
梨酵母双杂交cDNA文库的构建与鉴定
2023-06-15 10:28:50易善萍聂显双李谢雨程寅胜
中国南方果树 2023年3期
龚 宏,易善萍,聂显双,刘 政,李谢雨,杨 立,程寅胜,张 义,伍 涛
(1 长江大学园艺园林学院,湖北荆州,434025;2 湖北省农业科学院果树茶叶研究所,武汉,430064;3 华中农业大学园艺林学学院,武汉,430070)
蛋白质是生物重要组成部分和功能单位,深入探究蛋白互作机制将为了解蛋白质的功能提供重要信息[1]。酵母双杂交技术是研究蛋白互作的有效分子生物学方法,在检测蛋白之间互作和筛选未知互作蛋白方面得到了广泛的应用[2-3]。例如,前人构建了海岛棉纤维均一化酵母cDNA文库,筛选到了4个可以与GbTCP5互作的蛋白[4]。郭振华等构建了桃酵母双杂交cDNA文库,筛选出了26个与生长素响应因子PpARF4互作的蛋白[5]。程寅胜等利用砀山酥梨cDNA文库筛选获得了13个与梨糖转运相关蛋白PbTMT4潜在互作的蛋白[6]。这些研究促进了相关基因功能机制的解析,而构建高质量酵母双杂交文库是开展相关研究的前提。
梨是我国重要的经济水果,提高光合作用效率以增加干物质积累是提升梨产量和品质的有效技术途径[7-8]。本课题组前期利用生理测定和转录组联合分析的方法,挖掘出了与梨树光合作用密切相关的关键基因PpPRR5,并通过瞬时遗传转化的方法验证其具有光合作用的功能,然而其调制机制尚不明晰[9-10]。PpPRR5同源基因AtPRR5被报道是拟南芥伪应答调节蛋白家族成员之一[11-12],PR(Pseudo-Receiver)结构域和CCT(CONSTANS,CONSTANS-LIKE和TOC1)结构域分别位于该家族N端和C端[11-13]。AtPRR5能与其他家族成员协同调控拟南芥生物钟节律[13]。在调控机制方面,AtPRR5的转录水平和翻译水平并不完全同步;在光形态建成早期阶段,AtZTL蛋白通过结合AtPRR5的PR结构域介导其泛素化降解;AtZTL的LOV结构域可以被蓝光诱导发生构象变化,使AtZTL与AtGI互作利于增强其蛋白稳定性,阻止AtZTL对AtTOC1、AtPRR5的泛素化降解[14]。此外,AtPRR蛋白(AtPRR9、AtPRR7和AtPRR5)通过与AtBBX19发生蛋白水平上的互作,进而调控AtCCA1、AtLHY和AtRVE8启动子[15]。通过酵母双杂交挖掘与PpPRR5互作的蛋白,也将有助于进一步解析调控靶基因的分子机制,阐明其调控光合作用的分子机理。因此,本研究以“圆黄”梨(PyruspyrifoliaNakai cv. Wonhwang)叶片和果肉为材料,构建酵母双杂交cDNA文库,旨在为后续利用酵母双杂交技术筛选出关键基因的互作蛋白,为深入明晰调控网络奠定基础。
1 材料与方法
1.1 试材
1.1.1 植物材料 本研究材料以湖北省农业科学院果树茶叶研究所展示园中的“圆黄”梨树(6年生成年树)为研究试材,采集成熟叶片和果肉组织(盛花后约140天)样品后液氮速冻,置于-80 ℃冰箱中保存。
1.1.2 试剂与药品 cDNA文库构建试剂盒(货号:630490)、Taq DNA Polymerase(货号:639141)和T4 DNA Ligase(货号:2011A)购自于Clontech公司。Tris(货号:T8060)购自于Solarbio公司;凝胶回收试剂盒(货号:9762)购自于Fermentas公司。DEPC(货号:ST036)购自于碧云天公司。RNase H(货号:2158-1)购自于TAKARA公司;DMSO(货号:D8418)购自于Sigma公司,异丙醇等其他试剂均为分析纯,购自于国药集团化学试剂有限公司。
1.2 试验方法
1.2.1 RNA抽提和双链cDNA的合成 取“圆黄”梨叶片和果肉组织研磨成粉末状后等质量混合,利用Trizol法提取总RNA。采用琼脂糖凝胶电泳和NanoDorp 2000检测所提取总RNA,并检测其质量和浓度。对总RNA进行纯化处理,采用SMART技术反转录合成cDNA。采用cDNA文库构建试剂盒中提供的5’-和3’-PCR primer,通过LD-PCR(Long-distance-PCR)方法合成双链cDNA,使用试剂盒中的CHROMA SPIN Columns + TE-400柱对产物进行纯化,纯化产物进行琼脂糖凝胶电泳检测,纯化cDNA的长度范围为500~2 000 bp,具体步骤参照试剂盒说明书的方法进行。
1.2.2 梨cDNA文库的构建 将pGADT7载体线性化,并与梨dscDNA进行同源同组反应。取连接产物加入大肠杆菌感受态细胞DH5α,通过电转化后涂布于LB(氨苄抗性)平板上,37 ℃过夜培养后收集菌体,加入适量甘油后置于-80 ℃冰箱中保存。
1.2.3 梨cDNA文库质量鉴定 取10 μL文库构建菌液,按102、104、106倍逐级稀释,取100 μL涂布氨苄抗性的LB固体平板,37 ℃倒置培养16 h,统计菌落数并计算文库滴度。
文库滴度cfu/mL=克隆数目/铺板体积mL×稀释倍数。
随机挑取转化后的平板上的单克隆20个,液体摇菌培养至对数期,提取质粒并进行PCR鉴定,鉴定文库插入片段的大小和阳性率。
2 结果与分析
2.1 RNA和双链cDNA质量检测
梨果肉和叶片组织等质量混合后提取总RNA。琼脂糖凝胶电泳检测结果如图1所示,RNA主带清晰,完整性好,无降解。其中A260/A280=1.86,浓度为0.122 8 μg/μL,表明所提取的RNA含量和纯度满足酵母双杂交cDNA文库的建库要求。
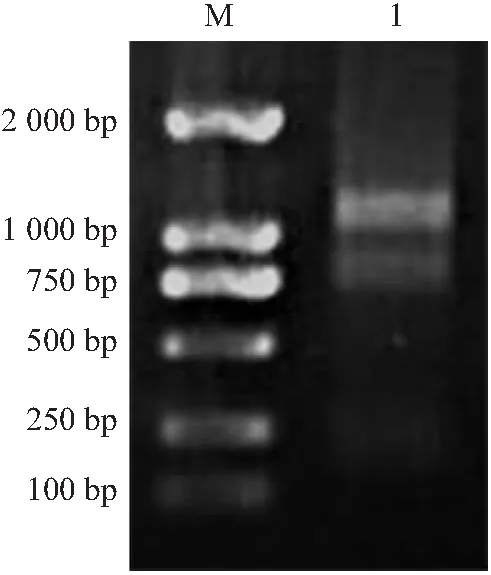
注:M:DL2000 DNA ladder Marker;1:总RNA
双链cDNA琼脂糖凝胶电泳检测如图2所示,未纯化的双链cDNA主要集中在500~2 500 bp。经纯化柱纯化后的双链cDNA,主要分布集中在600~1 500 bp。说明纯化后的双链cDNA较为集中,分布均匀完整,多态性较好(见图3)。
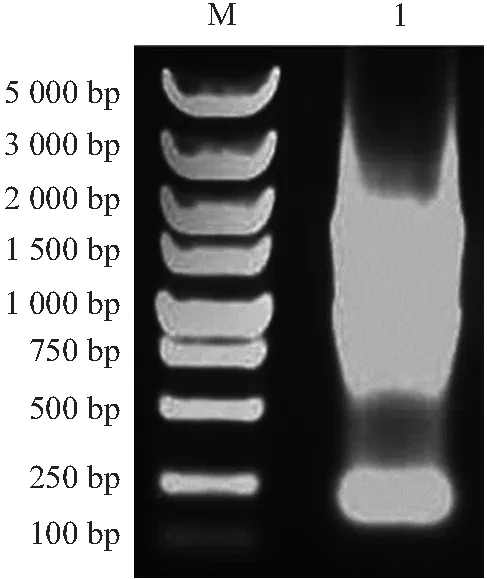
注:M:DL2000 DNA ladder Marker;1:LD-PCR产物
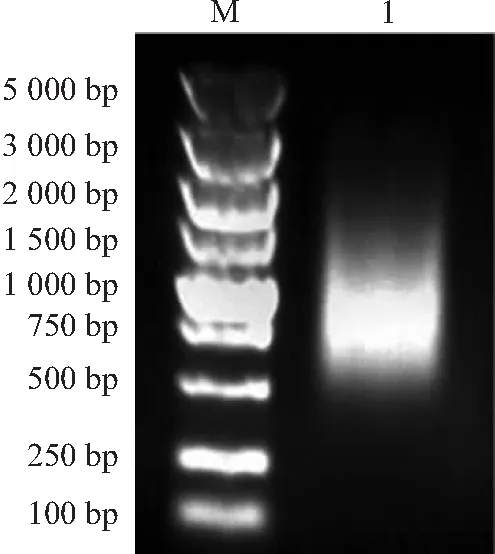
注:M:DL2000 DNA ladder Marker;1:纯化后的cDNA
2.2 cDNA文库滴度测定及阳性率检测
梨cDNA文库滴度鉴定结果如图4所示,根据文库滴度计算公式可得文库滴度为1.76×108cfu/mL≥1×107cfu/mL,说明所构建的梨cDNA文库所包含的基因信息较完整,文库质量较好,覆盖度较广。
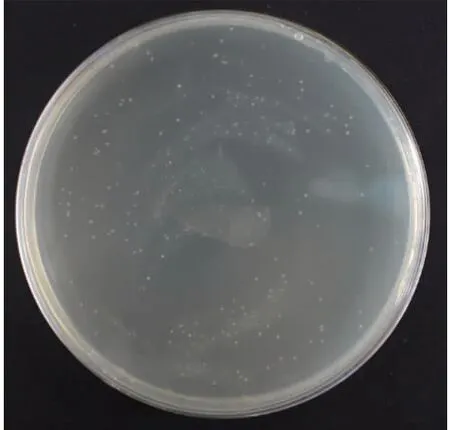
图4 梨cDNA文库滴度鉴定
为检测文库重组序列的完整性,对在平板上随机挑选20个单克隆进行菌落PCR鉴定。结果显示,扩增条带大小主要分布在600~1 500 bp,表明所构建的cDNA文库空载率低,cDNA重组序列的完整性较好(见图5)。

注:M:DL2000 DNA ladder Marker;1-20:PCR产物;+:阳性对照(以pGADT7-T为模板PCR);-:阴性对照(以水为模板PCR)。
3 讨论
酵母双杂交技术已被广泛应用于研究蛋白质之间的相互作用[16]。例如,通过构建草莓不同发育时期果实的酵母双杂交cDNA文库,筛选出13个与促分裂原活化蛋白激酶FvM4K1互作的蛋白,进一步明晰了FvM4K1参与调控草莓果实发育的分子机理[17]。利用已构建的花生叶片的cDNA文库,筛选出5个与AhCaM互作的蛋白,初步提出了AhCaM提高抗逆性的可能分子机制[18]。张竹君等[19]成功构建了干旱胁迫诱导的高质量月季cDNA文库,并利用酵母双杂交技术筛选到了7个与RhRD22相互作用的蛋白。本研究以成熟时期“圆黄”梨的叶片和果肉组织为试材,构建梨的酵母双杂cDNA文库。高纯度、完整性好的mRNA是构建cDNA文库的关键,并且直接影响着文库的质量[20]。本研究提取的梨组织混合RNA样本经琼脂糖凝胶电泳检测,RNA纯度较高,完整性较好,符合建库的要求(见图1)。cDNA文库质量主要依赖文库的容量、文库的重组率和插入片段的大小等方面的评判[21]。一个良好的cDNA文库要求文库滴度至少为1×107cfu/mL[22]。若是文库的库容量过低,则会使文库所包含的基因信息不全面,从而会降低筛选到低丰度基因的概率[23]。本研究通过同源重组的方法构建的梨酵母双杂交cDNA文库,其文库滴度为1.76×108cfu/mL,说明文库质量较好(见图4)。文库中的重组cDNA插入片段大小和基因重组率代表重组序列的完整性。植物的cDNA长度一般在500~3 000 bp之间,文库插入片段过小或过大均对文库质量产生影响[24]。本研究随机检测构建的cDNA文库,其基因重组率为100%,插入片段主要分布在600~1 500 bp,达到了构建文库的标准,说明文库构建质量较好,能为后续利用酵母双杂交技术筛选关键基因的互作蛋白奠定基础。
猜你喜欢
湖南农业大学学报(自然科学版)(2022年2期)2022-05-11 05:50:28
太原理工大学学报(2021年6期)2021-11-25 13:33:20
作物学报(2021年11期)2021-08-31 05:37:08
海外星云 (2021年21期)2021-01-19 14:17:31
山东畜牧兽医(2021年6期)2021-01-11 10:31:57
中国海洋大学学报(自然科学版)(2019年7期)2019-05-21 07:26:06
中国海洋大学学报(自然科学版)(2019年7期)2019-01-04 16:33:12
广州化工(2016年11期)2016-09-02 00:42:59
当代经济(2016年26期)2016-06-15 20:27:14
河南科技(2014年12期)2014-02-27 14:10:34
