尾木苗木成活及死亡株数。计算保存率,计算公式如下:
保存率=成活株数/总株数×100%
1.3.2 光合参数的测定
2020 年3 月25 日晴天上午9:00—11:00 进行光合参数的测定。使用Li-6400 XT 便携式光合仪(LICOR,USA),红蓝光源、光强设置为1200 mol·m-1·s-1,随机选取各处理3 株苗木,每株苗木选取中上部3 片叶片连续测定5 次,取平均值。测定同一叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)等参数。计算水分利用效率(WUE),计算公式如下:
1.3.3 叶形态指标的测定
在不同郁闭度八角林下每个小区内随机选择5 株苗木,从不同方位摘取成熟健康叶30片迅速带回室内测定。采用LI-3000C便携式叶面积仪测定叶长、叶宽和叶面积。采用电子天平测定叶片鲜重、干重,直接称量鲜叶测定鲜叶重,鲜叶经恒温干燥(115 ℃杀青30 min,75 ℃恒温下烘干至恒重)后称干叶重。计算叶片含水率与比叶面积公式如下:
叶片含水率=(鲜叶重量-干叶重量)/鲜叶重量×100%
比叶面积(SLA)=叶面积/叶片干重
1.3.4 光合色素含量的测定
采用浸提法测定光合色素含量[14]。各处理中随机采集健康成熟叶片,洗净吸干并除去中脉后剪碎,以95%乙醇和无水丙酮按1∶1比例制备浸提液,黑暗浸提24 h 直至绿色消去,用紫外可见分光光度计分别在663、645、470 nm 波段测量吸光值,计算3 种光合色素含量。
1.4 数据分析
利用Microsoft Office Excel 2010和SPSS20.0进行数据统计与分析,采用单因素方差分析(one-way ANOVA)及Duncan 法进行多重比较(α=0.05)。采用隶属函数分析法对各处理指标进行综合评价。隶属函数的计算公式如下[15]:
式中:Xi为指标测定值,Xmax为某处理指标最大值,Xmin为某处理指标最小值。
2 结果与分析
2.1 不同郁闭度对鳞尾木生长量及保存率的影响
不同郁闭度对鳞尾木生长量及保存率的影响结果见表1。可以看出,不同郁闭度对鳞尾木株高生长量、地径生长量和保存率有显著影响(p<0.05)。植株保存率、株高生长量和地径生长量均随着郁闭度增加先上升后下降。不同郁闭度处理下,保存率、株高生长量、地径生长量均大于CK 处理。其中B2 处理下保存率为96.37%,株高生长量为21.42 cm·a-1、地径生长量为0.48 cm·a-1,均为各处理中的最大,分别是CK的1.20、1.58、1.66倍。
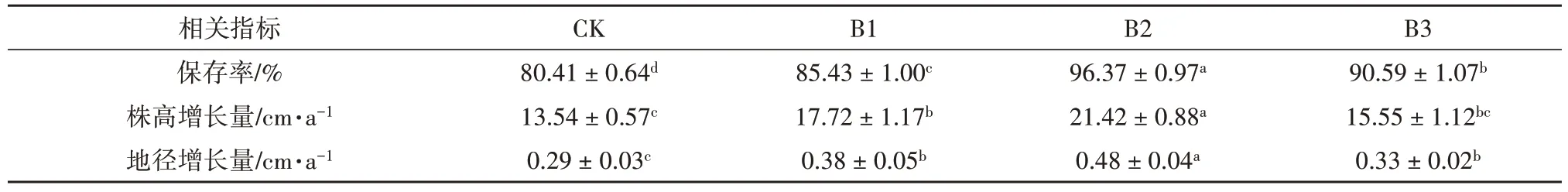
表1 不同郁闭度对鳞尾木生长的影响
2.2 不同郁闭度对鳞尾木叶片形态的影响
不同郁闭度对鳞尾木叶片形态的影响结果见表2。从表2 可知,郁闭度对鳞尾木叶片的LS、LL、LW、LB、CLWC、SLAM均有显著性影响(p<0.05)。随着郁闭度的增加,叶面积、叶长、叶宽均先增加后减少,在B2 处理下最大,分别为341.02 cm2、132.61 mm、38.45 mm,比CK 高39.93%、12.86%和9.17%;长宽比整体呈先下降后上升的趋势,B2 处理下最小为3.45,B3 处理下最大为3.96;叶片含水量、比叶面积均随着郁闭度的增加呈逐渐增加的趋势,在B3处理下最大,分别为67.88%、127.29 cm2.g-1,比CK高9.22%、102.63%。
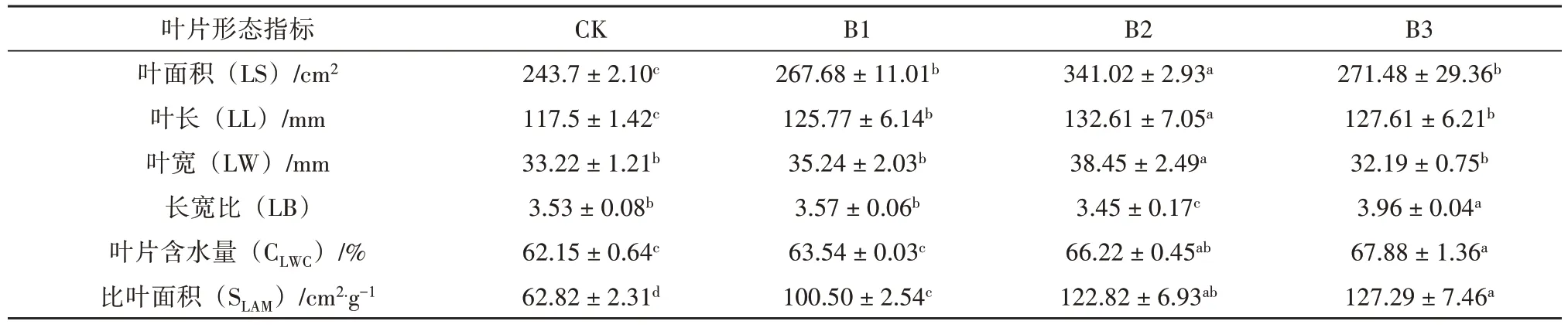
表2 不同郁闭度对鳞尾木叶片形态指标的影响
2.3 不同郁闭度对鳞尾木光合特性的影响
不同郁闭度对鳞尾木光合特性的影响结果见表3。由表3 可知,不同郁闭度对鳞尾木的净光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE)、胞间二氧化碳浓度(Ci)、气孔导度(Gs)均有显著影响(p<0.05)。Pn、Tr、WUE、Gs 随着郁闭度增加先上升后下降,以上指标B2 处理均为最大,分别为3.87 μmol·m-2.s-1、2.63 μmol·m-2.s-1、2.82 μmol·mol-1、0.13 μmol·m-2·s-1,是CK 的4.72、1.65、1.94、2.17倍;胞间二氧化碳浓度(Ci)随着郁闭度的增加先下降后上升,B2 处理最小(305.75 μmol·mol-1),B3 处理下最大(381.33 μmol·mol-1),是CK 处理的1.13倍,各处理间差异显著。当郁闭度低于0.5 时,随着郁闭度的增加,除胞间二氧化碳浓度以外,净光合速率、蒸腾速率、水分利用效率均随着叶片气孔导度上升而上升。
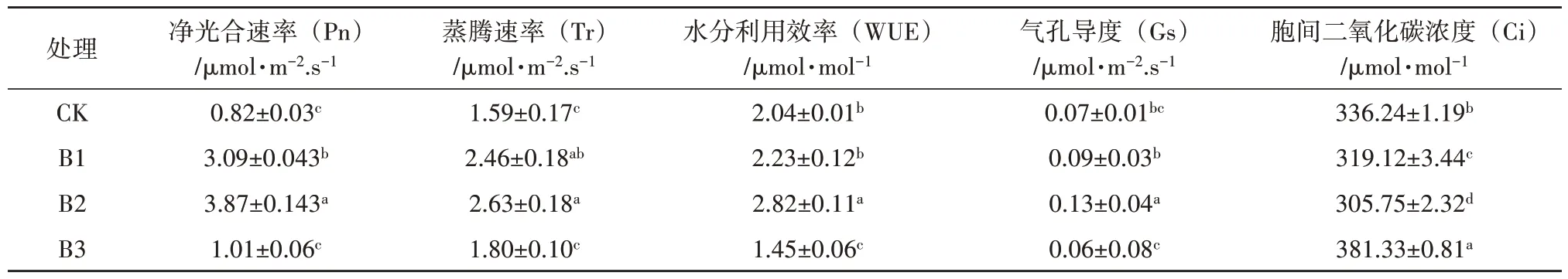
表3 不同郁闭度对鳞尾木光合参数的影响
2.4 不同郁闭度对鳞尾木光合色素含量的影响
不同郁闭度对鳞尾木光合色素含量的影响结果见表4。由表4可知,不同郁闭度对鳞尾木的光合色素含量均有显著影响(p<0.05)。随着郁闭度的增大,叶片叶绿素含量逐渐增加,叶绿素总量(Chl a+b)、叶绿素a(Chl a)、叶绿素b(Chl b)、类胡萝卜素(Caro)的含量均在B3 处理时达到最大值。叶绿素a/b(Chla/b)值则随着郁闭度的增大逐渐减小,CK处理最大。
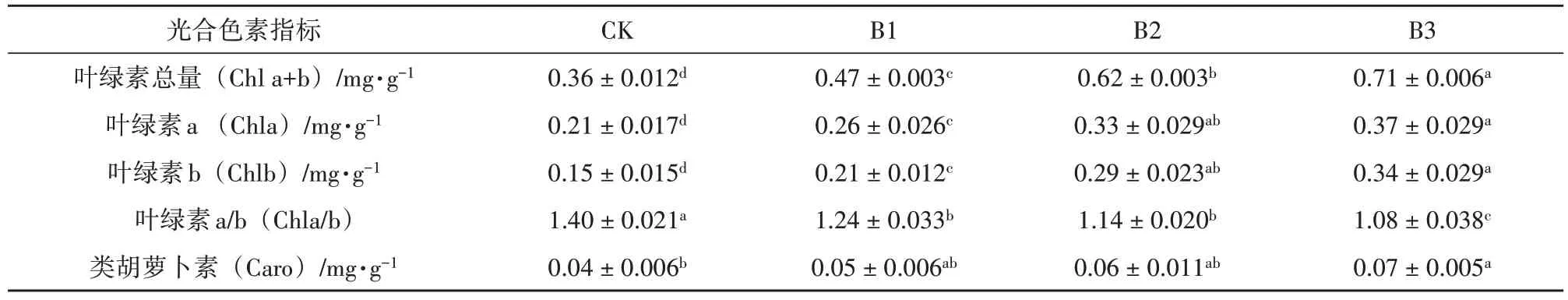
表4 不同郁闭度对鳞尾木光合色素含量的影响
2.5 隶属函数分析
对鳞尾木各项生长和生理指标进行隶属函数分析,可以综合评价不同郁闭度处理下苗木生长状况,分析结果见表5。各处理组的隶属函数均值从大到小依次为:B2>B3>B1>CK。林下各处理组的隶属函数值均大于CK。
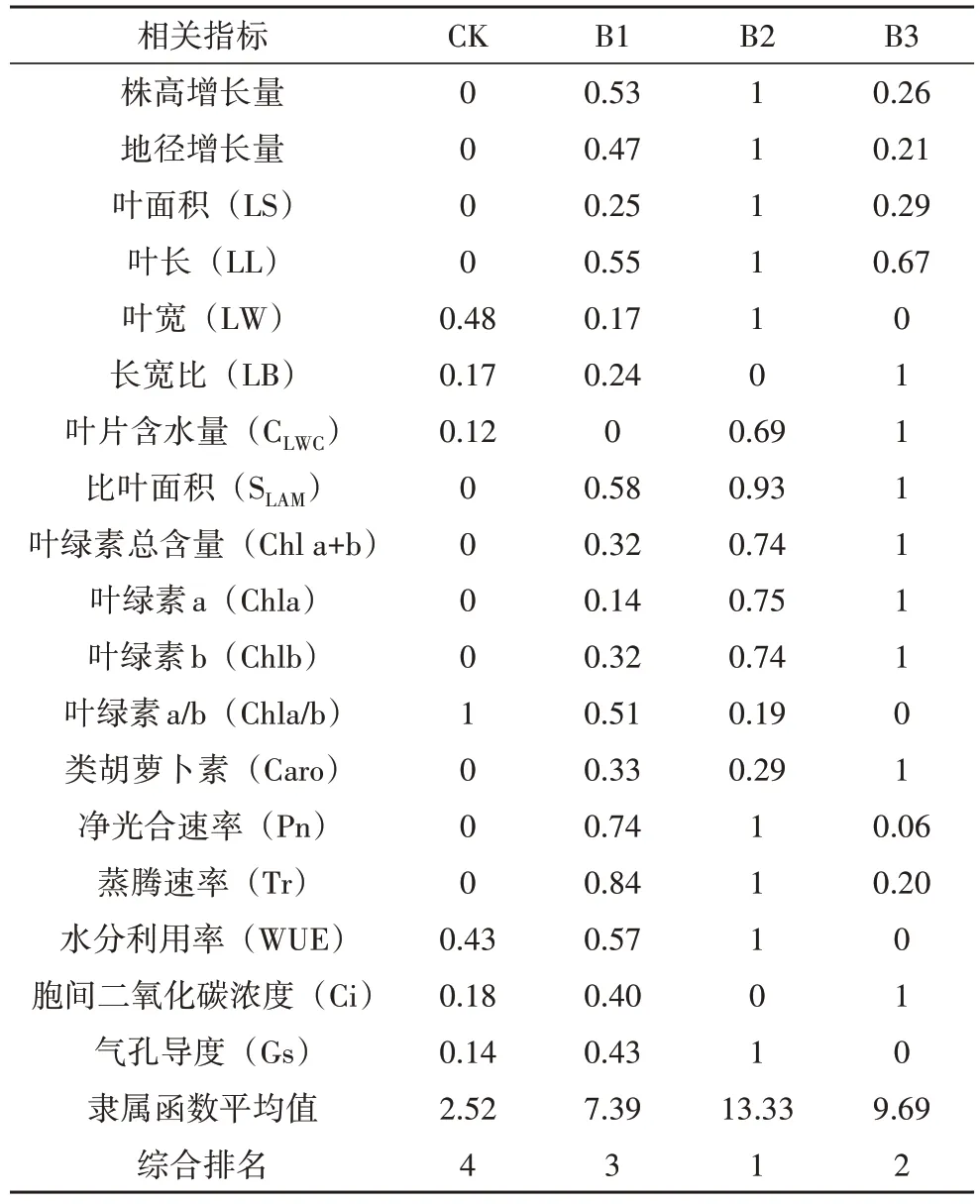
表5 不同郁闭度对鳞尾木生长的隶属函数综合评价
3 小结与讨论
光照是影响植物生长和发育的重要因子之一,植物受到强光胁迫时光合能力下降,有机物积累减少,生长不良甚至死亡[16]。株高和地径是反映植物是否适应环境的直观表现[17]。叶片是对环境变化最为敏感且可塑性较强的器官[18],其形态多变且直接影响植物生理生化过程。叶片含水量和比叶面积则随着郁闭度增大而增大,说明鳞尾木在弱光下通过增大比叶面积提高光合能力,以补偿光合有效辐射降低造成的损失[19]。气孔是外界与植物细胞内进行气体交换的窗口,直接影响到植物光合作用进程[20]。张建新等对马蹄莲的研究也发现这一现象[21]。胞间二氧化碳浓度随着郁闭度增大先下降后上升,B3处理下数值最大,B2处理下的最小。如果Pn 降低伴随着胞间CO2浓度升高,光合作用限制因素则是光合机构活性受到严重破坏,植物生长受阻[22]。光合色素各组分的含量及Chl a与Chl b 的比值一定程度上反映了植物是否适应所处的光照环境[23]。郁闭度变大导致光合色素含量增加,在三叶青(Tetrastigma hemsleyanum)栽培研究中也有发现[24],而叶绿素a/b 值因郁闭度变大而减少,在刻叶紫堇(Corydalis incisa)试验中有类似结论[25]。
试验结果表明,随着郁闭度的增加,鳞尾木的保存率、株高生长量和地径生长量均随着郁闭度增大呈先上升后下降的趋势,且均优于全光照的CK 处理,其中B2 处理时保存率、株高生长量、地径生长量最大,B3处理时各指标明显降低,说明林下郁闭度有利于鳞尾木生长,最适宜的郁闭度为0.5(B2 处理),郁闭度过高则生长不良;叶面积、叶长和叶宽均呈先增加后减少,在B2处理下最大,长宽比呈先下降后上升的趋势。鳞尾木的Pn、Tr、WUE、Gs 随着郁闭度增加呈先上升后下降的趋势,B2 处理下的各数值均最大,B3 处理时显著低于B2 处理,表明在一定的郁闭度范围内,鳞尾木通过提高气孔导度来维持较高的蒸腾速率,保证光合作用及有机物的合成,但林下郁闭度过高则使气孔导度下降。鳞尾木叶片的叶绿素总量、叶绿素a、叶绿素b、类胡萝卜素均随着郁闭度的增加逐渐增加,叶绿素a/b 值则随着郁闭度的增大逐渐减小,说明鳞尾木在林下遮阴条件下合成更多的叶绿素以适应弱光环境。在B3处理下鳞尾木叶片的叶绿素含量很高,但Pn值不高,可能是与它在林下郁闭度过高的环境下,光照过弱不能充分进行光合作用有关。
为综合评价八角林不同郁闭度对鳞尾木生长效果的影响,进行了隶属函数综合评价,结果表明,B2处理下的鳞尾木幼苗隶属函数值最大,综合排名第1,鳞尾木长势最好,与实际观测结果吻合。综合分析,鳞尾木在栽培时应适当遮阴,建议选择郁闭度为0.5的林下环境进行,可有效提高成活率,促进苗木生长,林下郁闭度不合适的光照环境均不利于鳞尾木生长。
猜你喜欢
世界热带农业信息(2022年11期)2022-10-18 10:20:12
绿色科技(2021年21期)2021-11-26 09:12:06
内蒙古林业调查设计(2021年3期)2021-07-06 14:12:00
吉林林业科技(2020年6期)2020-12-16 10:09:36
福建林业(2020年5期)2020-03-18 08:23:02
热带林业(2019年4期)2019-03-05 09:53:58
森林工程(2018年3期)2018-06-26 03:40:46
现代园艺(2018年1期)2018-03-15 07:56:13
农民致富之友(2014年7期)2014-10-21 20:06:25
河南科技(2014年24期)2014-02-27 14:19:48
