
HBV慢性感染诱导的染色体超倍化及靶向治疗策略
2023-06-01 12:41施旭佳尧晨光李涵泺魏艳红胡康洪
世界华人消化杂志 2023年8期
施旭佳,尧晨光,李涵泺,魏艳红,胡康洪
施旭佳,尧晨光,李涵泺,魏艳红,胡康洪,湖北工业大学中德生物医学中心;教育部及国家外专局细胞调控与分子药物学科“111”创新引智基地;湖北省/教育部工业发酵省部共建协同创新中心 湖北省武汉市 430068
0 引言
乙型肝炎病毒(hepatitis B virus,HBV)是嗜肝病毒科的DNA病毒成员之一,其特异性感染肝脏,从而诱发慢性肝炎(chronic hepatitis B,CHB)、肝硬化及肝细胞癌(hepatocellular carcinoma,HCC),全球约50%的HCC患者感染HBV[1].
既往研究表明,HBV可通过基因整合或突变引起宿主细胞有丝分裂异常,染色体数目成倍或非成倍增加,这种现象被称为染色体超倍化.染色体数目成倍增加的现象称为多倍化,而非成倍增加的现象称为非整倍化.HBV诱导的超倍化、染色体不稳定性(chromosomal instability,CIN)、抑制免疫微环境及端粒破坏等途径是HCC形成的重要因素(图1)[2-4].高度CIN会使细胞死亡,然而,在多种肿瘤细胞中,CIN导致的突变累积、拷贝数变异等活动使细胞适应不利环境,驱动癌症发生[5,6]和转移[7],并导致肿瘤患者预后不良与化疗抵抗[8,9].此外,近乎一半的CIN可由细胞分裂过程中染色体出现的超倍化(即多倍化和/或非整倍化)通过染色体错误分离或畸变等途径诱导产生[10].

图1 CHB导致肝癌简易机制图.HBV感染可直接通过病毒蛋白抑制免疫微环境、破坏端粒和超倍化等途径促进HCC的发生发展,也可通过早期HBV DNA整合宿主基因组和或诱导CIN等途径促进HCC的进展,同时,CIN可作为超倍化诱因,而超倍化也可诱导CIN.HBV: 乙型肝炎病毒;CHB: 慢性乙型肝炎病毒感染;CIN: 染色体不稳定性;HCC: 肝细胞癌.
一个完整的细胞周期分为G0、G1、S、G2、M期,其中G1、S和G2期又称为有丝分裂间期,主要为有丝分裂M期储备充足的核酸和蛋白质来复制基因组.当胞质分裂阻滞或M期被截断时,细胞会停滞在间期或M期从而产生多倍体细胞[11,12].有丝分裂过程中还会发生纺锤体装配检查点缺陷、内聚缺陷、Merotelic附着,中心体扩增等错误产生超倍化、DNA损伤与CIN等事件,均有助于肿瘤的发生和发展[13].
在正常肝细胞中,多倍体细胞如四倍体或八倍体约占25%,这样的多倍体化与终末分化和衰老相关[14].多倍体细胞的有丝分裂有两种形式,一种是多极分裂产生多个非整倍体细胞,另一种是双极分裂产生两个多倍体细胞,产生的多倍体细胞还会进一步分裂产生多倍体细胞或非整倍体细胞.正常肝细胞的多倍体化是一种复杂的、有序的生理过程,受到多种因素的调控,包括细胞周期调控因子、细胞信号通路、肿瘤抑制基因等.在这种调控下,细胞可以进行有序的多倍化,而不会导致HCC[13-18].部分肝切除术、黄曲霉素、代谢过载或辐射等也能诱导肝细胞多倍体比例的上升[14].在HBV与HCV感染相关肝炎、肝纤维化、肝硬化及肝癌中也发现肝细胞的倍性改变,表现为多倍体化或非整倍体化,受病毒对细胞的影响,这些多倍体化或非整倍体化难以经细胞调控而修复从而发生致癌级联反应[19,20].
因此,以上证据表明,HBV与超倍化及CIN具有高度相关性,超倍化可能作为HBV诱导HCC的一条额外途径.
1 慢性HBV感染诱导肝细胞染色体超倍化及致癌转化的途径
1.1 HBV上调Polo样激酶1 Polo样激酶1(polo-like kinase 1,PLK1)是有丝分裂和非有丝分裂过程中的重要调节因子之一,在多种癌症中高表达,其可调节包括中心体成熟、姐妹染色单体分离、纺锤体组装、有丝分裂退出、DNA复制、DNA损伤反应、G2 DNA损伤检查点恢复、染色体动力学和微管动力学等多个环节[21,22],因此其功能障碍可能促进癌症发生发展并与预后不良相关[22-24].

表1 迄今已报道的针对超倍化相关的部分小分子抑制剂及其靶点
HBV表达的HBx蛋白有多种生物学功能,包括参与解除对细胞周期检查点的调控、激活/抑制细胞凋亡、提高糖异生水平、激活主要代谢信号通路、表观遗传调控等[25,26].在HBx转基因小鼠中,HBx能通过p38和ERK1/2途径上调PLK1(图2),诱导细胞周期检查点破坏,导致肝细胞多倍化、双核性减少、DNA损伤累及和炎症,从而促进致癌转化(同时HCC祖细胞标记物Ly6D表达上升)[27].另外,值得注意的是,有研究显示[28],在表达HBx的4pX-1细胞系中,HBx介导的多倍化可能与致癌转化无直接关系,在此过程中,PLK1诱导的DNA损伤累及才是致癌转化的关键.免疫共沉淀分析其可能是通过下调ZNF198和SUZ12,从而增强HBV复制、DNA损伤及调节p53活性,促进转化[29].另一项研究也提出[30],70%的HBV慢性感染患者体内LHBS阳性肝细胞的细胞核大小及超倍化水平均显著高于LHBS阴性肝细胞,可能是由LHBS反式激活PLK1,从而破坏G2/M检查点,诱导胞质分裂失败,但LHBS具体如何上调PLK1的机制尚不明晰.二倍体细胞胞质分裂失败应产生四倍体/多倍体,但该研究显示大多数LHBS阳性肝细胞染色体数为51条-70条(非整倍体),可能的解释为LHBS阳性细胞的多倍体化伴随着异常的多极分裂,这最终导致超倍体产生[30].
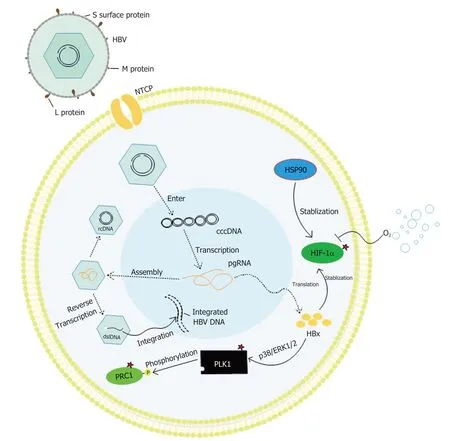
图2 HBV感染导致超倍化机制示意图.HBV病毒颗粒通过其肝脏特异性受体NTCP进入细胞后,其基因组可以转化为cccDNA从而转录pgRNA作为翻译模板,得到包括HBx在内的病毒蛋白质,进而上调PLK1、PRC1及CCNE1,诱导超倍化产生;病毒基因组也可整合入宿主基因组中,通过前述各途径诱导超倍化,超倍化包括非整倍体化和多倍体化.HSP90和HIF-1α可促进癌细胞中非整倍化介导的细胞耐药及存活.红色星号标记为直接诱导超倍化的蛋白靶标.HBV: 乙型肝炎病毒;NTCP: 钠牛磺胆酸共转运多肽;cccDNA: 乙肝病毒共价闭合环状DNA;rcDNA: 松弛环状DNA;dslDNA: 双链线性DNA;pgRNA: 前基因组RNA;HBx: 乙肝病毒X蛋白;PLK1: Polo样激酶1;PRC1: 蛋白调节因子1;CCNE1: 细胞周期蛋白E1;HSP90: 热激蛋白90;HIF-1α: 缺氧诱导因子-1α;ERK1/2: 细胞外调节蛋白激酶1/2.
PLK1可以调节有丝分裂后期胞质分裂蛋白调节因子(protein regulator of cytokinesis 1,PRC1)的激活(图2)[12],PRC1是与CIN相关性排名第二的基因[31],通过稳定纺锤体中间区结构来稳定促进胞质分裂,其过表达与敲低均可引起细胞分裂异常[12],进一步促进非整倍体、肿瘤发生和不良临床患者结果[32-37].由此可见,PLK1是HBV相关超倍化诱导致癌转化过程中的关键蛋白,但HBV到底是通过怎样的相互作用来控制PLK1还尚不清楚.
1.2 HBV DNA与宿主染色体的整合并上调CCNE1 作为一种DNA病毒,HBV可以通过其双链线性DNA(dsl DNA)将基因整合到宿主染色体(图2),在约80%-90%的HBV感染的HCC患者肝癌细胞中发现HBV DNA与宿主细胞染色体整合现象[38,39].HBV DNA整合到宿主基因组的位置存在随机性,但在HCC中,HBV DNA与宿主细胞基因整合位点主要富集在CpG岛(与DNA甲基化有关)、端粒及一些DNA易突变或重排的基因组区域[40].
细胞周期蛋白E1(cyclin E1)细胞周期的G1和S期发挥作用,促进细胞周期顺利完成S期,编码cyclin E1的CCNE1基因座被观察到在许多类型的癌细胞中扩增,而HBV基因能整合在CCNE1基因附近,并通过病毒本身的增强子使CCNE1过表达(约32倍),从而诱导肝细胞超倍化,选择性诱导肝脏细胞CIN和DNA双链断裂(DBS)诱导肝细胞癌变[41,42].此外,CCNE1还可通过过度激活PLK1等因子来破坏细胞分裂的动力学[41].
1.3 HBV的L和S蛋白具有促进多细胞融合的潜力 在一些病毒诱导作用下,宿主细胞会发生融合现象,从而产生多倍体细胞.HCV和HPV含有细胞融合诱导蛋白(HCV的E1、E2蛋白,HPV的E5蛋白)导致合胞体形成并诱导细胞多倍体化[43,44].值得一提的是,细胞融合过程如HPV介导的细胞融合包括细胞质膜与核膜的融合,质膜融合后通常会形成双核多倍体细胞,这些细胞大多会死亡,只有少部分细胞发生核膜融合变为单核多倍体细胞,这些细胞具有不稳定的基因组和基因表达,最终容易发展成为癌细胞[44].
有研究表明,HBV的L(preS1)和S蛋白是HBV能够发生膜融合从而进入细胞的关键因素[45].ERp57蛋白能促进HBV表面S蛋白交叉链二硫键构象变化,导致preS1中融合肽的暴露,激活HBV的膜融合能力[45].更重要的是,L、S蛋白在胞内和胞外均可介导细胞融合,且这种融合不依赖于与HBV感染受体牛磺胆酸钠共转运肽(NTCP)相互作用和胞吞作用[45].有研究表示[10],在HCC肝组织和慢性乙肝肝炎中有更高的ERp57的表达,且与不良预后有关.
2 靶向HBV相关超倍化药物研究进展
目前,肝癌化疗药物主要有索拉非尼、阿替利珠单抗、贝伐珠单抗等,但由于耐药、肿瘤异质性等问题的存在,这些化疗药物的治疗效果对部分患者仍不理想.对于CHB诱导的超倍化相关HCC,抑制HBV大表面蛋白(HBV large surface protein,LHBs)、HBx和宿主PLK1、PRC1的表达以及使低氧诱导因子-1α(hypoxia inducible factor-1,HIF-1α)失稳可能是治疗HCC的有效策略(表1).
PLK1和PRC1为胞质分裂关键调节因子,降低PLK1和PRC1的表达或抑制其活性可以抑制肿瘤的生长[46].PLK1在多种癌细胞中特异性高表达,目前,已开发出一些直接靶向PLK1的小分子抑制剂如TAK-960、BI-2536、GSK461364[47,48]、Volasertib[49]和Rigosertib[50].这些小分子抑制剂已经被证明可以单独或联合其他药物抑制多种肿瘤[49-51].此外,由于PLK1致中心体扩增的特点[6],癌细胞通过中心体聚集而诱导细胞发生非整倍化,因此一系列破坏细胞中心体异常聚集的药物也值得我们关注[13],如HSET/KifC1抑制剂CW069、AZ82和SR31527[52-54].
PRC1参与纺锤体中间区的形成及平衡着丝粒张力,这决定了其在细胞分裂、染色体不稳定性及肿瘤异质性中存在的重要作用[12].遗憾的是,目前仅有一些PRC1上游靶点的抑制剂,如CBXs,BMI-1和RING1B的抑制剂[55],尚没有直接靶向PRC1的药物.PRC1在多种癌症中异常表达并与预后相关,因此其还可作为预后相关生物标志物[56].
非整倍体细胞对毒性应激和代谢应激类化合物更敏感,因此这类化合物可应用于HBV相关非整倍体导致的HCC[13,57],如HSP90抑制剂17-AAG、格尔德霉素衍生物和XL888[58]或HSF1抑制剂,从而加剧非整倍体蛋白折叠障碍[59].抑制HSP90的药物同时还能诱导HIF-1α降解[60],抑制HIF-1α蛋白水平可以降低高倍体细胞[61]与肿瘤细胞非整倍体耐药性[62,63].非整倍体细胞还表现出神经鞘脂代谢失调,导致促凋亡脂质神经酰胺水平升高,使得增加神经酰胺水平的药物如神经酰胺类似物DLPDMP对非整倍体细胞的毒性大于二倍体细胞,促进非整倍体细胞凋亡[13,64].
清除HBV cccDNA是治疗HBV感染相关HCC的终极目标.鉴于cccDNA的致密超螺旋微染色体结构,其很难从患者体内被清除,现阶段的治疗策略主要为利用核苷(酸)类似物抑制HBV DNA复制或利用干扰素免疫抑制HBV DNA复制,但这两种策略清除cccDNA的周期中位数约34年,存在cccDNA整合至宿主基因组、病毒耐药或患者依从性差等问题.目前,一些直接或间接靶向cccDNA的方法正在开发,如基于CRISPR的基因编辑直接降解cccDNA,碱基编辑直接靶向cccDNA引发其甲基化、突变及降解[65],人工转录因子靶向HBV增强子Ⅰ抑制其转录和复制[66].
HBV基因组与宿主细胞染色体整合后,HBV转录可逃避免疫监视从而长期潜伏在宿主体内,为治疗HBV相关HCC带来了极大的挑战.HBV整合机制极具争议,六种可能的模型被提出,包括松弛环状DNA介导的单链缺口模型、直接重复序列介导的整合模型、“滚入”模型、部分序列同源模型、基于细胞酶介导的非同源末端连接和微同源性介导的末端连接[67].更加深入和清晰地解析cccDNA整合机制有助于开发未来针对HBV相关HCC的新疗法.
3 结论
流式细胞术和单细胞测序均证实正常肝组织中存在一定比例的多倍体肝细胞[68],HBV感染进一步导致肝细胞的多倍化或非整倍化,从而诱导广泛的染色体不稳定性和肿瘤的发生及转移.此外,一些研究表明在化学诱导的肝细胞向HCC转化过程中,细胞倍性减少,可能是由于二倍体细胞而非多倍体细胞对隐性突变或有毒物质的保护更少[7],而在HBV感染的肝细胞,倍性更倾向非整倍体变化.非整倍体化在土拨鼠肝炎病毒所致的土拨鼠肝癌细胞中也有发现[32].产生这种差异的可能解释是,HBV感染诱导肝细胞细胞周期或胞质分裂调节相关蛋白的失调[67],从而导致肝细胞染色体超倍化和肝癌的发生.
HBV可通过多种途径使PLK1及PRC1失调从而诱导超倍化.目前,它们的一些抑制剂已经进入开发阶段.此外,p53也在多种病毒介导的肿瘤转化中发挥重要的作用[69,70].p53的存在与激活可以防止细胞周期异常,进而控制细胞倍性和染色体不稳定性[71].尽管HBV感染初期导致的非整倍体可能并不影响p53功能,但在HBV相关多倍体肝癌细胞中p53失活,这提示了p53与HBV相关超倍化有着密切的联系[41,72].近来,两种p53功能恢复剂的出现使细胞检查功能的恢复成为可能,包括三氧化二砷和更为低毒的酒石酸锑钾[73,74].
HBV与超倍化及CIN之间的关联可能还不止如此,生物信息学分析发现与CIN相关性最强的TPX1基因[31]在HBV相关HCC中表达上调,其上调可导致多核性及多倍化[75],参与HCC发生发展,可以用作独立预后基因之一[76,77].
猜你喜欢
今日农业(2020年14期)2020-12-14
诊断学(理论与实践)(2020年6期)2020-12-09
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
广东医科大学学报(2020年6期)2020-02-06
中国妇幼健康研究(2019年12期)2019-12-31
中华肺部疾病杂志(电子版)(2018年1期)2018-03-12
临床与实验病理学杂志(2017年3期)2017-03-07
中国卫生标准管理(2015年4期)2016-01-14
实用皮肤病学杂志(2015年4期)2015-12-22
