
基于细胞焦亡途径探讨中医药抗肿瘤机制研究进展*
2023-06-19 01:20岳亚楠柳青峰
中医学报 2023年7期
岳亚楠,柳青峰
1.辽宁中医药大学,辽宁 沈阳 110000; 2.辽宁省金秋医院,辽宁 沈阳 110000
细胞焦亡是一种最初被认为是与免疫沉默的细胞凋亡相似且依赖caspase-1介导的程序性细胞死亡。随着研究的深入,细胞焦亡被重新定义为细胞肿胀伴随细胞膜破裂并释放炎性因子的新型促炎性细胞坏死形式。除了两条经典或非经典途径,凋亡半胱天冬酶、颗粒酶等介导的细胞焦亡新通路也逐渐被发现。
细胞焦亡在心肌缺血再灌注损伤、阿尔茨海默病、自身免疫性疾病等多种疾病中的作用逐渐受到关注,在肿瘤发生、发展及转归中的双刃剑作用及与肿瘤免疫的密切联系也得到了证实,诱导细胞焦亡从而进一步抗肿瘤已成为肿瘤领域的研究热点。细胞焦亡本身与中医阴阳学说的联系也为中医药调控该途径提供了理论依据,随着中药中各类化合物对细胞焦亡通路的靶向机制研究越来越多,对中医药抗肿瘤也有了新的探索。
1 细胞焦亡概述与机制
细胞焦亡是由GSDM 蛋白介导的区别于非炎症性细胞凋亡的一种促炎性程序性死亡方式。与细胞凋亡过程中细胞皱缩后死亡但细胞膜结构完整的特征不同的是,细胞焦亡表现为膜破裂、细胞肿胀及可激活炎症反应的细胞内容物的释放。这种独特的细胞死亡模式最初于1992年在福氏志贺氏菌感染的小鼠巨噬细胞中被发现,后将其归为细胞凋亡。1997年,研究发现福氏志贺氏菌诱导的“凋亡”过程中负责切割多向性促炎因子白细胞介素-1β(interleukin-1β,IL-1β)的caspase-1可被激活,直至2001年被正式描述为“焦亡”[1-3]。细胞焦亡早期被Cookson等认为是caspase-1依赖性单核细胞死亡,但后续研究发现,小鼠caspase-11和其人类同源蛋白caspase-4、caspase-5在识别胞浆脂多糖时可直接结合脂多糖引起激活从而触发焦亡,且有学者通过全基因组编辑筛选确定了GSDM家族成员之一GSDMD是caspase-1、caspase-4、caspase-5、caspase-11的通用底物,活化的炎性半胱天冬酶对GSDMD进行切割裂解,并通过释放具有膜打孔活性的寡聚化的GSDMD-N端结构域破坏细胞膜,从而引发细胞焦亡,GSDMD是细胞焦亡最关键的蛋白分子[4-5]。此外,GSDM家族成员GSDME等也被证明参与其中,因此细胞焦亡的定义转变为由GSDM介导的程序性死亡方式。
由GSDMD介导的细胞焦亡途径分为依赖caspase-1的经典焦亡途径和依赖caspase-4、caspase-5、caspase-11的非经典焦亡途径。在经典焦亡途径中,炎性caspases-1的激活是由多分子蛋白复合物炎症小体的组装介导的,而炎症小体的代谢活化主要由模式识别受体(pattern recognition receptors,PRRS)识别病原体相关分子模式(pathogen associated molecular patterns,PAMPs) 或宿主细胞胞质中的内源性危险信号启动。激活后的炎症小体与凋亡相关斑点样蛋白(apoptosis-associated speck-like protein containing a CARD,ASC)和caspase-1的前体形成复合物,促进caspase-1前体的自我剪切,从而得到活性caspase-1;被活性caspase-1切割后的GSDMD-N与细胞膜磷脂酰肌醇、磷脂酸和磷脂酰丝氨酸结合,使细胞膜形成膜孔,导致细胞肿胀破裂;被活性caspase-1切割得到的成熟IL-1β以及IL-18也可通过膜孔释放出来,促进炎症反应的激活[6-7]。而非经典焦亡途径中,caspase-4、caspase-5、caspase-11除了同革兰氏阴性菌脂多糖结合从而引起非经典焦亡外,活性caspase-11还可以切割并激活pannexin-1通道以诱导ATP释放,并通过GSDMD 切割形成的细胞膜膜孔促使K+外排,以激活NLRP3炎症小体,从而诱导细胞焦亡发生[8]。
研究发现,GSDM 家族另一成员GSDME可被肿瘤坏死因子-α(tumour necrosis factor-α,TNF-α)或化疗药物激活的caspase-3切割而产生GSDME-N片段,导致某些GSDME高表达的癌细胞由细胞凋亡向细胞焦亡转变[9]。另外,Du等[10]研究发现自然杀伤(natural killer,NK)细胞中的颗粒酶B可以在D270位点后直接裂解GSDME,并间接激活caspase-3,从而切割D270位点以诱导肿瘤靶点的GSDME依赖性焦亡,这一发现可能为克服肿瘤逃避免疫提供了新的思路。
2 细胞焦亡与肿瘤
2.1 细胞焦亡与肿瘤发生发展随着研究的不断深入,细胞焦亡在包括肿瘤在内的各系统疾病中的参与逐渐被人们了解。细胞焦亡影响肿瘤发生发展的各个阶段,并在肿瘤微环境中发挥着抑瘤或促瘤的双向调节作用。一方面,在焦亡过程中,炎症小体的激活及炎症因子的释放能够导致多组织炎症扩散,炎症性肿瘤微环境的形成对某些肿瘤的生成和转化具有促进作用,并与其不良预后密切相关[10]。Boone等[11]研究显示,在胰腺癌原位肿瘤模型中,炎症小体NLRP3表达上调,且其信号通路促进了血小板的聚集,其活化和聚集水平的升高可以促进胰腺的肿瘤生长。Daley等[12]研究显示,胰腺导管腺癌的发展有赖于巨噬细胞中NLRP3信号对其肿瘤微环境中的免疫抑制CD4+T细胞极化的支持。另一方面,适度的细胞焦亡可抑制肿瘤细胞的增殖,并且与免疫沉默的细胞凋亡不同,细胞焦亡过程中触发的可控的炎症反应可为激活获得性免疫系统从而产生抗肿瘤免疫提供前提。研究表明,敲除GSDME促进了肿瘤生长,故GSDME可作为肿瘤抑制因子,通过诱导细胞焦亡增强抗肿瘤免疫能力,且其表达可提高肿瘤浸润的NK细胞和CD8+T淋巴细胞的数量和功能[13]。综上,尽管对细胞焦亡认识的深入为抗肿瘤研究提供了新的方向,但如何把控细胞焦亡的程度以及发挥细胞焦亡对肿瘤微环境的有利影响仍是研究者面临的重要问题。
2.2 细胞焦亡与抗肿瘤治疗
2.2.1 细胞焦亡与抗肿瘤化疗治疗继研究发现多柔比星和顺铂可通过激活caspase-3诱导表达GSDME的神经纤维瘤SH-SY5Y细胞或黑色素瘤 MeWo细胞出现依赖性焦亡后,紫杉醇等化疗药物通过介导经典/非经典以及颗粒酶等焦亡途径诱发多种癌细胞焦亡的研究成果越来越多。Zhang等[14]通过细胞生存能力分析、裂解细胞死亡实验等发现,顺铂及紫杉醇均能引起caspase-3的激活和GSDME-NT的生成,从而诱导表达GSDME的一些A549肺癌死亡细胞出现继发性坏死/焦亡的特征性形态,且顺铂诱导焦亡的能力远高于紫杉醇。Yan等[15]在研究中纳入20例基于顺铂的新辅助化疗的三阴性乳腺癌患者,选取进行新辅助化疗前的穿刺活检组织、进行新辅助化疗后切除的标本组织以及三阴性乳腺癌细胞株MDA-MB-231进行研究,结果显示,化疗后的标本组织与化疗前的标本比较,不仅焦亡效应分子GSDMD表达明显升高,MEG3、NLRP3、caspase-1、IL-18和IL-1β表达也呈上调趋势;顺铂作用后的MDA-MB-231细胞株GSDMD-FL及GSDMD-N表达上调,MEG3、NLRP3、pro-caspase-1和caspase-1的裂解(C-caspase-1)均升高,且IL-18、IL-1β的积累呈时间依赖性,而敲除MEG3可抑制顺铂对细胞焦亡的激活作用。Wang等[16]研究证明,经5-氟尿嘧啶处理后,胃癌细胞SGC-7901和MKN-45出现增殖抑制以及被激活的caspase-3切割而产生的剂量依赖性的GSDME裂解,从而发生焦亡。
2.2.2 细胞焦亡与抗肿瘤免疫治疗肿瘤免疫疗法依靠对免疫微环境的重塑和对免疫逃逸的有效抑制重新激活人体抗肿瘤免疫反应,已成为研究前景广阔的一种新兴疗法,而细胞焦亡与肿瘤免疫的复杂联系使其在抗肿瘤免疫中的机制及应用越来越受到关注。细胞焦亡对免疫激活的正向作用不仅与细胞焦亡释放的炎症介质触发天然免疫反应并招募免疫T细胞浸润从而促使效应T细胞的免疫增强有关,而且与淋巴细胞发挥细胞毒性作用使肿瘤细胞诱导TME进入免疫刺激状态有关[10]。Wang等[17]将生物正交化学体系应用于GSDM,使GSDM有选择性地释放到肿瘤细胞,结果显示,不到15%的肿瘤细胞发生焦亡便可清除基于4T1细胞所建立的乳腺肿瘤移植物,且免疫细胞数量显著增加,但当小鼠为免疫缺陷或T细胞衰竭小鼠时肿瘤不会被抑制,同时提出焦亡触发的炎症可与免疫检查点抑制剂协同作用。Hou等[18]研究表明,经典的免疫检查点抑制剂PD-L1在缺氧条件下可与活化的STAT3结合,促进核易位以及GSDMC基因转录的形成,而被caspase-8切割后的GSDMC在TNF-α的作用下可诱导乳腺癌细胞发生焦亡。临床试验也证明了,PD-L1抑制剂与化疗或放疗结合可诱导肿瘤细胞发生焦亡,从而提高患者的生存率[19]。除了肿瘤细胞焦亡,免疫细胞焦亡同样可以触发强烈的免疫反应。Hage等[20]以索拉菲尼药物干预肝细胞癌小鼠模型,证明索拉菲尼可上调IL-1β、IL-18等促炎细胞因子的表达增强caspase-1活性,诱导巨噬细胞焦亡,并且触发了巨噬细胞同NK细胞的相互作用,使癌细胞死亡数量增加4倍。而NK细胞和细胞毒性T淋巴细胞(cytotoxic T lymphocytes,CTLs)所属的细胞毒性淋巴细胞是抗肿瘤免疫治疗中不可缺少的细胞执行者。NK细胞和CTLs细胞释放出的颗粒酶A可以在穿孔素靶向细胞的作用下裂解GSDMB,促进靶细胞焦亡,而颗粒酶B对肿瘤靶细胞的焦亡作用则是通过直接裂解D270位点的GSDME和间接激活相同位点的caspase-3诱导的。
3 中医药靶向细胞焦亡途径在抗肿瘤中的应用
3.1 中医理论下的细胞焦亡作为中医理论的重要组成部分及哲学基础,阴阳学说与现代生物医学中内环境的动态平衡及生物体内对立制约的调控息息相关,包括细胞生命活动的各个环节[21]。《素问·阴阳应象大论》曰:“阴阳者,天地之道也,万物之纲纪,变化之父母,生杀之本始,神明之府也。”阴阳被认为是万物变化的起源和生长毁灭的根本,阴与阳既对立又依存,二者互根互用,相互转化。阴阳属性体现在细胞层面上即为细胞的生长与死亡。《道德经》云:“万物负阴而抱阳,冲气以为和。”又有《素问·生气通天论》言:“阴平阳秘,精神乃治。”阴阳消长处于动态平衡时,机体各环节方能有序运转。细胞焦亡相较于细胞增殖,虽属“阴”,但其本身亦处于一种阴阳动态平衡之中,焦亡不足或过度失控则属于阴阳失衡。《素问·阴阳应象大论》言:“阳胜则阴病,阴胜则阳病。”在肿瘤疾病过程中,焦亡不足会加速癌细胞增殖,焦亡过度则会导致炎症物质释放过多,引发炎性肿瘤微环境的形成并导致新的炎症性疾病产生。因此,调控细胞焦亡要遵循“补其不足,损其有余”的原则,使细胞焦亡维持在适度的范围内,并恢复到阴阳平衡状态。
3.2 中医药靶向细胞焦亡途径的抗肿瘤作用尽管有些化疗药物会诱导癌细胞焦亡从而抑制肿瘤发展,但也有研究显示,化疗期间GSDME介导的焦亡会产生不良反应,原因可能在于GSDMs除了在肿瘤组织细胞中高表达外,也在正常组织细胞中表达,即在介导癌细胞焦亡的同时,也会导致正常组织细胞发生焦亡[9,22]。中药具有多成分、多靶点以及不良反应少的优点,随着对其调控细胞焦亡研究的深入,许多中药单体及复方通过靶向细胞焦亡途径抗肿瘤均发挥着自身优势。
3.2.1 中药单体
3.2.2.1 酮类化合物木犀草素作为天然类黄酮化合物,在高脂血症、哮喘等多种疾病中具有抗炎活性。Yu等[23]在木犀草素抗非小细胞肺癌(Non-small cell lung cancer,NSCLC)的机制研究中发现,木犀草素不仅可抑制NSCLC细胞中caspase-1的激活和IL-1β的成熟,且可以抑制NSCLC细胞中高表达的黑素瘤缺乏因子2(absent in melanoma 2,AIM2)炎症小体的激活,以抑制其上皮-间充质转变,从而发挥木犀草素的抗肿瘤作用。银杏叶提取物是包含银杏黄酮类化合物与萜内酯类化合物成分的物质,李军等[24]将不同浓度的银杏叶提取物作用于乳腺癌细胞MCF-7,在银杏叶提取物的干预下,MCF-7中GSDMD、caspase-1、cleaved caspase-1蛋白表达上调,且细胞焦亡率呈剂量依赖性提高,而Calpain-1、NLRP3蛋白以及Th17、Treg蛋白下调水平也随着药物浓度的增加而增加,提示银杏叶提取物可能通过靶向细胞焦亡途径及调控Th17/Treg水平延缓了乳腺癌的生长速度。高良姜素是从传统中药中提取的天然黄酮类化合物,孔杨[25]研究发现,在利用3-MA阻断自噬后,高良姜素诱导的原发性胶质母细胞瘤细胞焦亡标记GSDME的N端蛋白表达量增加,且在小鼠模型中高良姜素联合自噬抑制剂组ki67蛋白的阳性表达率最低,提示高良姜素可通过诱导GSDME介导的细胞焦亡增强抗肿瘤作用。远志酮是中药远志的酮类提取物,Chen等[26]研究发现,经远志酮干预的肝细胞癌组织及细胞系中的上游分子NLRP3、caspase-1、成熟的IL-1和IL-18蛋白表达量均显著提高,同时,ki67的表达呈剂量依赖性抑制,表明远志酮能够通过激活caspase-1介导的焦亡通路抑制肿瘤生长。水飞蓟宾提取自水飞蓟,是一种广泛用于降酶保肝的黄酮类抗氧化剂,在三阴性乳腺癌细胞MDA-MB-231中干扰线粒体生物发生、减少活性氧产生的同时,抑制NLRP3炎症小体的激活及caspase-1和IL-β蛋白的转录翻译,从而抑制其侵袭和迁移[27]。
3.2.2.2 萜类化合物熊果酸是夏枯草、山茱萸、白花蛇舌草等中药中具有抗炎、抗氧化作用的五环三萜类化合物。杨晨等[28]为明确其抗肿瘤机制,以不同浓度的熊果酸溶液作用于人肝癌SMMC-7721细胞,并观察其焦亡形态,结果显示,SMMC-7721的增殖率及集落数随着药物浓度的增加而降低,表明熊果酸上调了GSDMD-N、caspase-1、NLRP3的蛋白表达,且在30 μmol·L-1的药物浓度组别中,细胞出现了肿胀破裂、内容物释放等焦亡形态。Yuan等[29]研究显示,葫芦素B作为一种提取自天然中药的四环三萜类化合物,可以通过结合 toll样受体4(TLR4)激活NLRP3从而裂解GSDMD,诱导焦亡,亦能通过诱导A549细胞的线粒体障碍而引发焦亡。此外,在非小细胞肺癌小鼠模型中,葫芦素B组的肺肿瘤生长受到抑制,且无剂量依赖性。白桦脂酸是中药桑白皮中的五环三萜类化合物成分,研究表明在白桦脂酸单独或联合顺铂干预食管癌细胞TE-11建立的裸鼠异种移植模型的体内实验中,白桦脂酸联合顺铂组的Caspase-1阳性表达率与白桦脂酸组比较显著升高,而ki67阳性表达率明显降低,提示白桦脂酸通过细胞焦亡方式对顺铂治疗食管癌的效力具有增敏作用[30]。雷公藤甲素是中药雷公藤的主要活性成分之一,属于二萜类化合物。Cai等[31]通过体内外实验证明,雷公藤甲素可以下调头颈癌HK1、C666-1细胞的c-Myc/HK-Ⅱ 轴的表达水平及有氧糖酵解,并以沉默的HK-Ⅱ 激活caspase-3和GSDME的切割,诱导由GSDME介导的头颈部癌细胞的焦亡。黄独素B(Diosbulbin B,DB)是提取自黄药子的富含二萜内酯类成分的化合物,Li等[32]以低剂量黄独素B处理胃癌细胞,可下调PD-L1的表达,从而激活NLRP3炎症小体,促使顺铂治疗后的CR-GC细胞引发焦亡,提高顺铂治疗的敏感性。
3.2.2.3 生物碱类化合物来源于中药苦参的活性成分氧化苦参碱在多种肿瘤中发挥着抑制作用,张景正等[33]在氧化苦参碱诱导结肠癌HCT116细胞死亡的分子机制研究中发现,氧化苦参碱可以下调Wnt/β-catenin信号通路,促进细胞凋亡,而且在不同药物浓度下,关键蛋白GSDME的表达均有显著增高,cleaved caspase-3水平上调,表明氧化苦参碱在促进癌细胞死亡从而抗肿瘤方面可能较有潜力。Yao等[34]研究黄连的主要生物碱成分小檗碱对NLRP3炎性小体通路的抑制作用,以小檗碱对三阴性乳腺癌细胞进行干预,结果显示小檗碱不仅可以抑制MDA-MB-231活性及迁移能力,亦可下调NLRP3炎症小体级联中NLRP3、pro-caspase-1、ASC、IL-18、IL-1β蛋白的表达;同时,在小檗碱的作用下,MDA-MB-231细胞中P2X7的表达降低,表明小檗碱可通过抑制细胞因子的成熟和分泌改变肿瘤微环境,并抑制p2x7介导的NLRP3炎症小体激活。
3.2.2.4 苷类化合物重楼皂苷H是存在于中药重楼中的偏诺皂苷元型甾体皂苷,孙光强等[35]以重楼皂苷H干预耐替莫唑胺胶质瘤SHG44R细胞株,结果显示重楼皂苷H会诱导SHG44R细胞出现细胞膜受损及内容物释放等形态的焦亡改变,而且重楼皂苷H组的caspase-1、caspase-11、GSDMD、IL-1β、IL-18和NLRP3蛋白表达水平显著升高。芍药苷作为白芍提取物,具有抗炎、抗氧化及抗肿瘤药理活性,Wang等[36]研究表明,芍药苷通过泛素-蛋白酶体依赖途径对胶质母细胞瘤U87和U251细胞中TLR4具有下调作用,且NLRP3、caspase-1和pro-IL-1β等下游效应蛋白的表达水平降低,因此,芍药苷对胶质母细胞瘤的抑制作用是通过TLR4的分解及炎性因子释放的减少实现的。薯蓣皂苷是提取自山药的一类天然甾体皂苷类化合物,Ding等[37]在体内外实验研究中将薯蓣皂苷用于干预骨肉瘤细胞及MNNG/HOS细胞建立的骨肉瘤裸鼠模型,研究表明薯蓣皂苷可抑制细胞增殖及瘤体增长,并能使细胞出现特征性的焦亡形态,同时,GSDME-N的表达上调,且随着剂量的增加而增加;使用caspase-3特异性抑制剂Z-DEVD-FMK进一步干预后,GSDME-N与c-caspase-3的表达量均明显下调,证明薯蓣皂苷可通过靶向caspase-3-GSDME-N轴诱导骨肉瘤细胞焦亡,以发挥其抗肿瘤活性。提取自中药麦冬、麦冬皂苷B是苷类化合物的一种,程子昱的研究[38]通过电镜证明了麦冬皂苷B对3种非小细胞癌系细胞有不同程度的焦亡形态的改变,在其作用下,耐药细胞株A549/DDP中Cox2蛋白的表达以及线粒体膜电位显著降低,乳酸脱氢酶(lactate dehydrogenase,LHD)释放率更为显著,且焦亡通路中caspase-1、IL-1β、GSDMD等基因的表达均上调,因此麦冬皂苷B可通过激活经典焦亡通路以促进顺铂耐药株发生焦亡,逆转其耐药性。
3.2.2.5 多酚类化合物虎杖苷是从虎杖根茎中分离的活性成分,为白藜芦醇苷的多酚类化合物,Zou等[39]研究发现,虎杖苷在抑制非小细胞癌A549和H1299细胞增殖与迁移的同时,还可以抑制NLRP3、ASC和pro-caspase-1等的表达水平。此外,研究表明NF-κB与NLRP3炎性小体呈正相关关系,虎杖苷抗肿瘤可能是通过NF-κB信号通路抑制NLRP3的激活。姜黄素是一种分离自姜黄块茎中、不溶于水且具有抗癌活性的多酚类化合物,Liang等[40]用不同浓度的姜黄素干预肝癌HepG2细胞以研究其抗肿瘤机制,结果显示姜黄素处理后的HepG2细胞活性氧水平提高,细胞在出现肿胀、膜溶解等形态学改变的同时释放大量LHD,且随着姜黄素浓度增加,全长GSDME蛋白表达下调水平递增,GSDME-N上调水平递增,提示姜黄素能通过诱导焦亡促进癌细胞死亡。中药单体基于细胞焦亡途径抗肿瘤作用机制见表1。
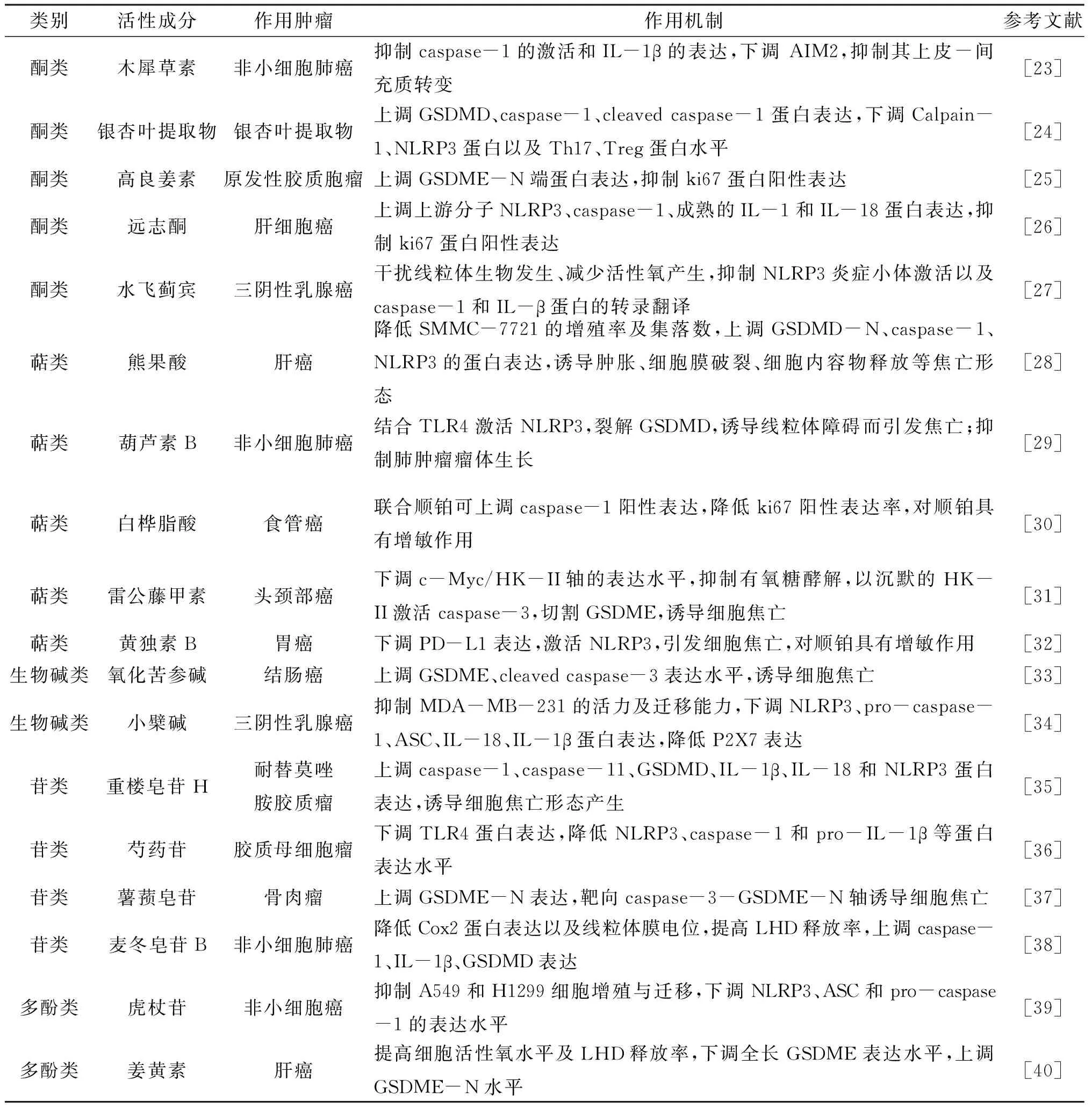
表1 中药单体基于细胞焦亡途径抗肿瘤的作用机制
3.2.2 中药复方西黄丸是源于《外科证治全生集》的抗乳腺癌经典方剂,临床广泛应用于肿瘤的治疗中。郭垠梅[41]研究表明,西黄丸干预肝癌SMMC-7721细胞后可以上调剪切体caspase-3、NLRP3、GSDME以及剪切体GSDME等蛋白的表达,而加入caspase-3抑制剂后,这些蛋白的表达水平降低,提示西黄丸能通过诱导细胞焦亡抗肝癌。健脾消癌方是蒋益兰教授根据临床经验及结肠癌“虚瘀毒”的病机特点自拟的方子,卢林竹[42]用SD大鼠制备健脾消癌方含药血清并将其作用于人结肠癌HCT116,观察细胞形态并检测各炎症因子及关键蛋白的表达,结果显示含药血清组细胞出现焦亡现象,且IL-1β、IL-18炎症因子的表达与正常组比较显著上升,各浓度含药血清组的caspase-3及全长GSDME的蛋白表达明显降低,而10%、20%药物浓度下 GSDMD蛋白表达下调,说明健脾消癌方对肝癌细胞焦亡的诱导机制可能与caspase-3/GSDME通路有关。固脾消积饮是由四君子汤加减而来的潘敏求教授的经验方,常被用于治疗原发性肝癌。翦慧颖[43]的研究显示,以固脾消积饮含药血浆干预HepG2.2.15细胞后,caspase-1抑制剂 VX-765 联合含药血浆组的乳酸脱氢酶及炎症因子IL-18、IL-1β的释放量均明显降低,而含药血浆组的caspase-1、GSDMD-N蛋白表达量与正常组比较均上调,固脾消积饮对肝癌细胞焦亡的诱导作用是通过激活caspase-1/GSDMD经典通路来实现的。
4 小结与展望
细胞焦亡途径已然成为肿瘤治疗的一个潜力靶点。究其机制,一方面,通过依赖caspase-1的经典焦亡途径和依赖caspase-4、caspase-5、caspase-11的非经典焦亡途径介导炎症小体的激活和炎症因子的释放,可促进癌细胞焦亡的发生;另一方面,TNF-α或肿瘤化疗药物均可通过诱导GSDME的裂解而完成细胞焦亡。随着对细胞焦亡认识的深入以及细胞焦亡与抗肿瘤相关性的探索,通过诱导细胞焦亡以促进化疗药物、免疫治疗等疗法发挥更好地抑制肿瘤增殖及瘤体生长效用的机制研究愈加深入,但“过度杀伤”正常细胞及其不良反应亦不容忽略。中医阴阳平衡理论与细胞焦亡的有序适度控制具有重要的指导意义,而中医药凭借其多靶点、多途径的特点对肿瘤化疗及免疫治疗增敏、减少耐药性的优势逐渐显现。但目前中医药对细胞焦亡途径的调控依然存在不足:首先,中药成分靶向细胞焦亡途径的研究尚处在基础阶段,且以单味药、单成分为主,其靶向机制和上下游靶点仍需进一步明确。其次,中药复方调控该途径的研究相对较少,因成分复杂且可能涉及多条信号通路,发挥细胞焦亡作用的主要成分并不明确。因此,结合网络药理学及分子对接等生物信息技术或基因敲除等方式对中药各成分影响细胞焦亡的机制进行多维度、多靶位以及多因子基础分析,并在中医平衡理论以及“损其有余,补其不足”治疗原则的指导下引导适度可控的焦亡,仍是中医药以及中西医结合调控细胞焦亡途径面临的挑战。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
医学综述(2022年7期)2022-04-19
数学物理学报(2021年4期)2021-08-30
昆明医科大学学报(2020年12期)2021-01-26
新世纪智能(数学备考)(2020年10期)2021-01-04
世界科学技术-中医药现代化(2020年2期)2020-07-25
中学生数理化·七年级数学人教版(2019年10期)2019-11-25
中学生数理化·高一版(2018年9期)2018-10-09
湖南教育·C版(2018年3期)2018-06-05
中成药(2017年12期)2018-01-19
