
胡桃楸果实表型性状及遗传多样性分析
2023-06-19 12:04王子豪苏迅解孝满仝伯强丁平韩彪刘丹张萍孔冬瑞胡德昌
山东农业科学 2023年5期
王子豪,苏迅,解孝满,仝伯强,丁平,韩彪,刘丹,张萍,孔冬瑞,胡德昌
(1. 鲁东大学生命科学学院,山东烟台 264025;2. 山东省林草种质资源中心,山东济南 250014)
胡桃楸(Juglans mandshuricaMaxim.)为胡桃科(Juglandaceae)胡桃属(JuglansL.)植物,又称核桃楸、野核桃,是一种高大温带落叶乔木,常作材用,与水曲柳和黄波罗并称东北地区三大阔叶树种,且种仁可食[1,2]。 作为核桃的近缘种,与栽培核桃杂交育种、用作核桃砧木等成为近年胡桃楸研究的热点[3-5]。 另外,Xu 等[6]揭示了核桃青皮酚类物质能够有效抵御胶孢炭疽菌,对下一步开发植物源核桃炭疽病绿色防控农药具有重要意义。 因此,对胡桃楸进行相关研究具有巨大实际应用潜力。
果实作为繁殖器官,与叶片等营养器官相比特征相对稳定,能在不同地理位置、复杂多变的环境下表现出较强的适应性,能较好地体现进化上的变异规律。 陈思羽等[7]发现分布在长白山不同区域的核桃楸,种子质量、种子长度和种子宽度等性状变异明显。 宋佳兴等[8]将辽东山区胡桃楸种质资源的果实根据表型性状划分为球形、卵形等十余种果实形态,变异类型丰富。 高张莹等[9]认为不同种群间核桃楸果核性状存在极显著差异,主要受生境因子影响,但也受到很强的遗传控制,基于表型性状数据的聚类结果与地理分布并不完全一致,呈现随机变异特征。 李红莉等[10]研究得知黑龙江野生毛榛果实表型性状多样性丰富。 张深梅等[11]调查发现大别山19 个核桃野生居群的果实表型性状在居群间和居群内均存在极显著差异,具有丰富的表型变异。 董胜君等[12]发现东北杏的果实性状变异较为丰富,且变异主要来源于居群间。 王尧等[13]研究得知山东省盐肤木果实表型性状多样性较为丰富,变异以居群间变异为主,而且受地理位置影响明显,可作为盐肤木优质种源筛选的重要指标。 可见,以果实作为试验材料能更直观地反映胡桃楸各个居群间的表型差异。
胡桃楸主要分布于我国东北及华北地区,然而近几年的资源普查发现山东省的泰山、仰天山、昆嵛山等地也有部分自然种群分布,呈现狭域分布格局。 目前,该物种已被列为山东省珍稀濒危植物,相关的物种保护工作也已开展[14-16]。 由于个体数量少且结实率低、啮齿类动物取食以及人为采集等因素影响,获取一定数量的群体样本和果实材料相对困难,这导致胡桃楸种质资源相关研究甚少。 前期研究结果表明,各居群胡桃楸存在一定程度的表型性状分化,遗传背景不明确。因此,为探究不同种源地胡桃楸果实变异规律及造成其形态学特征差异的原因,本研究采集山东地区3 个生境的地理居群以及胡桃楸原生地吉林地区的长白山居群共13 个单株的289 枚胡桃楸果实,测定其表型性状,并通过差异性、相关性及聚类分析研究其变异程度,同时,基于ITS 区域测序获得的生物学信息检测其遗传多样性水平,挖掘各居群的遗传背景,以期为胡桃楸种质资源的分类鉴定、物种保护、资源圃建立以及核桃新品种选育等提供参考。
1 材料与方法
1.1 果实材料采集
本试验对山东地区胡桃楸主要居群泰山居群(TS,117.10°E、36.25°N,山东泰安)、仰天山居群(YTS,118.45°E、36.71°N,山东潍坊)、昆嵛山居群(KYS,121.74°E、37.30°N,山东烟台)以及吉林地区长白山居群(CBS,121.08°E、38.46°N,吉林延边)分别进行单株取样;居群内各供试植株相距100 m 以上,以避免克隆单株。 于2021 年9 月底到10 月初果实成熟期采集成熟果实,分别编号并带回实验室,人工剥离青皮获得内果实,蒸馏水清洗干净后,置于通风良好处室温条件下风干,然后装入自封袋保存备用。
共收集到4 个胡桃楸居群13 个单株的289枚果实(表1)。 由于结实数量存在差异,从各单株采集的果实数量不同,部分单株已达取样极大值。 其中,泰山居群6 个单株140 枚果实,仰天山居群2 个单株60 枚果实,昆嵛山居群2 个单株46枚果实,长白山居群3 个单株43 枚果实。

表1 试验材料
1.2 DNA 提取
于各供试单株上分别采集成熟叶片3 ~5 枚,迅速置入盛有变色硅胶的封口塑料袋内带回实验室。 随机抽取泰山居群的4 个单株、仰天山居群的2 个单株、昆嵛山居群的2 个单株、长白山居群的3 个单株共计11 份材料进行ITS 区域测序分析。 应用CTAB 法进行DNA 提取,并通过紫外分光光度计和琼脂糖凝胶电泳检测其浓度及质量,-20℃保存备用。
1.3 果实表型性状数据获取与统计分析
使用游标卡尺(精准度±0.2 mm)测量胡桃楸果实的横径(垂直于纵轴无棱处最大横截面的长度)、侧径(垂直于纵轴的2 条纵棱之间的长度)与纵径(果实纵轴的长度),精确到0.001 mm,并计算果型指数(果型指数=纵径/横径),果型指数0.9~1.1 为圆形,>1.1 ~≤1.5 为椭圆形,>1.5 为长椭圆形。 人工观察判断果核尖端类型、表面纹饰、纵脊数目等特征,果核尖端类型分为长尖、渐尖、无尖三种,表面沟纹分为深、浅两种。
运用Microsoft Excel 2016 软件进行数据处理,采用SPSS 25.0 软件进行统计分析。 不同居群果实表型性状差异采用单因素方差分析(ANOVA),用Duncan’s 最小显著差异法进行多重比较,显著、极显著水平分别设置为0.05、0.01;将数据进行标准化转换,采用组间联结-系统聚类法,以欧式平方距离进行聚类分析;采用Pearson 法进行相关性分析。
变异系数CV =S/X×100%,其中S 为性状标准差,X 为性状平均值;
相对极差RR =(Xmax- Xmin)/X×100%,其中Xmax、Xmin分别为性状最大、最小值;
居群遗传力H2=1-1/F,其中F 为方差分析中的F 检验值;
三径均值D=(D纵径+D横径+D侧径)/3;
果实体积V=4/3π(1/2D)3。
1.4 ITS 片段PCR 扩增与数据分析
应用PCR 技术特异扩增11 份胡桃楸材料相关ITS 片段,并进行序列分析,获得遗传信息,检测变异程度。 引物序列(ITS4:5'-TCCTCCGCTTATTGATATGC - 3'; ITS5: 5' - GGAAGTAAAA GTCGTAACAAGG-3')、扩增体系及扩增程序参考张浩[17]的方案。 引物合成及所获得的PCR 产物单向测序委托生工生物工程(上海)股份有限公司进行。 利用MEGA-X 软件对序列信息进行比对,分别计算各片段的碱基含量、变异位点数目、简约性信息位点等信息。 运用DNAsp 5 软件计算ITS 序列单倍型多样性指数(Hd)、核苷酸多样性指数(Pi)以及平均核苷酸多样性差异(K)等相关遗传多样性指数;应用Arlequin 3.5 软件对ITS 区域遗传信息进行分子方差(AMOVA)分析,计算其遗传分化系数(FST)及基因流(Nm),检测胡桃楸居群内和居群间的遗传变异关系。
2 结果与分析
2.1 不同居群胡桃楸果实表型性状差异性分析
如图1 所示,山东地区不同居群胡桃楸果实表面纹饰均较浅,多为8 个纵脊、具长尖(仅泰山居群17 枚果实具渐尖和无尖),而吉林地区长白山居群多为表面纹饰较深、8 个纵脊、具长尖。 经比较,所收集的泰山居群果实体积中等,果型变化丰富,140 枚果实中,57.9%(81 枚)为椭圆形,30.0%(42 枚)为长椭圆形,12.1%(17 枚)为圆形;仰天山居群胡桃楸果实体积偏大,51.7%(31枚)为椭圆形,48.3%(29 枚)为长椭圆形;昆嵛山居群果实体积整体较小,84.8%(39 枚)为椭圆形,15.2%(7 枚)为长椭圆形;长白山居群胡桃楸果实体积最大,32.6%(14 枚)为椭圆形,67.4%(29 枚)为长椭圆形。
4 个居群胡桃楸果实的横径、侧径、纵径、果型指数、三径均值、体积变幅分别为1.85 ~3.15 cm、1.89~3.25 cm、2.17 ~5.17 cm、1.00 ~2.13、2.03 ~3.96 cm、4.38~32.50 cm3,平均值分别为2.39 cm、2.51 cm、3.34 cm、1.40、2.77 cm、11.56 cm3,变异系数在7.56%~36.07%之间,经F 检验居群间差异均达极显著水平(P<0.01),遗传力分别为0.93、0.97、0.98、0.97、0.98、0.98(表2),推断这些果实表型性状受较强的遗传特性控制,居群间差异相对稳定。
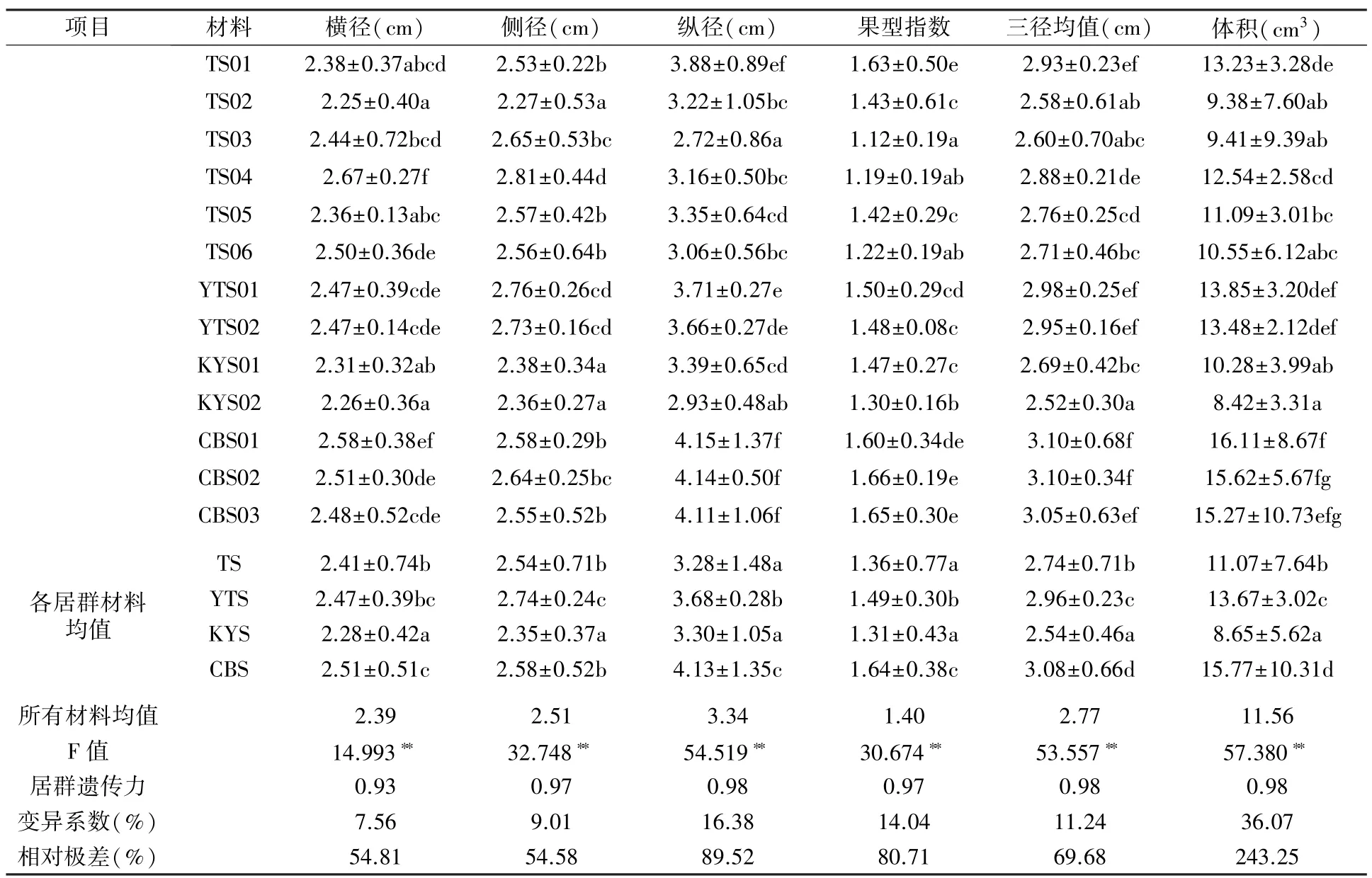
表2 不同居群胡桃楸果实表型性状统计分析结果
不同居群中,各单株果实表型性状的变化程度存在差异。 对比果型指数,CBS 与YTS 居群内个体间无显著差异,果实性状相似度较高;极值(最大2.14,最小0.93)均出现在TS 居群材料内,说明该居群个体间差异明显,果实类型最为丰富,其中TS01 与TS02、TS05 单株之间差异显著,TS03、TS04、TS06 之间差异不显著。
2.2 胡桃楸果实表型性状间的相关性分析
由表3 可知,胡桃楸果实的横径、侧径、纵径、体积、三径均值间均呈极显著正相关关系,果型指数与果实纵径、三径均值、体积均极显著正相关,表明果实横径、侧径、纵径的变异具有互相适应的协同变化特征。
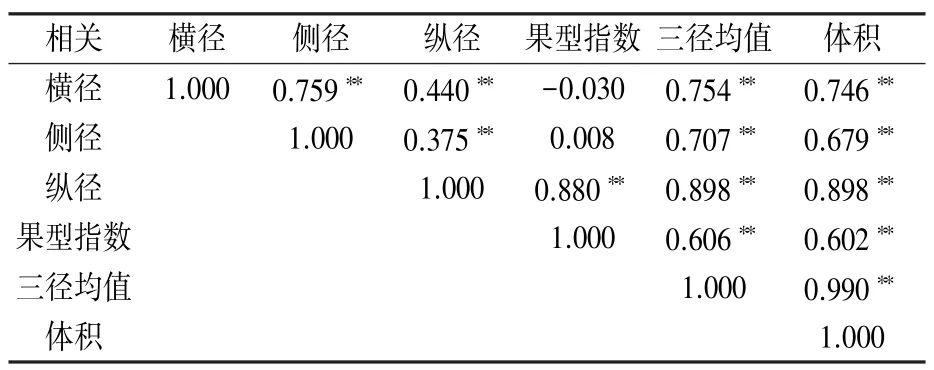
表3 胡桃楸果实表型性状间的相关性系数
2.3 基于果实表型性状的不同居群胡桃楸材料聚类分析
根据胡桃楸果实表型性状数据,运用欧氏距离类平均法进行聚类分析,结果(图2)显示,在欧氏距离25.0 处,可将胡桃楸居群分为2 组,第1组包括 CBS01、 CBS02、 CBS03、 TS01、 TS04、YTS01、YTS02,第2 组包括TS02、TS03、TS05、TS06、KYS01、KYS02。 可见泰山居群内多样性更丰富,呈现一定程度的分化,分别与长白山居群、昆嵛山居群有一定程度的相似。 供试材料间欧氏距离相对较小,相互混杂,推断山东地区胡桃楸不同居群间以及与长白山居群间存在着基因交流。
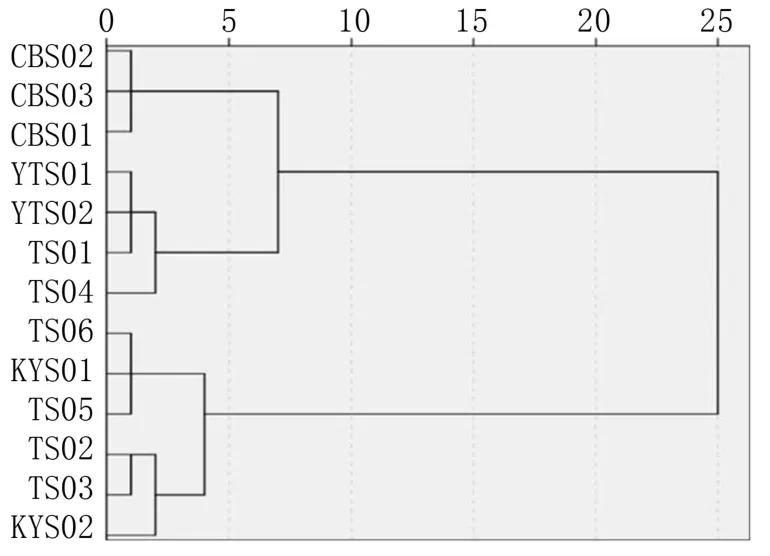
图2 基于欧氏距离构建的不同居群胡桃楸果实表型特征聚类图
2.4 基于ITS 序列的不同居群胡桃楸遗传分析
由表4 可见,YTS 居群的Hd、Pi、K 值均为零,其余3 个居群的Hd 均为1.000、Pi 和K 均表现为CBS>KYS>TS,表明YTS 居群遗传多样性水平最低,TS 和KYS 居群遗传多样性相对偏高,CBS 居群遗传多样性最高。 测序信息经比对后,不同材料的碱基变异位点无明显规律,呈现随机变化。 综合上述结论,果实表型性状分析结果与遗传分析结果基本一致。 因此,推断山东地区3个胡桃楸居群以及东北地区CBS 居群各个体的表型性状与遗传背景存在一定程度的相关性。

表4 基于ITS 序列的胡桃楸居群遗传多样性
由遗传距离分析结果(表5)可知,4 个胡桃楸居群间遗传距离普遍较小(0.001~0.048),平均值为0.027,其中TS 与KYS 居群遗传距离相对较远,呈现最大遗传距离(0.048);而TS 与YTS 居群遗传距离相对较近,遗传距离最小(0.001)。CBS 居群与TS、YTS、KYS 居群地理距离较远,但居群间遗传距离较小,表明各胡桃楸居群间的遗传距离与地理距离间未表现明显的相关性。

表5 基于ITS 序列的胡桃楸居群间遗传距离
应用Arlequin 3.5 软件对ITS 区域遗传信息进行分子方差(AMOVA)分析,结果(表6)显示该物种群体总遗传变异中有41.99%发生在居群内,58.01%的变异发生在居群间。 3 个胡桃楸居群间的遗传分化系数FST为0.665,居群间基因流Nm较大,为1.93,表明胡桃楸居群间存在基因交流。

表6 胡桃楸居群ITS 序列AMOVA 分析
3 讨论与结论
植物个体的表型性状是基因型与环境共同作用的结果。 变异系数越大说明对环境适应性越强,变异系数小则说明性状相对稳定,遗传稳定性较好,不易受到环境等因素的影响。 胡桃楸果实体积、形状等相关指标均属于数量性状,与个体生长的立地生境和树木生理年龄等因素存在一定相关性。 于国斌[18]发现东北地区胡桃楸果实存在丰富的变异类型,不同种源地的胡桃楸果实横径、纵径、侧径之间有显著性差异,推断变异类型受多种环境因素影响。 Cosmulescu 等[19]对核桃属J.regia的坚果形态学特征进行比较后认为所观察的性状变异可能是由农业气候条件和果实自身繁殖特性引起。 本研究结果表明,胡桃楸果实6 个表型性状的变异系数在7.56%~36.07%之间,在居群间存在极显著差异,4 个居群的地理位置、气候条件和种群大小存在较大差异可能是导致其果实性状变异的主要原因;另外,泰山居群的果实类型最为丰富,表明居群内的单株间也存在一定程度的遗传变异;与逄宏扬等[20]的研究结果一致,本研究也发现胡桃楸果实横径、侧径、纵径之间存在极显著正相关关系,表明这三个性状间具有互相适应的协同变化特征。 丰富的形态多样性可为后期进行胡桃楸资源收集和种质保存提供研究基础。
聚类分析结果显示,泰山居群胡桃楸果实表型性状呈现一定程度的分化,分别与长白山和仰天山居群、昆嵛山居群的果实表型较为相似,而且居群间遗传距离较小,判断可能存在着基因交流;而仰天山居群与昆嵛山居群间表型性状差异较大,未聚在一起。 表明这4 个居群的胡桃楸果实形态特征呈现片断化分布,这与高张莹等[9]的核桃楸果核形态特征在部分地区呈现区域板块化特征的结论一致。
当遗传基础较窄时,为适应复杂的生态环境,物种的表型性状有可能会随着外界环境条件的变化而产生与之相适应的遗传变异。 宋佳兴等[8]调查发现立地条件相对一致的胡桃楸果实表现出丰富的变异类型,推断胡桃楸种群具有丰富的遗传多样性。 高张莹等[9]研究发现东北到华北天然分布的9 个核桃楸种群间果核性状存在极显著差异,且各性状的遗传力均大于0.9,受到很强的遗传控制,各表型性状并未完全按地理距离而聚类。 本研究基于ITS 序列分析结果显示,山东地区的仰天山居群遗传多样性水平最低,泰山居群、昆嵛山居群和长白山居群遗传多样性相对偏高;基于形态学特征分析的结果也表明仰天山居群果实表型性状较为单一,泰山居群的性状变异相对丰富。 两方面分析所得结论基本一致,推断胡桃楸果实表型性状的差异是其适应不同生境条件的方式之一,且这种变异是不连续的,可能受遗传多样性影响。 因此,若要完全阐明胡桃楸果实表型变异规律,还需进一步扩大种质资源的收集范围,并结合分子手段对其遗传多样性和亲缘关系等进行深入研究。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
疯狂英语·初中天地(2019年10期)2019-10-23
草地学报(2018年5期)2018-11-07
现代园艺(2017年21期)2018-01-03
学与玩(2017年4期)2017-02-16
小学教学研究(2016年36期)2016-05-17
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
