
柑橘大实蝇产卵器的超微结构观察(双翅目:实蝇科)
2023-05-27 14:01:00周仁迪桂连友刘梅柯何章章华登科刘文茹姜振宇汤建涛孙康林
环境昆虫学报 2023年2期
周仁迪,桂连友*,刘梅柯*,何章章,杨 璇,华登科,刘文茹,姜振宇,汤建涛,孙康林
(1. 长江大学农学院,湖北省农林病虫害预警与调控工程技术研究中心,湖北荆州 434025;2. 湖北省农业科学院,农业质量标准与检测技术研究所,武汉 430064)
柑橘大实蝇Bactroceraminax(Enderlein)属于双翅目实蝇科,是柑橘类重要害虫之一(陈世骧和谢蕴贞,1955;罗杰等,2016;杨璇等,2022)。柑橘大实蝇雌虫产卵于柑橘幼果中,幼虫在果实内部蛀食,常使果实未熟先黄并提前脱落,使其完全失去食用价值,严重影响柑橘产量和品质(黄大树等,2007;王小蕾和张润杰,2009)。
雌性昆虫的外生殖器称为产卵器,在昆虫交配、产卵和排泄行为中发挥着关键作用(Norrbom and Kim, 1988)。产卵器的基本构造为管状构造,由三对产卵瓣构成,分别为第一产卵瓣(腹产卵瓣),第二产卵瓣(内产卵瓣)和第三产卵瓣(背产卵瓣)(Hünefeldetal., 2012)。昆虫的产卵器主要分为两大类:产卵器或真产卵器,起源于腹节附肢,保留了产卵器最基本的产卵瓣结构,并出现了相应的发展、特化或消失。如直翅目、半翅目和膜翅目等的产卵器;另一类是伪产卵器,其产卵瓣的结构己经退化或与腹部愈合,不具备由附肢特化的产卵瓣结构,只是腹部后端几个体节逐渐变细,可以伸长、收缩、套叠的管状结构。如鞘翅目叶甲科、双翅目实蝇科和鳞翅目姬蜂科的产卵器(张璐,2015)。
双翅目实蝇科昆虫的伪产卵器是由第7、8和9腹节演化而来(Drew, 1969; Dodson, 1978; Norrbom and Kim, 1988; 吴佳教等,2009)。第7腹节为产卵器基节,背腹扁平,背板和腹板在侧面愈合,1对气门位于腹外侧,腹板向前形成了一个三角形区域,其前部形成了一个薄的卵圆形膜(Drew, 1969; Stoffolano and Yin, 1987)。第8腹节为翻缩膜(eversible membrane),长而柔软呈管状(Hanna, 1938; Steyskal, 1986),具三角形骨化齿或鳞片(Hanna, 1938; Stone, 1942; Drew, 1969; Dodson, 1978; Stoffolano and Yin, 1987; Norrbom and Kim, 1988)。第9腹节为产卵针,由一块背片和两块腹片组成,背片高度骨化,一直延伸到尖端,腹片短于背片(Drew, 1969; Stoffolano and Yin, 1987)。两块腹片之间、背片与腹片之间均以柔性的角质层或柔性膜连接(Berube and Zacharuk, 1983; Stoffolano and Yin, 1987)。靠近产卵针的尖端的两侧有2个凹槽,每个凹槽内有3对化学感受器(Stoffolano and Yin, 1987)。在实蝇科昆虫中,产卵器的生殖孔和肛门开口合为一体,从而形成了泄殖腔结构(Dean, 1935; Stoffolano and Yin, 1987)。实蝇科昆虫的产卵器内部不仅有调节产卵器内部液体压力的泵和阀门,还有直肠、输卵管、气管和血淋巴(Dodson, 1978; Berube and Zacharuk, 1983)。除此之外,还存在肌肉组织,如内腹肌(Medioventral muscle)、侧腹肌(Lateroventral muscle)、背腹肌(Tergosternite muscle)、背肌(Dorsal muscle)(Ovchinnikova, 2008; Ovtshinnikova, 2010; 2012; Berube and Zacharuk, 1983)。产卵器收缩时,首先第9腹节(产卵针)折叠在第8腹节内(翻缩膜),然后再一起折叠在第7腹节内(基节),因此,形成了产卵针、翻缩膜和基节3个腹节的套叠(Drew, 1969; Berube and Zacharuk, 1983)。
柑橘大实蝇的产卵器在产卵过程中能够完成多种运动类型,如套叠、弯曲、伸长和收缩(李宗锴等,2014;桂连友等,2020)。在产卵过程中,用产卵器敲击柑橘果实表面,找寻合适产卵位点,确定位点后,基节留在果实外,翻缩膜伸展产生压力使产卵针垂直刺入果实,产卵针刺入合适深度后在橘果中产卵(尤克西等,2012;李宗锴等,2014)。基于光学显微镜,对柑橘大实蝇产卵器的形态结构研究较多,柑橘大实蝇产卵器基节为橙褐色,表面密被细毛,大致为漏斗状,背腹扁平,端部细长、渐尖;翻缩膜的中部有鳞片状纹;产卵针表面光滑无锯齿,末端尖锐且坚硬,利于雌虫刺穿果皮(Christenson and Foote, 1960; 李宗锴等,2014)。而基于扫描电镜,对柑橘大实蝇产卵器感器的观察结果尚未统一,张国娜(2012)仅清晰的观察到有毛形感受器和钟形感受器,宋傲群等(2015)仅清晰的观察到有毛形感受器和腔锥形感受器。
综上所述,有关柑橘大实蝇产卵器的超微形态结构研究较少,特别是对其产卵器的内部结构包括其套叠结构的描述未见报道。本研究基于光学显微镜、扫描电镜、石蜡切片和透射电镜观察,对柑橘大实蝇的产卵器形态结构进行观察描述,将为理解昆虫进化和多样性,以及分析昆虫的产卵、交配和代谢物排泄等行为机制提供新的例证。
1 材料和方法
1.1 供试虫源
2020年10月从湖北省宜昌市(30°37′N, 111°3′E)未采取防控措施的柑橘园采集虫果,带回长江大学昆虫生态学实验室,埋在河沙(厚度5~7 cm)中,幼虫化蛹后,分期分批收集蛹,转移到盛有河沙(厚度5~7 cm) 的塑料盆中(直径17 cm,高11 cm),保持土壤湿度20%左右。收集羽化后的成虫,置于养虫笼(35 cm×35 cm×35 cm)中饲养,饲喂的食物是酵母菌浸粉(纯味型,青岛海博生物技术有限公司)和蔗糖化学纯(北京康普汇维科技有限公司),并按1∶3的质量比混合(何章章等,2022)。室内环境条件温度为25±2℃、相对湿度为65%±5%,以所饲养的20日龄雌虫作为供试虫源。
1.2 研究方法
1.2.1基于光学显微镜对产卵器的解剖、观察和测量
产卵针收缩状态下的产卵器样本:在徕卡光学显微镜下,剪去其头部、胸部以及腹部1~5节,仅保留收缩状态下的产卵器样本;产卵针完全伸出产卵器样本:在徕卡光学显微镜(Leica M205A)下,首先剪去雌成虫的头部和胸部,然后用镊子挤压腹部,使产卵针完全伸出,待产卵针不收缩后,将腹部1~5节去除,获得产卵针完全伸出产卵器的样本;最后进行观察和测量(Drew, 1969),记录产卵器各组成部分的长度与宽度(各部分的1/2处)。共测试24头成虫。
1.2.2基于扫描电子显微镜对产卵器超微形态观察和拍照
第一步:固定。用2.5%戊二醛将上述产卵器固定。第二步:清洗和脱水。首先将样品放入磷酸缓冲盐溶液(PBS)中,然后用超声波清洗 10 s,最后用30%、50%、70%、90%和100%乙醇溶液依次梯度脱水。第三步:干燥。将产卵器置入冷冻干燥机(Christ-Alpha 1-2 LD)干燥。第四步:喷金。将产卵器放在粘有双面碳导电胶带(8 mm×20 mm,浙江立晶光电科技)的扫描电镜样品台(直径12.7 mm)上;将样品台放于离子溅射仪(Quorum-SC 7620)中,喷金60 s。第五步:扫描观察。将样品台放在扫描电子显微镜(TESCAN-VEGA 3 SBU)中观察和拍照。
1.2.3产卵器石蜡切片的扫描、观察和测量
第一步:固定。分别取产卵器在收缩状态和部分伸出状态时标本,固定于4%多聚甲醛。第二步:脱水、透明和浸蜡。将上述样本先进行乙醇梯度脱水,然后用无水乙醇和二甲苯进行透明,最后浸蜡。第三步:包埋和切片。将上述样本通过包埋机(KD-BM)包埋制作成蜡块,随后用石蜡切片机(RM2016)切片,最后放进60℃烘箱(DGX-9003B)内烤片。第四步:染色。将上述切片用HE(苏木精-伊红)染色。第五步:扫描观察和测量。首先将上述切片置入倒置荧光显微镜(NIKON ECLIPSE TI-SR)下初步观察,然后在切片扫描仪(Pannoramic DESK, P-MIDI, P250, P1000)下将所需要的部分进行扫描成像,最后用Caseviewer 2.3软件对产卵器的各部分长度进行测量。
1.2.4基于透射电子显微镜对产卵针内部超微形态结构进行观察和拍照
首先将解剖雌虫的产卵器的产卵针样本固定在1%锇酸中,然后放于不同浓度的乙醇及丙酮中脱水,接着用包埋剂包埋,放于60℃烤箱中聚合,将其置于超薄切片机(Leica UC7)中进行切片,紧接着使用2%醋酸铀染色,再在2.6%枸橼酸铅溶液中染色,最后在透射电子显微镜下观察(HT7800/HT7700),采集图像分析。
1.3 命名参考
产卵器的各部分名称参考McAlpine(1973)、McAlpine(1977)、Freidberg and Mathis(1986)和Norrbom and Kim(1988)。感受器的命名参考Dickensetal.(1988)、张国娜(2012)和宋傲群等(2015)。肌肉命名参考的是Berube and Zacharuk(1983)和Ovtshinnikova(2012)。
2 结果与分析
2.1 基于光学显微镜对柑橘大实蝇产卵器的形态结构观察与测量
柑橘大实蝇成虫产卵器分为产卵器基节、翻缩膜和产卵针(图1),基节分为基节基部与基节端部,翻缩膜分为骨化部和膜质部,骨化部又分为骨化带和骨化环,产卵针分为基部和端部。产卵器基节、产卵器基节基部和产卵器基节端部的长度分别为5.292±0.077(M±SE,下同)、3.319±0.036和1.972±0.057 mm,其宽度分别为1.975±0.024、1.522±0.023和0.453±0.008 mm;翻缩膜、骨化部、骨化带、骨化环和膜质部的长度分别为5.296±0.114、2.662±0.046、1.772±0.036、0.891±0.024和2.634±0.096 mm,其宽度分别为0.870±0.025、0.574±0.018、0.301±0.008、0.288±0.007和0.296±0.010 mm;产卵针、产卵针基部、产卵针端部的长度分别为5.114±0.079、4.815±0.080和0.300±0.016 mm,其宽度分别为0.290±0.008、0.164±0.004和0.126±0.005 mm;产卵器的长度为15.695 ±0.235 mm。测量发现,产卵器基节、翻缩膜和产卵针的长度几乎相等。
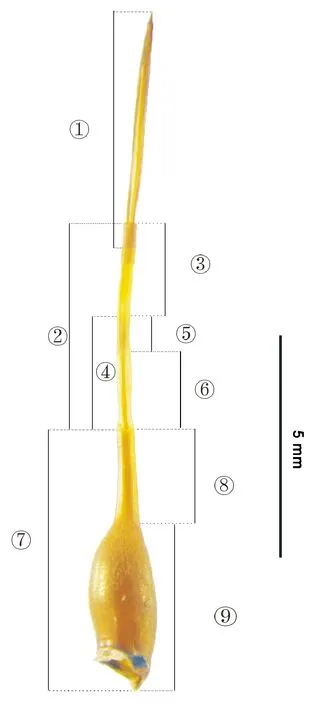
图1 柑橘大实蝇产卵器结构Fig.1 Composition of ovipositor of Bactrocera minax 注:①产卵针;②翻缩膜;③膜质部;④骨化部;⑤骨化环;⑥骨化带;⑦基节;⑧基节端部;⑨基节基部。Note:① Aculeus;② Eversible membrane;③ Membranous section;④ Sclerotized section;⑤ Sclerotized ring;⑥ Sclerotized belt;⑦ Oviscape;⑧ Oviscape distal;⑨ Oviscape basal.
产卵器基节是由连续硬化的角质层组成。呈棕褐色,漏斗状(图1),其表面着生大量细小刚毛。在产卵器基节基部靠近腹部第6体节处,有1对骨片和1个圆形透明膜(图2-A)。产卵器基节基部两侧各有1个气门(图2-B)。翻缩膜是浅黄色的柔软的管状结构,其基部与产卵器基节相连,端部与产卵针相连。产卵器基节与骨化带的交界处呈齿状(图2-C)。在翻缩膜的近端为骨化带,有4条纵向的、褐色的柔化的离散带(图2-D),其中两条位于背内侧,另外两条位于腹面外侧(图2-E),其离散带上有密集的棕褐色颗粒状斑点,离散带之间由柔性的角质层连接。骨化环近端与骨化带相连,远端与膜质部相连。在骨化环背面和腹面有棕褐色的骨片且网状排列(图2-F),其侧面也有骨片,但未完全闭合,中间有一条浅黄色的缝隙(图2- G)。膜质部远端与产卵针基部相连,膜质部浅黄色,其表面的骨片与骨化环的骨片相似,但比骨化环的骨片小(图2-H)。产卵针在自然状态下伸出时,产卵针基部常常有一小部分套叠于膜质部内(图2-I)。产卵针褐色、剑状,末端尖锐(图2-I)。
在产卵器完全收缩状态下,翻缩膜和产卵针都收缩于基节内(图2-A),基节端口呈菱形,由4条骨化带上的离散带组成4条边(图2-J)。在产卵器部分伸出状态下,骨化带露出,骨化环部分露出,膜质部未露出,产卵针部分露出(图2-K)。产卵器完全伸出状态下,翻缩膜和产卵针均完全从基节中露出,膜质部在伸出或收缩过程中,能够有较大程度的弯曲(图2-L),骨化带和骨化环在伸出或收缩过程中,弯曲程度极低(图2-L),产卵针不能够弯曲(图1)。
2.2 基于扫描电镜对柑橘大实蝇产卵器的超微形态结构观察
产卵器基节基部左右两侧各有1个内闭式气门(图3-A)。基节与骨化带之间的交界处有凹陷,而且其凹陷表面有褶皱(图3-B)。骨化带上的离散带表面有排列紧密的多边形花纹(图3-C),离散带之间的膜质表皮上有不规则且不连续的纵向条纹(图3-D),骨化带与骨化环的交界处,其膜质表面有半圆形的花纹(图3-E)。骨化环和膜质部表面均有角质化鳞片,鳞片的一端固定在表皮上,另外一端未固定,鳞片以覆瓦状排列包围在骨化环和膜质部表面(图3-F,J)。相比于骨化环的鳞片,膜质部表面的鳞片的顶端更圆滑,但鳞片更小(图3-G),其鳞片的端部和中部均有多个小齿状突起,距离产卵针越近,鳞片上的小齿状突起越多(图3-H,I)。

图3 扫描电镜下柑橘大实蝇的产卵器的形态特征Fig.3 Morphological characteristics of ovipositor of Bactrocera minax under scanning electron microscope注:A,基节;B,产卵器基节与骨化带连接处;C,骨化带表面的离散带;D,离散带之间的膜质部分;E,离散带之间的膜质部分与骨化环的连接处;F,骨化环;G,膜质部;H,鳞片;I,膜质部表面的齿状突起的鳞片;J,膜质表面翘起的鳞片;K,产卵针基部突起;L,产卵针背片;M,产卵针腹片;N,泄殖腔;O和P,产卵针端部侧面凹槽;Q和R,产卵针端部;①,毛形感受器4;②,气门;③,泄殖腔;④,毛形感受器5;⑤,腔锥形感受器;⑥,钟形感受器。Note: A, Oviscape; B, Junction between the sclerotized belt and the oviscape; C,Discrete bands of sclerotized belt surface; D, Membranous section between the discrete bands; E, Junction of the membranous section between the discrete bands and the sclerotized ring; F, Sclerotized ring; G, Membranous section; H, Scales; I, Denticulate scales of surface of the membranous section; J, Scales with raised surface of membranous section; K, Protuberance at the base of aculeus; L, Tergite of aculeus; M, Sternite of aculeus; N, Cloaca; O and P, Lateral groove of aculeus end; Q and R, The end of the aculeus; ①, Sensilla trichoid 4; ②, Spiracle; ③, Cloaca; ④, Sensilla trichoid 5; ⑤, Sensilla coeloconic; ⑥, Sensilla campaniform.
产卵针是由1块背片和2块腹片组成,背片高度骨化,腹片短于背片。产卵针的基部(背片)的宽度不等,端部船形,仅由背片部分形成,背片顶端渐尖。产卵针基部的凹槽内有多个多边形花纹(图3-K)。在产卵针的背面中部和腹面中部形成宽度不等的纵向凹槽,背面中部凹槽未到达产卵针端部(图3-L,M)。在产卵针腹面末端的凹槽内有产卵孔、交配孔、排泄孔组成的泄殖腔(图3-M,N)。
柑橘大实蝇产卵器有3种感受器:毛形感受器,腔锥形感受器和钟形感受器。其中毛形感受器4(Sensilla trichoid 4,ST4)(图3-A)整体细长,端部尖细,分布于产卵器的基节表面。毛形感受器5表面光滑,端部弯曲,分布在产卵针背片端部的侧沟中,在侧沟内有5根或7根毛形感受器5(Sensilla trichoid 5,ST5)(图3-O,P)。腔锥形感受器(Sensilla coeloconic,SCo)呈锥状,有圆形凹陷,分布在产卵针的背面(图3-Q)。钟形感受器(Sensilla campaniform,CA)为圆球状,位于凹陷中,分布在产卵针的端部(图3-R)。
2.3 基于石蜡切片对产卵器横切面形态结构观察和测量
在产卵器收缩状态下,观察产卵器基节上A、B、C和D 4个位置横切图(图4),在产卵器部分伸出状态下,观察产卵针、翻缩膜、基节上A、B、C、D和E 5个位置横切图(图5)。翻缩膜和产卵针套叠在产卵器基节内的相对位置,由内及外排列顺序为:内层为产卵针(图4-A~C;图5-B~D),中间层为紧贴在产卵针外层的翻缩膜(图4-A~C;图5-B~D),外层为产卵器基节表皮层(图4;图5-B~E)。中间层与外层之间存在一定间隙,处在产卵器基节基部位置的间隙较大(图4-C;图5-D),处在产卵器基节端部位置的间隙次之(图4-A,B;图5-C)。
产卵针的背片和腹片、以及产卵针内部的直肠、输卵管,因处在产卵器收缩状态或部分伸出状态时的位置不同,其横切面的形态结构也存在一定差异。HE染色后,紫色为细胞质,蓝紫色为细胞核。产卵针的两腹片之间、背片与腹片之间均通过柔性的角质层连接(图4-A,C;图5-A,D)。产卵针背片向侧面延伸包裹整个产卵针的背侧面,且向侧内方延长弯曲呈钩状(图5-A~B)。在产卵器部分伸出状态下,位于产卵针和骨化带位置的切片,其背片中部没有向内凹陷(图5-A~B)。但在基节位置的切片,产卵针的背片中部均有不同程度的凹陷(图4-A~C;图5-C~D)。在产卵针两腹片之间和产卵针两侧面背片与腹片的交接处均向内内陷(图4-A~C;图5-A~D)。除了位于产卵针位置的切片,其产卵针背片两内侧无突起外(图5-A),其余位置的产卵针背片两内侧均向产卵针中心突起。在基节位置的切片,背片两内侧向内突起形成未封闭的圆环(图4-A~C;图5-B~D)。
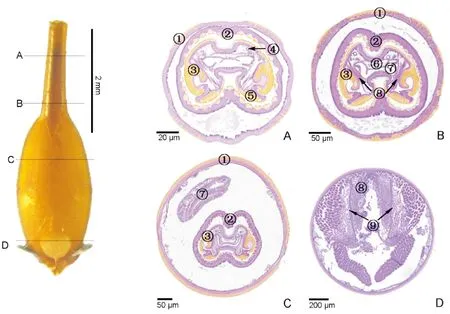
图4 基于石蜡切片的柑橘大实蝇产卵器收缩状态下的产卵器基节内部结构Fig.4 Internal morphological structure of oviscape of Bactrocera minax under the ovipositior contraction based on paraffin section注:横切面图:A,基节端部前端;B,基节端部后端;C,基节基部前端;D,基节基部后端。①, 产卵器基节表皮层;②, 翻缩膜;③, 产卵针背片;④, 气管;⑤, 产卵针腹片;⑥, 直肠;⑦, 输卵管;⑧, 肌肉;⑨, 泵和瓣膜。Note: Cross section: A, Front end of the oviscape end; B, Back end of the oviscape end; C, Front end of the oviscape base; D, Back end of the oviscape base. ①, Cuticular layer of oviscape; ②, Eversible membrane; ③, Tergite; ④, Trachea; ⑤, Sternite; ⑥, Rectum; ⑦, Oviduct; ⑧, Muscle; ⑨, Pumps and valves.
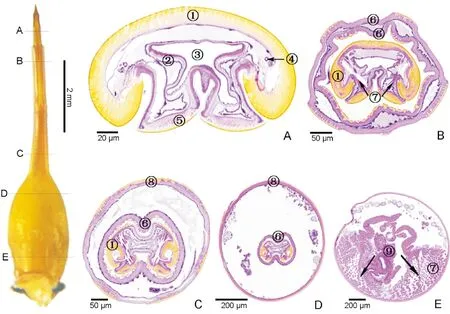
图5 基于石蜡切片的柑橘大实蝇产卵器部分伸出状态下的产卵器内部结构Fig.5 Internal morphological structure of ovipositor of Bactrocera minax under the aculeus partially everted out of sclerotized belt based on paraffin section注:横切面图:A,产卵针;B,骨化带;C,产卵器基节端部;D,产卵器基节基部前端;E,产卵器基节基部后端。①, 产卵针背片;②, 直肠;③, 输卵管;④, 气管;⑤, 产卵针腹片;⑥, 翻缩膜;⑦, 肌肉;⑧, 产卵器基节表皮层;⑨, 泵和瓣膜。Note: Cross section: A, Aculeus; B, Sclerotized belt; C, Front end of the oviscape; D, Front end of the oviscape base; E, Base of the oviscape base. ①, Tergite; ②, Rectum; ③, Oviduct; ④, Trachea; ⑤, Sternite; ⑥, Eversible membrane; ⑦, Muscle; ⑧, Cuticular layer of oviscape; ⑨, Pumps and valves.
靠近腹片与柔性的角质层交接处附近、背片侧面与背片凹陷交接处附近各有1支支气管(图4-C),共4支支气管。背片凹陷的下方为直肠,输卵管位于两腹片间内陷的上方及直肠下方(图4-B)。在产卵器基节内,存在肌肉,输卵管、泵或阀以及血淋巴等组织或器官(图4-C~D;图5-D~E)。
产卵针的横向平均长度为197.7 μm,其纵向平均长度为138.6 μm(图4-A~C;图5-A~D),产卵针切片的横向长度超过纵向长度。产卵针背片的未封闭圆环的横向平均长度为43.1 μm,其纵向平均长度为70.9 μm(图4-A~C;图5-C~D),圆环的横向长度小于纵向长度。背片凹陷的平均深度为43.5 μm,而两块腹片内陷的平均深度为69.6 μm(图4-A~C;图5-C~D),产卵针的腹片内陷的深度更深。上端两支气管间平均距离为93.1 μm,下端两支气管间距离为平均76.9 μm,上端到下端支气管距离相近平均距离为44.0 μm(图4-C)。
2.4 基于透射电镜对柑橘大实蝇产卵针的超微形态结构观察
对产卵针进行横切片,发现产卵针的内部为肌肉、气管、输卵管和直肠等结构(图6-A)。背片和腹片均为角质层,最外一层为包膜,依次是上表皮、外表皮、中表皮,内表皮和真皮(图6-B)。产卵针的腹片向内弯曲,且中表皮层较厚,共5层(图6-A~B)。靠近腹片与柔性的角质层交接处附近、背片侧面与背片凹陷交接处附近各有1支支气管(图6-A)。
第9节段(产卵针)的背腹肌(Tergosternal muscles of segment 9, TSM9))发达,肌细胞的一端着生于腹片内侧(靠近两腹片柔性的角质层交接处一端),另一端着生于背片内侧,斜向连接背片和腹片(图6-A,C)。输卵管和直肠均弯曲呈折叠状,直肠紧贴背片,输卵管靠近腹片(图6-A)。输卵管的管腔小,内壁有数个小褶皱,而且排列整齐。直肠的管腔大,内壁的褶皱比输卵管的褶皱大,褶皱内部有很大孔隙,弯曲弧度大(图6-A,D)。

图6 基于透射电镜的柑橘大实蝇产卵针内部结构Fig.6 Internal morphological structure of aculeus of Bactrocera minax under transmission electron microscopy注:横切面图:A,产卵针部分图片;B,表皮层放大图;C,肌肉放大图;D,输卵管和直肠放大图。Ⅰ,腹片;Ⅱ,背腹肌肉;Ⅲ,输卵管;Ⅳ,直肠;Ⅴ,气管;Ⅵ,背片;Ⅶ,包膜;Ⅷ,上表皮;Ⅸ,外表皮;Ⅹ,中表皮;Ⅺ,内表皮;Ⅻ,真皮。Note: Section: A, Partial picture of the aculeus; B, Enlarged view of the epidermis; C, Enlarged view of muscles; D, Enlarged view of oviduct and rectum. Ⅰ, Sternite; Ⅱ, Tergosternite muscle; Ⅲ, Oviduct; Ⅳ, Rectum; Ⅴ, Trachea; Ⅵ, Tergite;Ⅶ, Envelope; Ⅷ, Epicuticle; Ⅸ, Exocuticle; Ⅹ, Mescuticle; Ⅺ, Endocuticle; Ⅻ, Epidemis.
3 结论与讨论
柑橘大实蝇雌虫产卵器由产卵器基节(腹部第7节)、翻缩膜(腹部第8节)和产卵针(腹部第9节)3部分组成,这些结构组成与其他学者研究的实蝇科昆虫的产卵器组成类似(Drew, 1969; Berube and Zacharuk, 1983; Stoffolano and Yin, 1987)。其骨化带、骨化环、膜质部三者之间除了有明显形态特征差异外,还因为它们之间的弯曲程度也存在差异,如膜质部弯曲程度较大(李宗锴等,2014;桂连友等,2020),骨化带和骨化环弯曲程度极小,因此,本实验将翻缩膜进一步分为骨化部和膜质部,又将骨化部分成骨化带和骨化环。
本研究首次基于石蜡切片观察表明,柑橘大实蝇产卵器在收缩状态时,首先产卵针折叠在翻缩膜内,然后再一起折叠在基节内,因此,形成了产卵针(内层)、翻缩膜(中间层)和基节(外层)3个腹节的套叠。这些套叠结构与其他学者研究实蝇科昆虫产卵器套叠结构类似(Drew and Hardy, 1981; Stoffolano and Yin, 1987; 李宗锴等,2014)。产卵器收缩时,会将产卵针收缩在产卵器基节内,因此,本研究认为产卵器基节呈漏斗状,其形态和大小与产卵针和翻缩膜在收缩于基节内时需要的内部空间密切相关,同时这样的收缩功能有利于在其运动过程中保护产卵针不受伤害,也因其体长缩短,更有利于其在复杂环境中快速运动。
研究结果表明,柑橘大实蝇产卵器基节的基部固定于腹末,端部游离,尽管产卵器基节可以旋转摇摆,但并不能伸出和收缩。产卵针本身不能够伸出、收缩或弯曲,它是随着翻缩膜的伸出、收缩或弯曲而发生运动。柑橘大实蝇产卵器上骨化环和膜质部的表面具有小齿和无齿两种类型的鳞片状结构并且网状排列,Drew and Hardy(1981)研究Ducus(Bactrocera)opiliae的产卵器上也存在这两种鳞片结构。柑橘大实蝇骨化带上有4条离散带,其离散带数量、位置及形态与Drew(1969)研究Urophorauflnis和Urophoraquadrifasciata结果类似。Drew and Hardy(1981)认为D.opiliae的鳞片能增加膜的韧性。苹果实蝇Rhagoletispomonella(Stoffolano and Yin, 1987)、Anastrepha属和Toxotrypana属(Norrbom and Kim, 1988)的产卵器上的鳞片有助于固定腹部或产卵针的基部。骨化带上的离散带及离散带上的颗粒状结构,以及骨化环和膜质部表面有网状排列的鳞片状结构,它们可能在产卵器做径向和轴向运动时起支撑作用,也可以在产卵器伸出、收缩及套叠过程中减少阻力(张占立,2007)。鳞片以覆瓦状排列在骨化环和膜质部表面,这种类似蛇皮表面的腹鳞结构,既可以防止自身的水分蒸发,又对外界的各种理化刺激及冷热变化有一定的抵抗作用,是翻缩膜很好的保护层(张占立,2007)。
柑橘大实蝇产卵针的背面(背片)中线、侧面(背片与腹片连接处)和腹面中线(2腹片连接处)存在宽度和深度不同的纵向凹槽或缝结构,这些结构与其他学者研究实蝇科昆虫产卵针的结构类似(Berube and Zacharuk, 1983)。推测这些结构可能与雌雄成虫交配时,雄成虫的抱握器夹持雌成虫的产卵器行为有关(Eberhard and Pereira, 1993)。
产卵针背片、腹片及背片与腹片之间连接的柔性角质层内存在发达的中皮层(Hackman, 1971),可能是背腹肌(TSM)收缩和舒张交替进行,驱动背片和腹片相对运动,使其背片两侧面向内突起形成1个未封闭的环、操控其背片、侧面和腹面向内凹陷、或调节产卵针内部的径向宽度,从而扩大或缩小产卵针内部的空间。这种结构有利于其成虫排泄和产卵(Stoffolano and Yin, 1987)。
柑橘大实蝇产卵器的表面有毛形感受器、腔锥形感受器和钟形感受器这3种类型感受器。这些感受器位置与类型与其他学者研究的实蝇科昆虫结果类似(Stoffolano and Yin, 1987; 张国娜,2012;宋傲群等,2015)。但在数量上存在一些不同,Stoffolano and Yin(1987)研究认为在苹果实蝇的产卵针端部的两条侧沟内有3对化学感受器,Ritter and Vasey(1989)研究认为在Eurostasolidaginis的产卵针端部的两条侧沟内,其中一条侧沟内为2根化学感受器,另一条为3根化学感受器。本研究的柑橘大实蝇的产卵针端部的侧沟内存在5根或7根两种数量的毛形感受器5(ST5)。
产卵器上的感受器在雌虫交配、产卵及寻找产卵场所过程中具有重要作用。橘小实蝇Bactroceradorsalis的产卵器上的毛形感受器均具有触觉功能,能帮助雌虫选择合适的产卵场所(张国娜,2012)。在产卵过程中,苹果实蝇产卵器上的毛形感受器能监测产卵器的运动,并提供产卵器位置信息(Stoffolano and Yin, 1987),在交配过程中感知产卵器延伸到雄虫生殖器的程度(Hooper and Pitts, 1972)。Luciliacuprina产卵器上的钟形感受器可以给雌蝇提供信息反馈,从而调节产卵量(Rice, 1976)。Aedesaegypti生殖器上的钟形感受器可能只在交配过程中起作用(Rossignol and McIver, 1977)。张国娜(2012)推测实蝇产卵器上的钟形感受器,可能在实蝇选择产卵场所时具有重要的作用。柑橘大实蝇产卵器上的腔锥形感受器可能根据温湿度帮助雌虫选择适宜的产卵时机,并且感受寄主的挥发物质(宋傲群等,2015)。目前,对于柑橘大实蝇产卵器上的3种感受器的具体功能还有待于进一步研究。
有关实蝇科昆虫产卵器运动的操纵机制的文献报道有4篇。如Dean(1935)对苹果实蝇、Hanna(1938)对地中海实蝇Ceratitiscapitata、以及Berube and Zacharuk(1983)对Urophoraaffinis和U.quadrifasciata的进行了详细研究。这些研究均表明,腹部第1~6体节内部收缩,压力减小,腹部血淋巴通过产卵器基节的瓣膜(泵和阀)流入产卵器内,基节内的第1对背侧肌肉DM1 vii收缩,关闭瓣膜;第2对背侧基节肌肉DM2 vii随后收缩,向后牵拉腹侧背内侧叶,使得第7和8段的血淋巴压力增加,从而促使翻缩膜伸长,而产卵器的收缩是其内部的肌肉运动引起的。另外,李宗锴等(2014)研究认为,自然状态下的柑橘大实蝇翻转膜(翻缩膜)在鳞片状纹处内折入基节内,仅露产卵管针突部分,产卵器伸长时翻转膜内叠部分恢复伸展,使产卵管完全露出。产卵针以产卵器基节为中心旋转,产卵器弯曲甚至可超过180°,呈 γ 形。并推测其弯曲原因:在产卵器翻转膜内壁分布有肌肉组织,通过两侧肌肉牵拉,相对应收缩—伸展使得产卵器可弯曲。产卵管要刺穿果皮需要极大的压力,压力则依靠翻转膜的伸展来获得,那么翻转膜内就存在发达的肌肉组织,但李宗锴等(2014)这些推测未经解剖验证。柑橘大实蝇基节内存在泵和阀及其控制泵和阀门的肌肉这一结论还有待验证。目前,柑橘大实蝇产卵器运动的操纵机制并不清楚,有待于进一步研究。
综上所述,柑橘大实蝇产卵器属于伪产卵器,由于长期进化或演化形成这些独特的结构或组织,如(1)产卵器基节呈漏斗状且其内含泵和阀的结构;(2)细长柔软的翻缩膜,其表面有鳞片且网状排列;(3)产卵针由1块高度骨化的背片和2块腹片组成,背片与腹片之间、腹片之间均与柔性角质层(内含发达中皮层)连接,产卵器上有感受器及产卵针内部存在背腹肌。这些结构及组织,一方面为了适应其伪产卵器外翻、弯曲、收缩和折叠的运动机制。另一方面为理解昆虫进化和多样性,以及昆虫的产卵、交配和代谢物排泄等行为机制提供新的例证。
猜你喜欢
今日农业(2024年13期)2024-02-25 00:00:00
云南农业大学学报(自然科学)(2021年1期)2021-01-22 09:23:18
中国现代医药杂志(2020年10期)2020-12-14 07:20:16
热带农业科学(2017年9期)2017-10-23 22:56:24
百科知识(2016年21期)2016-12-24 21:33:29
Acta Mathematica Scientia(English Series)(2016年5期)2016-11-24 11:59:31
蔬菜(2016年8期)2016-10-10 06:49:12
湖南农业(2016年3期)2016-06-05 09:37:36
湖南农业(2016年3期)2016-06-05 09:37:35
西南农业学报(2016年4期)2016-05-17 05:41:45
