
添加生物炭和氮素对酸性土壤CO2 和N2O 排放的影响
2023-06-26 06:45:32史雅童潘亚男曹文超郭景恒王娅静
河北农业大学学报 2023年3期
史雅童,潘亚男,曹文超,郭景恒,王娅静
(1.河北农业大学 资源与环境科学学院/河北省农田生态环境重点实验室,河北 保定 071000;2.中国农业大学 资源与环境学院,北京 100193;3.潍坊科技学院,山东 寿光 262700)
全球气温上升是当前环境领域关注的热点问题,其原因主要是大气中二氧化碳(CO2)、氧化亚氮(N2O)和甲烷(CH4)等温室气体(Greenhouse gas,GHG)浓度的日益增加[1]。土壤作为GHG的重要排放源,其对GHG 的贡献已引起国内外学者的高度重视。土壤排放CO2、N2O 和CH4分别主要是通过土壤呼吸和有机质的好氧分解、土壤硝化—反硝化过程及有机质的厌氧发酵,过程较复杂且受多种因素影响[2-3]。
生物炭作为一种低成本、环境友好的炭质多孔材料,在减少土壤GHG 排放方面具有潜在的重要性,已成为土壤固碳减排的研究热点之一[4-5]。生物炭具有pH 值高、孔隙发达、比表面积大等特点,将其施入土壤后可通过改变土壤理化性质或通过物理吸附、化学反应直接或间接影响GHG 排放[6]。有研究表明,添加生物炭可显著降低土壤GHG 的排放[7-8]。然而,也有研究表明添加生物炭对GHG排放无显著影响或有促进作用[9-10]。生物炭的不同作用效果可能与生物炭的施用量、种类、裂解温度以及土壤本身的理化性质等有关[11-12]。因此,进一步探索生物炭对土壤GHG 排放的影响机制尤为重要。N2O 在大气中的含量较低,但对全球变暖和平流层臭氧的影响却有重要作用[13]。土壤向大气中释放N2O 主要是通过反硝化和硝化过程,两者的相对贡献与土壤中氮素的形态有密切关系[14]。有研究表明,硝态氮和铵态氮的施用都能增加土壤N2O排放,但作用效果不同[15-17]。氮素形态对土壤N2O排放的影响尚无定论,其影响机制仍需进一步探索。全球元分析表明,低pH 值可促进土壤N2O 的排放[18]。我国南方森林土壤普遍具有较低的pH 值,同时受较高氮沉降量的影响,其对N2O 的贡献不容小觑。添加生物炭可在一定程度上增加土壤pH 值,对于减少N2O 排放有潜在意义。因此,本研究以贵州省植物园的酸性森林土壤为研究对象,向土壤中添加生物炭和不同形态的氮素,通过室内培养试验探索生物炭和氮素添加对土壤CO2和N2O 排放的影响,以期为实现土壤GHG 减排提供理论依据。
1 材料与方法
1.1 供试材料
供试土壤采自贵州省植物园(26°37′N, 106°43′E)。该地位于贵阳市北郊鹿冲关,属于亚热带湿润季风气候,年平均气温14 ℃,年平均降水量1 200 mm。采样点成土母岩为砂岩,土壤类型为黄壤。为减少土壤本底氮素的影响,根据土壤发生层采集B 层(20~40 cm 土层)土壤。经过去杂、过筛(2 mm)和充分混匀后装入密封袋中置于4℃冰箱中保存。利用常见分析方法测得土壤有机碳含量为58.23 g/kg,总氮含量为2.67 g/kg,硝态氮含量为0.01 mg/kg,铵态氮含量为2.04 mg/kg,含水率为28.67 %,pH 值为3.92。所用生物炭购于河南誉中奥农业科技有限公司,其有机碳含量为350 g/kg,pH 值为10.24,速效磷含量为10.20 g/kg,速效钾含量为55.65 g/kg,容重为0.19 g/cm3,总孔隙度为67.03 %。
1.2 试验设计
试验设置3 个生物炭水平,分别为0、30 和60 g/kg;氮素类型包括硝态氮(KNO3)和铵态氮((NH4)2SO4),均为2 个水平,分别为0 和30 mg N/kg。为明晰反硝化和硝化过程对土壤GHG 排放的贡献,添加硝态氮处理进行厌氧培养,添加铵态氮处理进行好氧培养。基于此,厌氧培养设置6 个处理:1)CK,空白对照(不添加生物炭和氮素);2)B30,仅添加30 g/kg 生物炭;3)B60,仅添加60 g/kg 生物炭;4)NN,仅添加30 mg/kg 硝态氮;5)B30-NN,添加30 g/kg 生物炭和30 mg /kg 硝态氮;6)B60-NN,添加60 g/kg 生物炭和30 mg /kg硝态氮。与厌氧培养的处理相似,好氧培养同样设置6 个处理,添加氮素为铵态氮(AN)。所有处理均设3 次重复。
1.3 试验过程
称取3 份1.0 kg 新鲜土壤分别加入0、30 和60 g生物炭,混匀后装入密封袋中于4 ℃冰箱中保存。分别称取30 g 不同生物炭水平的土壤样品于120 mL血清瓶内,用橡胶塞和铝盖密封。为进一步降低土壤本底氮素对后续试验的影响,需对样品进行预培养,即将密封后的血清瓶用充气抽真空清洗系统(北京帅恩科技有限公司)反复3 次用高纯氦气(He,99.999%)进行冲洗,在人工气候箱中恒温(20 ℃)厌氧培养5 d,然后再次进行抽真空并注入含有20%氧气的氦氧标准混合气,于人工气候箱中恒温(20 ℃)好氧培养5 d。对于厌氧培养,利用注射器向血清瓶中均匀喷洒3 mL 去离子水或浓度为300 mg N/L 的硝酸钾(KNO3)溶液,而后进行抽真空注入He。对于好氧培养,喷洒3 mL 去离子水或浓度为300 mg N/L 的硫酸铵((NH4)2SO4)溶液,而后进行抽真空注入含有20%氧气的氦氧标准混合气。用装有去离子水的注射器对所有血清瓶进行平衡气压,将血清瓶放入20 ℃恒温水浴锅中培养15 d。
1.4 指标测定
土壤理化性质采用常规分析方法进行测定,即利用元素分析仪测定土壤有机碳和总氮含量;用1.0 mol/L 的KCl 溶液浸提土壤后,利用靛酚蓝比色法测定浸提液中的铵态氮(NH4+-N)含量,重氮化偶合比色法测定浸提液中的NO2--N 含量,浸提液流经铜镉还原柱后采用重氮化偶合比色法测定NO3--N含量;采用复合电极法测定土壤pH 值(水土比为2.5∶1,即在25 mL 去离子水中加入10 g 风干土)。血清瓶中CO2和N2O 的浓度利用Robot 系统[19]进行测定,培养过程中每8 h 采集1 次气体并对其含量进行分析。
1.5 数据处理
在百年尺度上,N2O 的增温潜势是CO2的298倍[20],本研究通过以下公式计算全球增温潜势(Global warming potential,GWP)
GWP=R(CO2)+298 *R(N2O)
式中,GWP 表示CO2和N2O 气体排放的综合增温潜势(CO2-eq mg/kg);R(CO2)和R(N2O)分别表示试验结束时的CO2和N2O 累积浓度,单位为mg/kg。
使用Microsoft Excel 2016 进行数据的整理;利用SPSS 24 对数据进行统计分析;利用Origin 8.5绘图。
2 结果与分析
2.1 添加生物炭和硝态氮对土壤CO2 和N2O 排放的影响
土壤添加生物炭和硝态氮在厌氧条件下的CO2浓度变化如图1(a)所示。CK 处理CO2浓度以逐渐减小的速率缓慢增加,培养结束时增至5.74 mg C/kg。生物炭添加量为30 g/kg(B30)和60 g/kg(B60)处理的CO2浓度随时间的变化趋势与CK 相似,但增加速率明显高于CK,且B60 处理的速率明显高于B30 处理。反应结束时,B60 处理的CO2累积浓度为13.17 mg C/kg,显著(P<0.01)高于B30 处理(6.86 mg C/kg)和CK。表明添加生物炭显著(P<0.01)促进了厌氧条件下CO2的排放,且促进效果随生物炭添加量的增加而显著(P<0.01)增加。添加30 mg/kg 硝态氮(NN)处理的CO2浓度基本成线性增加,培养结束时增至6.96 mg C/kg,显著(P<0.01)高于CK。表明添加硝态氮明显促进了CO2的排放。与CK 相比,B30-NN 和B60-NN 处理具有更大的CO2排放速率,培养结束时CO2累积浓度分别增至7.18 和12.28 mg C/kg,均显著高于CK。表明同时添加生物炭和硝态氮显著(P<0.01)促进了CO2的排放。通过对比发现,在相同生物炭水平下添加硝态氮对CO2排放无显著(P>0.05)影响,在相同硝态氮水平下随着生物炭添加量的增加CO2浓度基本呈增加趋势。结合表1 可知,厌氧条件下生物炭水平和硝态氮水平均对土壤CO2累积排放量有显著影响,且生物炭水平和硝态氮水平对土壤CO2排放存在交互影响,单独添加生物炭对CO2排放的促进效果最大,其次是同时添加生物炭和硝态氮、单独添加硝态氮。

表1 生物炭和氮素水平对土壤CO2 和N2O 累积浓度影响的析因设计方差分析Table 1 Factorial design ANOVA of effects of biochar and N level on soil CO2 and N2O cumulative concentrations

图1 厌氧条件下CO2 和N2O 浓度的动态变化Fig.1 Changes in CO2 and N2O concentrations under anaerobic addition
由图1(b)可知,不同处理的N2O 浓度变化存在较大差异。CK 处理N2O 浓度随时间大致呈线性增加,培养结束时N2O 累积浓度为1.77 mg N/kg。B30处理N2O 浓度在192 h 内快速增加至2.28 mg N/kg,随后迅速下降,在280 h 降至低于0.10 mg N/kg,下降速率大于前期的增加速率。B60 处理N2O 浓度在72 h内迅速增加至1.95 mg N/kg,峰值低于B30处理,但其具有更大的产生速率。72 h 后N2O 浓度迅速下降,104 h 降至低于0.10 mg N/kg。由此可知,添加生物炭促进了N2O 的产生和进一步的还原,且促进效果随生物炭水平的增加而增加。NN 处理N2O 排放速率在104 h 内与CK 处理基本一致,而后逐渐增加,在208 h 后逐渐降低,培养结束时N2O 累积浓度为2.20 mg N/kg。结果表明,添加硝态氮促进了N2O 的排放。B30-NN 处理,N2O 浓度基本成线性增加,培养结束时N2O 的累积浓度为4.72 mg N/kg。B60-NN 处理,N2O 浓度在64 h 内以逐渐增大的速率快速增加,随后以逐渐减小的速率逐渐增加,培养结束时达到5.75 mg N/kg。对比可知,同时添加生物炭和硝态氮显著(P<0.01)促进了N2O 的排放。在相同生物炭水平下添加硝态氮显著(P<0.01)增加N2O 排放,在相同硝态氮水平下随着生物炭水平的增加N2O 浓度显著(P<0.01)增加。由表1 可知,厌氧条件下硝态氮水平对土壤N2O 累积排放量有显著影响,生物炭水平的影响不显著,生物炭水平和硝态氮水平存在交互影响。
2.2 添加生物炭和铵态氮对土壤CO2 和N2O 排放的影响
图2(a)为土壤添加生物炭和铵态氮在好氧条件下的CO2浓度变化。CK 处理CO2浓度以逐渐降低的速率缓慢增加,培养结束时增至7.31 mg C/kg。与CK 处理相比,B30 和B60 处理具有更高的CO2排放速率,培养结束时CO2累积浓度分别为8.69 和11.46 mg C /kg。表明添加生物炭促进了好氧条件下CO2的排放,且促进效果随生物炭添加量的增加而显著(P<0.01)增加。AN 处理CO2浓度随时间的变化趋势与CK 相似,但排放速率高于CK,培养结束时CO2累积浓度为7.56 mg C/kg,显著(P<0.05)高 于CK。B30-AN 和B60-AN 处 理 与CK 处 理 相比具有更大的CO2排放速率,且B60-AN 处理的排放速率大于B30-AN 处理。培养结束时,B30-AN、B60-AN 处理的CO2累积浓度分别为8.56 和11.78 mg C/kg,表明同时添加生物炭和铵态氮显著(P<0.01)促进了CO2的排放。此外,相同生物炭水平下添加铵态氮对CO2排放无显著(P>0.05)影响,相同铵态氮水平下随着生物炭水平的增加CO2浓度显著(P<0.01)增加。由表1 可知,好氧条件下生物炭水平对土壤CO2累积排放量有显著影响,铵态氮水平的影响不显著,且生物炭水平和铵态氮水平无交互作用。

图2 好氧条件下CO2 和N2O 浓度的动态变化Fig.2 Changes in CO2 and N2O concentrations under aerobic addition
如图2(b),好氧条件下各处理的N2O 排放量均较低(<0.020 mg N/kg)。在CK、B30 和B60 处理,N2O 浓度随时间的变化趋势基本一致,即N2O 排放量在72 h 内缓慢增加,随后基本保持不变。不同的是,72 h 内的增加速率随生物炭水平的增加而增加。培养结束时CK、B30 和B60 处理的N2O 累积浓度分别是0.003、0.009 和0.012 mg N/kg,表明添加生物炭显著(P<0.05)促进了N2O 排放。与CK 相似,AN 处理的N2O 排放速率在72 h 内缓慢增加,随后基本保持不变。培养结束时,N2O 累积浓度为0.003 mg N/kg,与CK 处理差异不显著(P>0.05)。培养结束时,B30-AN、B60-AN 处理的N2O 累积浓度分别为0.009 和0.012 mg N/kg,显著(P<0.01)高于CK 处理。对比可知,相同生物炭水平时添加铵态氮对N2O 排放无显著(P>0.05)影响,相同铵态氮水平时N2O浓度随生物炭水平增加而显著(P<0.01)增加。由表1 可知,好氧条件下生物炭水平对土壤N2O 累积排放量有显著影响,铵态氮水平的影响不显著,且生物炭水平和铵态氮水平无交互作用。
2.3 添加生物碳和氮素对全球增温潜势的影响
如表2 所示,添加生物炭对全球增温潜势的影响在厌氧条件和好氧条件下有相反的结果。
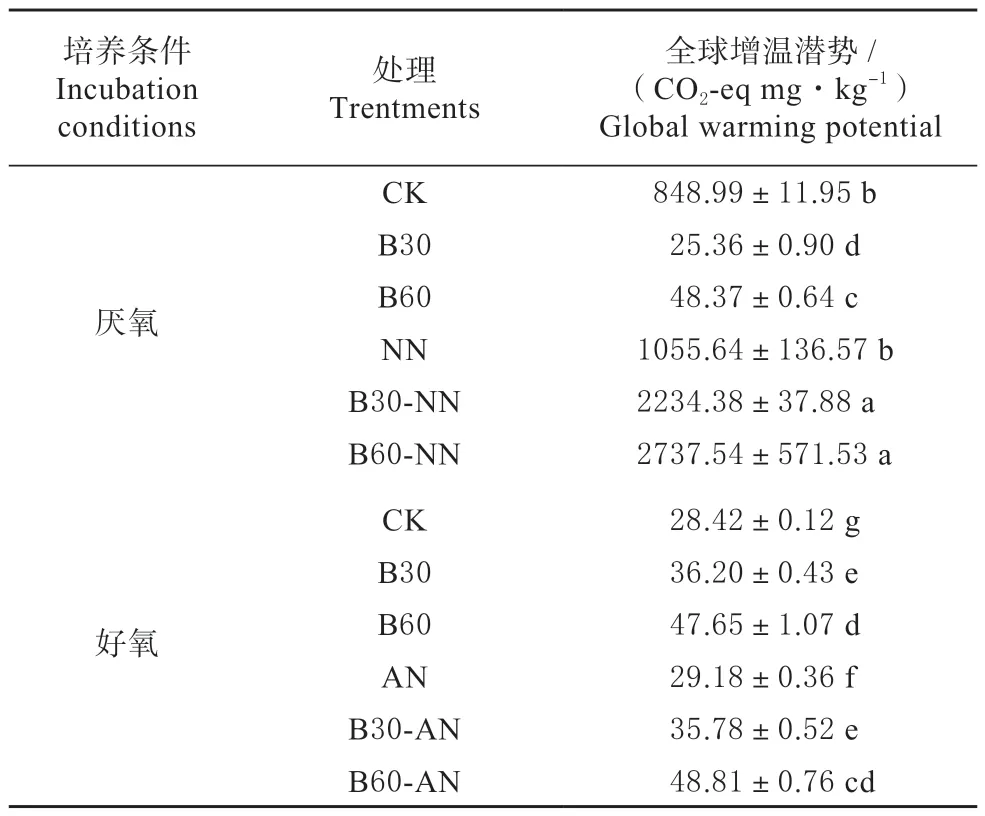
表2 不同处理的全球增温潜势Table 2 Global warming potential of different treatments
厌氧条件下,B30 和B60 处理的全球增温潜势分别比CK 处理降低了97.01% 和94.30%。然而,好氧条件下分别增加了27.38% 和67.66%。添加氮素在厌氧条件和好氧条件下均使全球增温潜势增加,分别增加了24.34%和2.67%。同时添加生物炭和氮素也使全球增温潜势显著增加,厌氧条件和好氧条件下分别增加了163.18%~222.45%和24.60%~71.75%。
3 讨论
试验结果表明,添加生物炭促进了厌氧和好氧条件下土壤CO2的排放,使CO2累积排放量增加了18.88%~129.44%,且促进效果随着生物炭水平的增加而增加。该结果与已有研究结果一致[21-22]。陶甄等[21]通过室内培养试验发现添加生物炭使土壤CO2累积排放通量增加14.29%。涂保华等[22]通过盆栽和大田试验发现添加生物炭使稻田土壤CO2排放量增加13.06%~26.59%。生物炭促进土壤CO2排放可能是由于生物炭本身含有的可溶性有机碳分解或生物炭添加到土壤后促进了土壤原有机碳的分解[23]。此外,生物炭添加到土壤后可通过影响土壤理化性质,如提高土壤pH 值和改善土壤通气性,进而提高微生物活性和土壤呼吸强度[24]。然而,也有研究表明添加生物炭会抑制土壤CO2排放[7]。究其原因,可能是:生物炭具有较大的比表面积和孔隙度,对土壤有机质、微生物、酶、CO2均存在着一定的物理吸附效应;生物炭中的矿物成分可与CO2结合形成碳酸盐,从而减少CO2的排放[6,24]。本研究在厌氧条件下添加生物炭显著促进了N2O 的产生和进一步的还原,使得反应结束后添加生物炭处理的N2O 累积排放量显著低于空白。该结果与涂保华等对水稻土进行的盆栽试验结果一致,发现添加生物炭使土壤N2O 累积排放量显著降低了17.20%~27.96%[22]。本研究在好氧条件下添加生物炭促进了N2O 的不断产生和释放,且在72 h 内N2O 的增加速率随着生物炭添加量的增加而增加。添加生物炭对N2O 排放的促进效果可能归因于提高了N 的有效性或增加了细菌amoA基因拷贝数,从而促进了反硝化或硝化过程产生N2O[10]。添加生物炭促进N2O 的进一步还原,主要是通过提高土壤pH 值,进而增强了N2O 还原酶的活性,促进N2O向N2的转化[13]。通过对比厌氧和好氧条件下添加生物炭对土壤N2O 累积排放量的影响可知,添加生物炭在厌氧条件下对土壤N2O 累积排放有抑制作用,而在好氧条件下对土壤N2O 累积排放有促进作用。
试验发现,在厌氧条件下添加硝态氮有效促进了N2O 的排放,在好氧条件下添加铵态氮对N2O 排放无显著影响。表明本试验条件下更有利于通过反硝化过程产生N2O。同本试验结果不同,有研究发现添加铵态氮比添加硝态氮更能促进温带森林和草原土壤N2O 的排放,此时硝化过程是土壤N2O 排放的主导过程[25]。目前,氮素类型对土壤N2O 排放的影响尚无定论,关于土壤N2O 排放是以硝化过程为主还是反硝化过程为主仍存在争论[26]。本研究通过室内培养试验表明所用森林土壤更有利于通过反硝化过程产生N2O,然而原位条件下的实际情况仍需进一步通过原位观测并结合15N 同位素示踪技术和分子生物学方法量化硝化和反硝化过程对土壤N2O 排放的贡献。
研究表明添加生物炭对土壤理化性质有重要影响[8,27]。本研究中添加生物炭30 和60 g/kg 后,分别使土壤pH 值增加了1.17 和2.28。土壤pH 的改变会进一步影响有机质的分解、微生物的活性、酶的活性,从而影响GHG 的排放。陈红卫等[27]向农田土壤中施加生物炭10~30 t/hm2使土壤pH 增加了0.56%~9.66%,土壤全氮含量增加了1.66%~5.89%,土壤容重降低了1.63%~10.40%。生物炭可以直接通过物理吸附或化学反应影响土壤GHG 排放的影响,也可以通过影响土壤理化性质间接影响。此外,生物炭本身的理化性质、生物炭种类、生产过程的不同对GHG的减排效果也不尽相同[12]。生物炭对GHG 排放的影响机制仍需进一步从土壤物理、化学和生物学多角度开展相关研究,以期为土壤GHG减排提供理论依据。
4 结论
对贵州省植物园酸性森林土壤的GHG 排放的初步研究结果表明:
(1)添加生物炭促进了厌氧和好氧条件下土壤CO2的排放,且促进效果随生物炭添加量的增加而增加,向土壤中添加30 和60 g/kg 生物炭使厌氧条件下CO2累积浓度分别增加了19.51%和129.44%,好氧条件下分别增加了18.88%和56.77%。
(2)添加生物炭显著降低了(P<0.01)厌氧条件下N2O 的累积浓度,生物炭添加量为30 和60 g/kg分 别 降 低 了99.97% 和99.99%;显 著(P<0.05)增加了好氧条件下N2O 的累积浓度,分别增加了170.82%和252.14%。
(3)添加氮素对土壤CO2和N2O 的排放均有重要影响,添加硝态氮和铵态氮分别使土壤CO2累积浓度增加了21.25%和31.71%,N2O 累积浓度增加了24.29%和0.00%。
(4)同时添加生物炭和氮素存在交互作用,但对土壤CO2和N2O 排放的促进效果低于仅添加生物炭或仅添加氮素处理。
(5)由于培养结束时厌氧条件下添加生物炭处理N2O 基本被完全消耗,使得厌氧条件下添加30和60 g/kg 生物炭分别降低了97.01%和94.30%的全球增温潜势,其余处理均使全球增温潜势显著增加。表明生物炭对于酸性土壤厌氧环境下的GHG 减排具有潜在的重要作用,应引起广泛重视。
猜你喜欢
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29 03:15:14
农业环境科学学报(2020年10期)2020-11-11 02:55:28
环境保护与循环经济(2017年7期)2018-01-22 02:58:12
环境污染与防治(2016年12期)2016-03-13 05:52:12
中国环境科学(2016年3期)2016-02-08 15:07:14
湖泊科学(2015年1期)2015-05-06 07:14:56
应用海洋学学报(2014年2期)2014-11-26 01:20:46
植物营养与肥料学报(2011年5期)2011-11-06 07:30:02
植物营养与肥料学报(2011年4期)2011-10-26 02:44:56
植物营养与肥料学报(2010年3期)2010-11-16 00:59:24
