
ame-miR-124-3p对西方蜜蜂气味学习的影响
2023-05-27 14:12:20王天宝邱园妹孙巧玲杨长生AqaiKalanHassanyar黄景南李志国苏松坤
环境昆虫学报 2023年2期
王天宝,邱园妹,孙巧玲,杨长生,Aqai Kalan Hassanyar,黄景南,李志国,苏松坤
(福建农林大学动物科学学院(蜂学学院),福州 350002)
蜜蜂是一种真社会性昆虫,它们的神经生理指标可以被记录,相关行为可以直接观察,被认为是研究学习与记忆的良好模式生物(Giurfaetal., 2012; Lafonetal., 2021)。研究表明,蜜蜂能够在实验室条件下的3D虚拟现实(virtual reality, VR)环境中学会根据颜色差异区分奖励和惩罚的刺激(Lafonetal., 2021)。在气味学习训练中,蜜蜂能够将气味与糖水奖励联系起来,表现出很强的联想学习能力,这在蜜蜂外出觅食中发挥着重要作用(Giurfa, 2007)。
miRNA是一种长度大约为21~22 nt的非编码RNA,是一类在转录后水平对mRNA进行负调控的关键调控因子(Bartel, 2018)。与mRNA一样,miRNA的表达模式在空间和时间上都是特定的,每个miRNA可以靶向数十到数百个mRNA,并可能因此调节多个基因网络(Friedmanetal., 2009; Bartel, 2018)。因此,在miRNA表达相对较小的情况下,就可以实现复杂行为的基因调控。
越来越多的证据表明,miRNA参与了昆虫的神经调控。研究发现,miR-132和miR-125影响小鼠大脑的神经可塑性和记忆(Edbaueretal., 2010)。Rajasethupy等人在海兔中发现170个miRNA,并在神经系统中发挥重要作用(Rajasethupathyetal., 2009)。Busto等人通过RNA干扰技术,发现miRNA在果蝇嗅觉学习与记忆上发挥重要作用(Bustoetal., 2015)。据报道,miRNA参与了蜜蜂的学习与记忆过程。研究发现,miR-276和miR-1000在蜜蜂脑部高表达,并通过原位杂交技术发现其富集在蘑菇体的keyon细胞中,有研究报道,蘑菇体是昆虫大脑的一个高级整合中心,蘑菇体外源性神经元(Mushroom body extrinsic neurons,MB Ens)可能参与调控蜜蜂的行为(Haehneletal., 2010; Fillaetal., 2015; Paffhausenetal., 2020),从而推测miR-276和miR-1000参与调控了蜜蜂的神经功能(Sayaka Hori, 2011)。此外,Cristino等人通过嗅觉学习实验发现miR-932的抑制会损害长期记忆的形成,miR-932通过靶向肌动蛋白,调控蜜蜂的记忆(Cristinoetal., 2014)。Qin等人通过迷宫训练实验,也发现miRNA可能在蜜蜂的学习和记忆过程中发挥关键作用(Qinetal., 2014)。这些研究表明,蜜蜂学习行为的变化会伴随着脑部miRNA的表达变化,miRNA在一定程度上参与了蜜蜂的学习与记忆过程。虽然研究人员对miRNA研究较多,但对于其如何调控蜜蜂学习行为的确切机制仍知之甚少。
在本实验室团队之前的研究中,通过利用small RNA-seq(sRNA-seq)技术对12日龄学会与学不会的西方蜜蜂Apismellifera个体脑部进行高通量测序,筛选出14个差异表达miRNA,其中ame-miR-124-3p在学会的蜜蜂组中表达上调,根据以往文献报道ame-miR-124-3p可能是西方蜜蜂学习的关键调控因子(Qinetal., 2014; Michelyetal., 2017),但ame-miR-124-3p对西方蜜蜂气味学习的作用没有得到直接的实验验证,其分子机制尚不明确。为了继续研究ame-miR-124-3p在调控蜜蜂气味学习的作用,在本试验中发现,ame-miR-124-3p的表达抑制降低了蜜蜂的学习比例,ame-miR-124-3p的过表达则能增强蜜蜂的气味学习能力,这表明ame-miR-124-3p在调控蜜蜂气味学习中起着重要作用。
1 材料与方法
1.1 供试蜜蜂
西方蜜蜂来自福建农林大学动物科学学院(蜂学学院)实验蜂场(福建,福州)。从3群健康蜂群中提取3张成熟子脾,置于3个干净的限王产卵框中,并放在恒温培养箱(温度34℃,相对湿度70%)中培养。每天用不同颜色的油漆笔标记刚出房的蜜蜂工蜂,记为1日龄,每天标记的蜜蜂大概在400~500头,将标记好的蜜蜂放入原蜂群。
1.2 干扰序列设计
根据ame-miR-124-3p的成熟体序列,委托上海吉玛制药有限公司(GenePharma)设计合成ame-miR-124-3p的抑制剂(ame-miR-124-3p inhibitor)、ame-miR-124-3p抑制剂的无意义序列(ame-miR-124-3p inhibitor-NC)、ame-miR-124-3p的模拟物(ame-miR-124-3p mimics)和ame-miR-124-3p模拟物的无意义序列(ame-miR-124-3p mimics-NC)(表1)。

表1 干扰序列及相关信息
1.3 蜜蜂的固定与合成序列的饲喂
待到蜜蜂11日龄时,用软镊子抓取并放入透气的玻璃瓶中,放置冰上冻晕,用胶带将蜜蜂快速固定于特制的铜管中,使其吻部可自由活动而露出的头部不可随意转动。将固定好的蜜蜂喂食10 μL含每种合成试剂6.6 μg的50%蔗糖溶液(Liuetal., 2017),所有的蜜蜂在处理后都用50%蔗糖溶液喂饱,置于黑暗培养箱中(28℃,相对湿度70%),24 h后进行气味联想性学习训练(Matsumotoetal., 2012)。
1.4 气味联想学习训练
气味联想性学习实验的训练方式参照Matsumoto等创建的PER标准方法(Matsumotoetal., 2012)。实验前用30%糖水触碰蜜蜂触角,只有表现出伸吻反应的蜜蜂才被用于实验。训练时采用A、B(A,1-壬醇;B,正己醇)两种不同的气味,A气味为条件刺激(Conditional Stimuli,CS),糖水奖励为非条件刺激(Unconditional Stimuli,US),A气味与糖水奖励配对训练(CS+),B气味作为对照气味,不与糖水奖励相配对(CS-)。为了排除顺序对实验的干扰,每组一半的蜜蜂按照ABBAAB的顺序,另一半的蜜蜂按照BAABBA的顺序,对每头蜜蜂分别进行3次联想性学习和非联想性学习。联想性学习的训练时间是42 s。先将蜜蜂静置10 s,再单独通入5 s的空气;随后通入4 s的A气味给予条件刺激,同时使用30%糖水触碰触角,使其伸吻,持续时间为2 s;而后让蜜蜂取食糖水1 s,最后再通空气20 s。非联想性学习训练是通B气味时不给予糖水奖励,每次联想性学习和非联想性学习训练的时间间隔为10 min。实验重复进行5次,每组至少有50头蜜蜂接受气味学习训练。本研究认为只有在蜜蜂触角接触到气味时,蜜蜂立即伸吻并表现出强烈摄食动作才算学会CS关联US,记为“1”,未表现出伸吻反应则记为“0”,通过蜜蜂对气味的伸吻反应率来评估3次实验中蜜蜂的学习情况。训练结束后立即用液氮将蜜蜂冻毙,获取蜜蜂头部并储存于-80℃超低温冰箱,用于后续脑部解剖。
1.5 qRT-PCR检测相对表达量
低温下解剖蜜蜂脑部,放入RNA-Free EP管中,每管3头,每组设置12个生物学重复。利用miRNA Design软件(Vazyme公司,中国)设计特异性的Stem-loop引物、上游引物和通用的反向引物,选择R为通用引物(表2),U6作为内参基因(Liuetal., 2019)。委托上海生工生物工程有限公司合成引物。通过磁珠法,利用LabServ Universal RNA kit分别提取处理组和对照组总RNA,按miRNA 1stStrand cDNA Synthesis Kit (by stem-loop)(MR101-01/02)(Vazyme公司,中国)说明书进行总RNA的茎环反转录。反转录体系为10 μL,5×g DNA Wiper Mix:1 μL,总RNA:v μL,Stem-loop primer(2 μM):1 μL,10 × RT Mix:1 μL,Hiscript II Enzyme Mix:1 μL,RNase Free ddH2O:6-v μL;反转录条件为:25℃ 5 min,50℃ 15 min,85℃ 5 min,得到的cDNA作为模板进行qPCR。qPCR反应体系为10 μL:SYBR Green Dye 5 μL,特异性上游引物0.2 μL,通用下游引物0.2 μL,cDNA模板1 μL,RNA-Free H2O 3.6 μL。反应条件为:95℃ 5 min,95℃ 10 s,60℃ 30 s,共40个循环,熔解曲线反应条件为:95℃ 15 s,60℃ 1 min。最后用2-ΔCT的值来计算相对表达量(Ishiguro, 1992)。

表2 用于荧光实时定量 PCR的引物序列
1.6 统计分析
利用SPSS 16.0软件对PER和qPCR数据进行t检验,GraphPad Prism 5.0软件进行结果的绘图。
2 结果与分析
2.1 ame-miR-124-3p抑制与过表达对气味学习的影响
ame-miR-124-3p的抑制组与ame-miR-124-3p抑制的对照组的有效样本数分别为54头和62头,与ame-miR-124 inhibitor-NC组(80.47%)相比,饲喂ame-miR-124 inhibitor组的蜜蜂经过3次训练后学习比例(63.17%)显著下降(t=3.081,P<0.05)(图1),这表明ame-miR-124-3p的抑制能够减弱蜜蜂的气味学习行为。ame-miR-124-3p的过表达组与ame-miR-124-3p过表达的对照组的有效样本数分别为62头和65头,与ame-miR-124-3p过表达对照组(67.73%)相比,饲喂ame-miR-124-3p过表达组的蜜蜂学习比例(84.39%)显著上升(t=2.771,P<0.05)(图2),说明ame-miR-124-3p过表达能够增强蜜蜂的气味学习行为,这表明ame-miR-124-3p可能参与气味学习的正向调控。
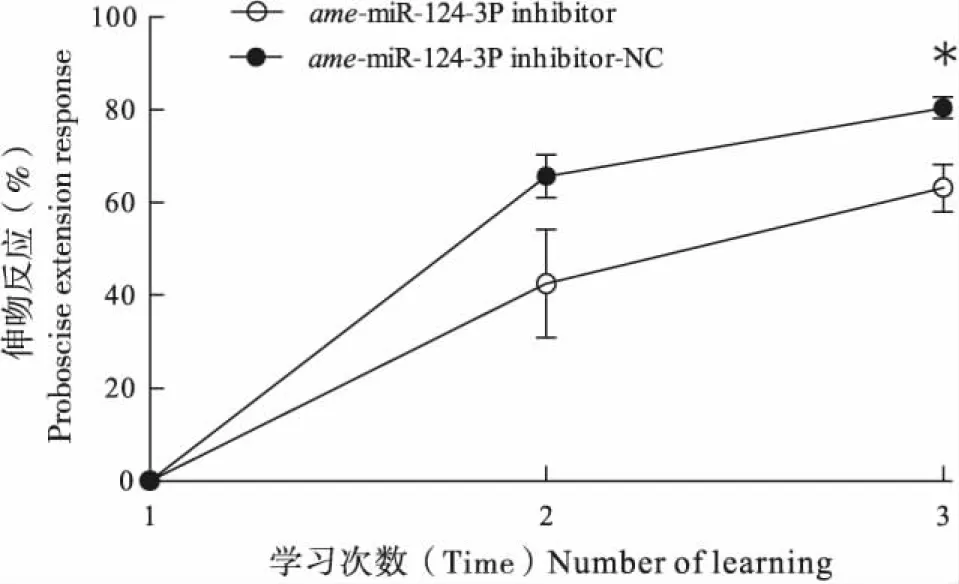
图1 ame-miR-124-3p抑制对西方蜜蜂气味学习的影响Fig.1 Effect of inhibition of ame-miR-124-3p on odor learning in Apis mellifera注:*表示在第3次学习试验中ame-miR-124抑制组与ame-miR-124抑制对照组中的伸吻反应率经t检验显著性差异(P<0.05)。Note: Single asterisk indicated that proboscis extension response rates were significantly different by t-test between ame-miR-124-3p inhibitor and ame-miR-124-3p inhibitor-NC from the 3rd learning experiments.
2.2 qRT-PCR验证气味学习后蜜蜂脑部ame-miR-124的相对表达水平
与ame-miR-124-3p抑制剂的无意义序列组相比,饲喂ame-miR-124抑制剂组蜜蜂脑部ame-miR-124相对表达量水平显著降低(t=2.655,P<0.05)而与ame-miR-124-3p模拟物的无意义序列组相比,ame-miR-124-3p模拟物组蜜蜂脑部的ame-miR-124-3p相对表达量水平显著升高(t=2.114,P<0.05)(图3),这揭示了ame-miR-124-3p在调控蜜蜂的气味学习中起着重要作用。
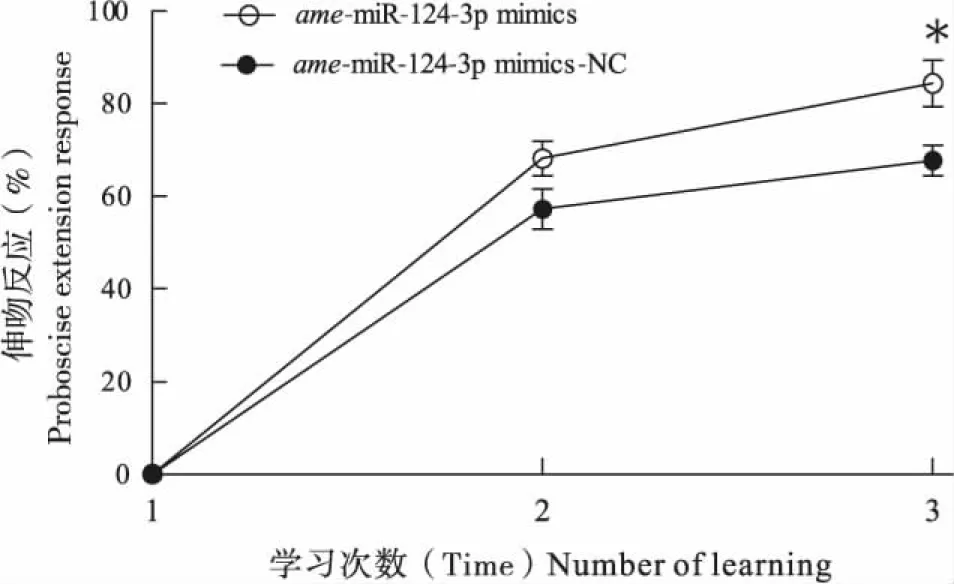
图2 ame-miR-124-3p过表达对西方蜜蜂气味学习的影响Fig.2 Effect of overexpression of ame-miR-124-3p on odor learning in Apis mellifera注:*表示在第3次学习试验中ame-miR-124-3p过表达组与ame-miR-124-3p过表达对照组的伸吻反应率经t检验呈显著性差异(P<0.05)。Note: Single asterisk indicated that proboscis extension response rates were significantly different by t-test between ame-miR-124-3p mimics and ame-miR-124-3p mimics-NC from the 3rd learning experiments.
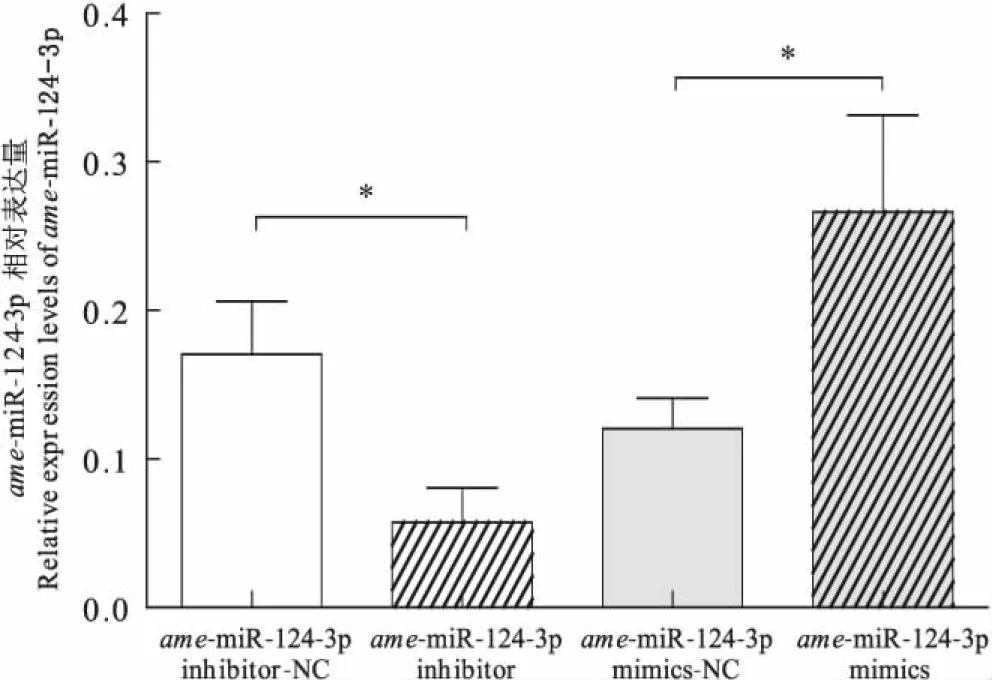
图3 ame-miR-124-3p的相对表达量Fig.3 Relative expression levels of ame-miR-124-3p注:*表示经t检验呈显著性差异(P<0.05)。Note: Single asterisk indicated that proboscis extension response rates were significantly different by t-test.
3 结论与讨论
学习是动物在自然界中赖以生存的重要能力,相对于大型动物,蜜蜂大脑体积小,包含的神经元数量少,但蜜蜂同样有复杂的社会行为(Lafonetal., 2021)。因此,利用蜜蜂作为模型动物由浅入深的研究学习机制极其重要。研究报道显示,12日龄的蜜蜂已经具有稳定且较好的气味学习能力(Laloietal., 2001; 邱园妹等,2020)。在本试验中只选取11日龄的蜜蜂作为实验对象,排除了日龄对实验的干扰。气味学习过程中提供两种较容易区分的气味(1-壬醇和正己醇),可减小蜜蜂对特定气味的喜好所产生的偏差(Rayetal., 1997; Michelyetal., 2017)。
目前,许多研究都在探索蜜蜂的气味学习分子机制。Wang等人将气味学习训练后的蜜蜂大脑与未经过气味学习训练的蜜蜂大脑进行基因表达谱(Digital gene expression,DGE)测序发现 259个差异表达基因,这为探究蜜蜂的气味学习与记忆的机制提供了重要线索(Wangetal., 2013)。Farooqui等人发现章鱼胺介导触角叶早期加工阶段嗅觉记忆组成部分的巩固(Farooquietal., 2003)。另外,Cristino等人研究发现miR-932的抑制能够损害气味学习后长期记忆的形成,mir-932靶基因的验证发现,miR-932与肌动蛋白相互作用,这表明miR-932可能能靶向肌动蛋白来调控蜜蜂的记忆形成(Cristinoetal., 2014),为miRNA参与蜜蜂学习与记忆提供了直接证据。此外,Michely等人在蜜蜂气味联想学习训练前4 h注射miR-124抑制剂,发现蜜蜂在3次气味联想学习后2 h的短期记忆和24 h的长期记忆显著降低,这说明miR-124在条件反射诱导的瞬变记忆的早期阶段起着重要作用(Michelyetal., 2017)。Cristino等人和Michely等人的研究虽然很好地阐述了miRNA与蜜蜂记忆的关系,但未揭示miRNA对蜜蜂气味学习能力的影响。
本研究通过miRNA的抑制和过表达发现了ame-miR-124-3p在调控蜜蜂气味学习能力中起着重要作用。ame-miR-124-3p被抑制后,蜜蜂的气味学习能力降低(图1),蜜蜂脑部ame-miR-124-3p的相对表达量显著降低(图3);而ame-miR-124-3p的过表达则会增强蜜蜂的气味学习能力(图2),这表明ame-miR-124-3P可能是调控蜜蜂学习能力的重要因子,这与Michely等人的研究发现产生了紧密的关联。以往的研究表明,miR-124在果蝇的中枢神经系统中表达(Starketal., 2005),也在鸡脊髓神经前体细胞中高度表达(Caoetal., 2007);miR-124是小鼠大脑含量最丰富的miRNA,占所有miRNA在大脑表达的25%~48%(Lagos-Quintanaetal., 2017)。用miR-124转染小鼠神经干细胞发现,miR-124能够诱导神经干细胞、刺激神经元的分化(Silberetal., 2017)。Cheng等人在小鼠脑室下区(Subventricular zone,SVZ)的活体实验发现,miR-124使SRY-box表达下调,从而抑制前体分裂并刺激分化(Chengetal., 2009)。Rajasethupathy等人通过分析miRNA在海兔不同组织中产生的克隆数量,确定每个miRNA的丰度和组织分布,他们发现含量最丰富的是miR-124(Rajasethupathyetal., 2009)。这些发现说明在果蝇、鸡、小鼠和海兔中,miR-124都是神经发生的重要调控因子。此外,Qin等人利用SRNA-seq技术,发现经过视觉学习后的蜜蜂个体脑部miRNA表达量出现变化,并检测出40个差异表达的已知miRNA,其中包含miR-124,这表明视觉学习的蜜蜂会在条件化之后直接诱导不同miRNA水平的变化,特定的miRNA可能通过调控学习,从而促进蜜蜂记忆的形成(Qinetal., 2014)。这些研究结果表明miR-124在各种物种中相对保守,并可能参与动物的神经调控。
miRNA在突触发育的不同阶段发挥着广泛的作用,包括树突形成、突触形成和突触成熟(Schratt, 2009)。此前有报道称miR-932位于蜜蜂神经连接蛋白2(Neuroligin 2, Nlg 2)基因的内含子2中(Biswasetal., 2008),并且已被证明参与脊椎动物和无脊椎动物的谷氨酸和γ-氨基丁酸能突触的发育(Biswasetal., 2008; Chenetal., 2012)。神经可塑性是指由于经验原因引起的大脑的结构改变。大脑由神经元细胞和神经胶质细胞构成,这些细胞互相连接,通过加强或削弱这些连接,大脑的结构可以发生改变。气味学习是一种会导致大脑功能和结构改变的一种行为(Haehnel, 2010; Paffhausenetal., 2020),miRNA可能是通过影响神经可塑性了来影响蜜蜂的气味学习能力。突出可塑性是蜜蜂学习与记忆的基础,研究表明,突出可塑性的长期保持有赖于蛋白质的合成(Zoghbi, 2003)。因此,在无脊椎动物模式生物中,miRNA与突触可塑性、学习和记忆的调控有关。以上表明,miR-124可能是通过影响蜜蜂神经可塑性进而调控气味学习能力,最终影响其早期记忆,miRNA与神经可塑性之间的联系需要进一步探索。
综上,抑制ame-miR-124-3p的表达能够降低蜜蜂气味学习的能力,ame-miR-124-3p过表达则能增强蜜蜂的气味学习能力,这表明ame-miR-124-3p在调控蜜蜂气味学习能力中起着重要作用。本实验结果有助于从分子水平上了解蜜蜂气味学习行为和大脑功能之间的联系。
猜你喜欢
文苑(2020年7期)2020-08-12 09:35:58
家庭医学(下半月)(2020年3期)2020-05-30 12:42:06
小学科学(学生版)(2017年11期)2017-12-20 05:56:01
小天使·一年级语数英综合(2017年9期)2017-10-20 21:12:02
家教世界·创新阅读(2017年7期)2017-08-09 00:57:53
奥秘(2016年6期)2016-07-30 17:34:30
阅读与作文(小学高年级版)(2016年5期)2016-05-10 23:32:23
分忧(2016年3期)2016-05-05 01:58:43
阅读与作文(小学低年级版)(2016年1期)2016-03-12 22:20:48
红蜻蜓·低年级(2015年11期)2016-02-02 10:55:46
