
10 kV树线故障树木暂态阻抗变化特性试验研究
2023-05-27 13:52赵燊元陈天翔徐会凯杨博闻任欣悦范松海吴驰
广西大学学报(自然科学版) 2023年2期
赵燊元, 陈天翔*, 徐会凯, 杨博闻, 任欣悦, 范松海, 吴驰
(1.成都理工大学 核技术与自动化工程学院, 四川 成都 610059;2.国网四川省电力公司电力科学研究院, 四川 成都 610072)
0 引言
树线故障引起输电线路跳闸和山火已经成为电网发展面临的一个重要问题[1-7]。树线故障引发的原因一般是由于树木过度生长或在风偏的作用下接触输电线路引起树线放电和着火,从而使输电线路发生故障[8-11]。树线故障作为众多输电线路故障中的一种,大多数学者将其归类为高阻接地故障[7]。
目前,国内外许多学者的研究主要是针对树线故障下的电弧进行研究,将多种电弧外特性模型引入输电线路树线故障模型,用以研究分析电弧的非线性特性对输电线路的电气参数和绝缘的影响,而忽略树木阻抗和内部的变化特性[12-17]。在实际情况中,不同地区引起树线故障的树木种类、内部结构、影响因素各不同,因此,研究树线故障除了研究树线放电的电弧特性,还需要研究树木暂态阻抗变化特性。
现阶段,针对树线故障下树木暂态阻抗变化特性的研究很少,主要集中在生物学领域,如研究树木电阻和电导率的测量、树木内部含水率的测量以及两者在判断树木内部缺陷与病害方面的应用。国外学者Lin[18]提出木材比电阻d和含水率u与温度T的关系,概括为d=f(u,T)。利用这一关系,金照怿等[19]设计了电导式木材测湿仪,利用树木电导与含水率的关系,准确测量树木的含水率。鲍震宇等[20]将电阻测试法应用于木材腐朽检测技术,曾祥君等[21]计算出了单相接地发生前后故障相和非故障相电流之间的变化关系,在稳态下根据故障相电压与泄漏电流之间的比值计算出过渡电阻的大小。董俊等[22]进行了零序分量与过渡电阻的关联性分析,计算不同接地方式下的零序电压和零序电流,推导出了零序电压与过渡电阻成反比关系,故障电流与零序电压成正比关系。这些方法可以参考应用于树木阻抗测量研究中,但无法有效地研究高电压下树木暂态阻抗的变化特性。
本文通过搭建10 kV树线故障试验研究平台,选取2种树线故障频发的典型树木——松树和竹子,进行高电压下树木暂态阻抗变化特性试验研究,分析不同树木对树线故障下树木暂态阻抗的影响因素和变化特性,基于树木内部结构初步建立树木阻抗等效电路,研究结论可为今后防治树线故障提供理论和试验依据。
1 10 kV树线故障试验概况
1.1 试验平台与试验对象
10 kV树线故障试验研究平台如下页图1所示。
试验平台包括2台型号为S11—400/10/0.4、联结组标号为Yyn0的三相电力变压器;6台可以无线控制投切的10 kV电容器柜,可实现电容电流1 A级差调节,电容电流最大可调节为60 A;4台用于无功补偿的0.4 kV电抗器柜。
其余设备还包括最大检测温度660 ℃的红外测温成像仪、采样频率为2 GHz高速故障录波器、精度为1 mm无线遥控升降平台、摄像探头。
试验对象选取山火频发地区的2种典型树木,分别为松树和竹子,如图2所示。分别在松树和竹子的同一长度下选取3组不同含水率的树木,对其分类处理,试验参数如表1所示。

(a) 试验研究平台现场布置示意图

(b) 试验研究平台原理接线图

(a) 松树

(b) 竹子
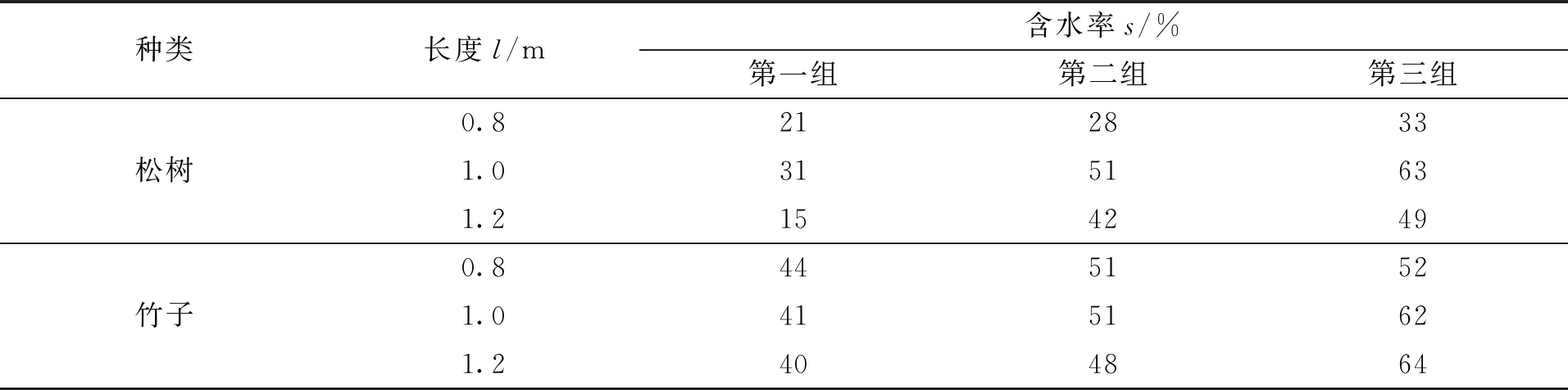
表1 树木试验参数Tab.1 Tree test parameters
实际情况树线故障下的树木大多是树枝顶部接触或靠近导线,多数种类的树木树枝顶部为新生枝丫或嫩芽,该部位半径较小且相近。基于此,本文试验不考虑树木半径对阻抗暂态变化特性的影响。
1.2 试验过程与方法
表1中试验树木含水率的测量方法为:截取试验树木下部5 cm长的块状木材,使用精度为0.01 g的称重仪测量该部位的质量,随后将块状木材烘干至质量没有变化,依据公式(1)计算:
(1)
式中:s为含水率;m1为原始质量,g;m2为烘干后的质量,g。
在试验过程中,保证2种树木的摆放位置、接触导线的相序、室内环境一致。为了便于研究树线故障暂态阻抗变化特性,使用10 kV电压等级试验研究平台,同时调整系统电容电流。为了能够更好地模拟实际树线故障,在升降平台上放置一个铁箱,在铁箱里放入土壤,并定期喷水保持湿润,将新鲜树木栽入土壤中,且正对于故障相导线的正下方,利用升降平台调整树木与导线距离。在铁箱外壁连接铜导线并接入地网,接地网电阻为0.28 Ω,在接地铜导线串接一个1 Ω的无感电阻,用于量取电阻两端电压信号以测量泄漏电流。三相电压及零序电压取自三相导线末端的电压互感器,上述电压信号接入高速故障录波器用以采集三相电压、零序电压及泄漏电流波形,信号采集频率为2 GHz,信号采集与视频录制可同步进行。
为了更好地模拟真实情况,将树木根部与土壤密切接触无缝隙,保留树木上的新鲜树叶。在每组试验进行之前,测量试验树木的含水率,并观察试验树木表面状况及生长情况。为了保证树木顶端能够与导线接触,每次试验前使用升降平台调节导线与树木之间的距离,将红外测温仪与视频录制器放置在试验平台正前方。本文不考虑大气环境因素对试验的影响,所有试验均在相同试验室内部环境中进行,以排除大气环境对试验的干扰。
2 树线故障试验分析
2.1 试验树木与原生树木的一致性分析
由于试验研究平台条件限制,无法利用原生高大松树和竹子进行试验研究,因此用于试验的树木为原生树木上截取而来。为了验证试验树木与原生树木的一致性,在户外采用接地电阻测量仪测量原生树木的阻抗,并结合树木结构与电流理论解释说明该试验方法是可靠的。
首先,使用接地电阻测量仪测量户外原生高大松树的阻抗,经过多次反复测量,平均阻抗为500~600 kΩ,从该松树上截取部分树枝测量阻抗值范围在400~800 kΩ,两者的阻抗值相近。
接地电阻的测量方法为:将两颗地钉打入距被测物体的20、40 m处,分别将两颗地钉连接测试仪内部电压端和电流端接线处,使用BY2517接地电阻测试仪进行电阻测量,测量多次取平均值。
电流的速度一般认为2.997 924 58×108m/s[23],无论对于1~2 m的树枝还是十几米甚至更高的树干,都能在纳秒级时间内形成电流通路,完成电子或导电离子在稳定电流通路内部的定向移动。根据树木内部结构和机制,树木内部的导管和筛管贯通树叶、树枝、树干直到树根,导管和筛管内部为运输树木吸收和产生的水分和有机物,这些水分和有机物不断流通直到树木死亡。由于导管和筛管含水率较高,且内部含有大量不断移动的导电离子,如Na+、K+等[24],因此电导率较大。树线故障形成电弧并击穿密度最高的木栓部后,电子到达导管和筛管处,由于该通路的电导率较大且通透性较好,因此,电流沿着导管和筛管流通。基于此,树线故障发生后,形成的电流通路无论是树枝还是树干都在相同部位,且该部位的电导率与树木的长度和体积没有关系,只与内部的含水率和离子数量有关系。
综上,可以证明试验树木和原生树木的一致性。
2.2 2种树木试验现象分析
松树和竹子在相同试验条件下的试验现象差异较大,除没有其他干扰因素外,电压等级统一为10 kV,电容电流统一为3 A,统一为树木顶端接触C相导线。
当树干接触C相导线瞬间,树干顶端与导线之间会产生微小的电弧,此时树干温度略高于室温(25 ℃);随着时间的推移,树干与导线之间的电弧越来越剧烈,树干顶部的温度越来越高,树干最外层的木栓部被涌入大量的电子能量击穿,使得电子进入树干内部,电流有了新的流通通道,此时,树干顶端的温度大于100 ℃。随着树干往下延伸,温度越来越低,树干最底部温度没有变化。树干顶端开始出现因温度过高以及电弧能量的冲击而导致树干有机物的燃烧,不断侵蚀树干顶端,侵蚀痕迹沿着树干表面不断向下延伸。树干与导线之间因燃烧以及电弧的冲击而出现了间隙,电弧长度随之延长,树干顶端温度随之升高,随着时间的推移,树干火焰燃烧面积越来越大,且沿着树干表面继续向下延伸焦炭通道,在焦炭通道中伴随着电弧不定期地重燃与熄灭。当树干顶端温度达到350 ℃以上时,树干部分区域开始向外喷射水蒸气和其他气体物质,2种气体的颜色明显不同,水蒸气的颜色偏白色,其他气体物质的颜色偏灰色。
当树干与导线之间的空气间隙绝缘强度大于电弧重燃绝缘强度时,电弧不再重燃,树干火焰燃烧与电弧放电随之消失,留下火焰燃烧后的焦炭通道,实验现象如图3所示。这是因为当树干接触导线后,树干与导线之间的接触电阻与周围空气间隙为并联关系,该接触电阻(经测量为105~106Ω)相较于空气间隙电阻要大得多,致使空气间隙被击穿。大量带电质子从导线侧涌入树木侧,形成的能量不断冲击树干顶端,直至树木木栓层被击穿后,电流流入树木内部,通过树木内部的横向木射线,在韧皮部的筛管和木质部的导管形成导电通道。此时,泄漏电流幅值明显增大,在这一过程中,树干顶端温度急剧升高,在固体介质的热效应作用下,温度高于树木内部部分有机物的燃点,使得树木内部有机物被点燃,形成着火现象,并形成前文所述的焦炭通道。
在此过程中,泄漏电流不断增加,最大有效值可增至2 A,在泄漏电流波形周期峰值处出现明显的高频信号,故障相电压小于非故障相电压,且随着树干火焰燃烧与电弧剧烈放电而降低,零序电压不断升高。火焰停止燃烧与电弧熄灭后,三相电压趋于平衡,零序电压为零,泄漏电流为零,树线故障消失,三相线路恢复正常运行水平。
上述试验现象为2种树木试验过程中的共性现象,不同树木种类的试验现象有差异。在相同含水率和长度情况下,松树和竹子试验现象如图3所示。松树的火焰燃烧与电弧剧烈发展的起始时间相对较长,且故障发生后,松树树干接触导线处与其他部分表面出现多处白色黏稠物质,判断该物质为松树内部的松油,由于温度过高随着水蒸气及其他物质被喷发出来,试验结束后温度逐渐降低,在树干表面凝结成白色固态物质。试验过程中,电弧首先在树木与导线接触点处形成,随着时间的推移电弧沿树木径向发展,并且同一部位的着火现象出现在电弧之后,剥开松树树皮可以发现在树皮内部形成了树状痕迹。

(a) 松树焦炭通道

(b) 竹子焦炭通道
竹子是一种比较特别的草本植物,生长时分段,每一段的两端是密封、中间为中空的结构,较难进行气体交换。试验过程中,在电压和电流的作用下,由于通透性较差,竹子内部温度升高,高温下的气体无法及时与外界气体进行交换,因此导致内部气压过高,发生爆竹现象,爆竹后的爆裂痕迹是沿着竹子径向排列。由图3可知,竹子的表面出现与松树内部相同且非常明显的放电或炭化痕迹,但松树必须剥开树皮才能看见。
3 树木暂态阻抗变化特性分析
试验树木接触导线后,造成系统单相接地故障,树线故障示意图如图4所示。
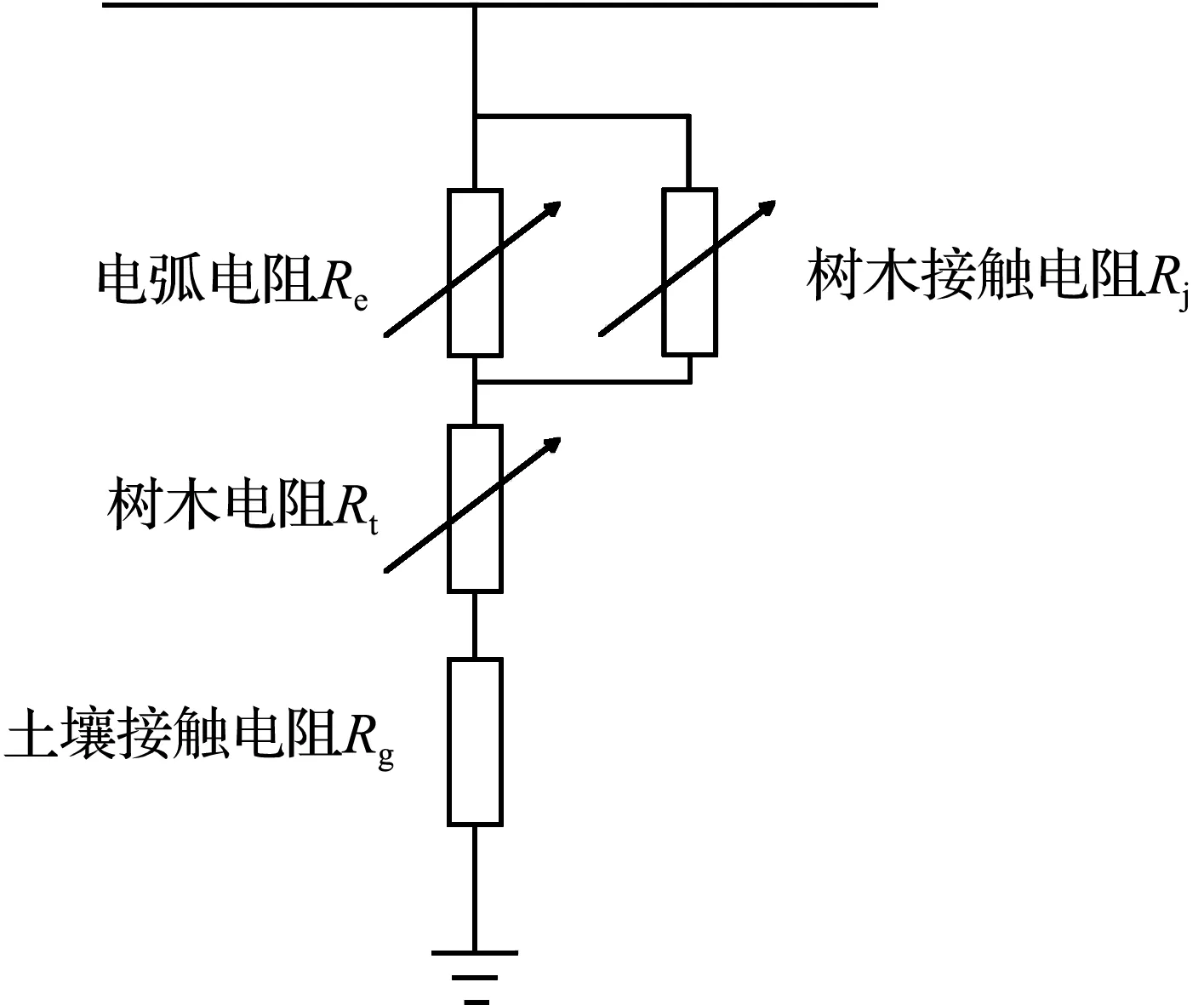
图4 树线故障示意图Fig.4 Schematic diagram of tree line fault
由于试验系统输电导线长度有限,因此无法根据线路长度增大线路对地电容,采取的方式是在变压器一次侧出线端并入用以模拟对地电容的耦合电容器,根据并入电容器容量大小模拟线路对地电容,据此可计算线路电容电流。
根据能量平衡理论,“控制论”电弧模型的一般表达式为[25]
(2)
式中:g为瞬时电弧电导;Gb为稳态电弧电导;τb为电弧时间常数。稳态电弧电导为
(3)
得到稳态电弧电阻为
(4)
式中:i为流经电弧的电流,即泄漏电流;Ub为单位弧长电压,一般取15 V/cm;lb为电弧长度。
在试验过程中,树木顶端与导线之间出现电弧后,除非由于电弧侵蚀或树木受高温影响软化,使树木与导线距离增加大于电弧绝缘熄灭距离时电弧不会重燃,否则电弧以稳态状态在树木与导线之间燃烧,因此,试验过程中的电弧可认为稳态电弧。根据上式计算可得到试验过程中的电弧电阻在1~100 Ω范围内,因此,电弧电阻远小于树木电阻,可以忽略。
当电弧产生并击穿树木表皮后,树木接触电阻与电弧电阻为并联关系,由于电弧电阻远小于树木接触电阻,因此,树线故障发生时树木接触电阻不计入接地电阻中。
土壤电阻率随着土壤湿度和温度的变化而变化,不同的土壤温度和湿度对土壤电阻率影响不同[26],但土壤种类一定且土壤温度与湿度为定值时,不考虑杂质、石头及其他情况,可以确定土壤电阻值,因此,土壤电阻为一定值。在实际环境中,树木的树根根植于土壤之中,即使土壤内部结构含有其他多种杂质,树根也能完全扎根并充分深入土壤内部吸收营养物质,不论是直根系植物还是须根系植物,其根部与土壤的接触面积非常大,根据电阻计算公式:
(5)
式中:ρ为电阻率;s为截面积;l为树木长度。
可以得到:接触面积越大,电阻越小,所以,树木根部与土壤之间的接触电阻可以忽略。综上,接地故障点电弧电阻Re、树木电阻Rt、土壤接触电阻Rg的串联简化为树木电阻Rt。树木电阻Rt的计算公式为
(6)
式中:U0为故障相电压;I0为流经树木的泄漏电流。
3.1 松树暂态阻抗变化特性分析
绝大多数被子植物具有导通的筛管和导管,为活细胞首尾相连而成,其水分和有机物的导通率相对于裸子植物较高。松树属于裸子植物,其内部结构中的运输部位较被子植物不同。松树内部的运输部位由沿树干径向排列的一系列导胞和筛胞构成,这些细胞排列紧密,没有完全把细胞之间的细胞膜褪掉,至今保留着一层膜,用于过滤杂质和运输营养物质,有一定的阻碍作用[19]。
为了分析长度和含水率对松树阻抗变化特性的影响,根据表1分类的松树进行试验,松树阻抗变化特性如图5所示。
(a) 长度为0.8 m松树阻抗变化特性
(b) 长度为1.0 m松树阻抗变化特性

(c) 长度为1.2 m松树阻抗变化特性
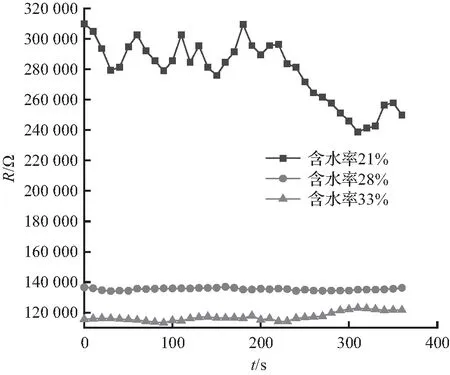
由图5可知,长度为0.8、1.0、1.2 m的松树的阻抗都是随着含水率的增大而减小,含水率越高,松树阻抗越小,其中,长度为1.2 m、含水率为15%的松树阻抗最大,超过了400 kΩ,长度为1.0 m、含水率为63%的松树阻抗最小,为40 kΩ。长度变化对松树阻抗的变化没有明显的影响。
每组试验过程中的松树阻抗都会出现不同程度的变化,且变化趋势不同,含水率较低时,松树阻抗的变化程度较小,含水率较高时,松树阻抗的变化程度较大。长度0.8 m、含水率为21%的松树阻抗总体呈减小趋势,含水率为28%的松树阻抗明显减小,含水率为28%的松树阻抗基本不变,含水率为33%的松树阻抗略有增加;长度1.0 m、含水率为31%的松树阻抗没有明显变化,含水率为51%的松树阻抗呈增大趋势,含水率为63%的松树阻抗总体没有明显变化;长度1.2 m、含水率为15%的松树阻抗没有明显变化,含水率为42%的松树阻抗总体呈增大趋势,含水率为49%的松树阻抗呈下降趋势,较初始阻抗减小了超过50%。9组松树阻抗试验皆呈现含水率越高阻抗越低的规律特性。
在试验发生树线故障这一过程中,一方面,树木的温度越来越高,随着温度升高,树木内部的水分子移动速度加快,加速树木内部水分的蒸发,使树木内部水分减少,从而使树木阻抗增大,同时,松树经过高温后会产生黏性很大的松油物质,该物质的电导率较低,也会使得测量的阻抗增大;另一方面,树木在高温作用下着火后形成炭化通道,炭化通道内部含有空气间隙,在电场作用下该空气间隙会被击穿从而形成稳定的电流通路,测量的树木阻抗会减小。
根据不同含水率和长度的松树试验现象及其阻抗变化特性曲线总结以下几点规律:
① 含水率越大,松树放电和着火现象越剧烈,使得松树阻抗变化越明显;
② 松树阻抗大小与松树含水率呈反比,表现为含水率越大,松树阻抗越小;
③ 松树长度变化对松树阻抗变化无明显相关性。
3.2 竹子暂态阻抗变化特性分析
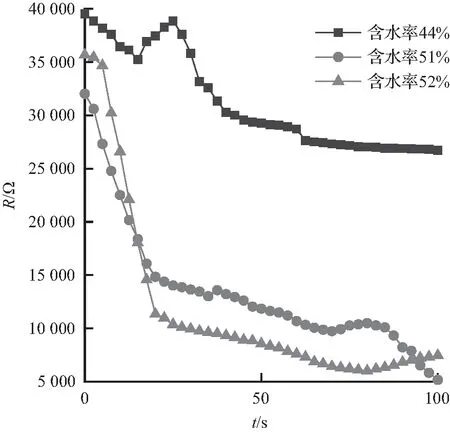
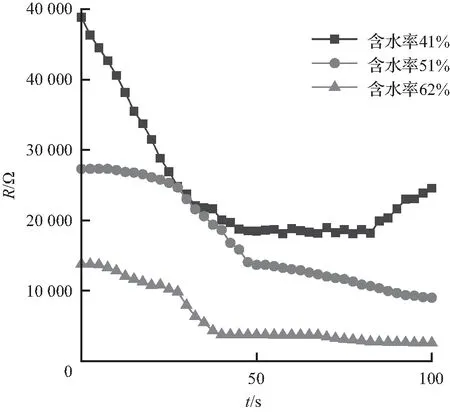
竹子阻抗变化特性如图6所示。图6中,长度为0.8、1.0、1.2 m的竹子的阻抗都是随含水率的增大而减小,含水率越高,竹子阻抗越小,其中,长度为1.2 m、含水率为40%的竹子阻抗最大,为62 kΩ,长度为 1.2 m、含水率为64%的竹子阻抗最小,为2.1 kΩ。长度变化对竹子阻抗变化没有明显的影响。
(a) 长度为0.8 m竹子阻抗变化特性
(b) 长度为1 m竹子阻抗变化特性

(c) 长度为1.2 m竹子阻抗变化特性
长度0.8 m的3组竹子的阻抗均减小,三者的含水率相差不大。初始阻抗相近,含水率为51%与52%的2组竹子在30 s时间内阻抗较初始阻抗减小了超过50%,之后阻抗减小速率较为稳定;长度1.0 m、含水率为41%的竹子阻抗呈现U形曲线先减小后增大,含水率为51%的竹子阻抗呈减小趋势,含水率为62%的竹子阻抗减小幅度较大,最低降至5 kΩ以下;长度1.2 m、含水率为64%的竹子阻抗没有明显变化,其阻抗值也是试验中最小的。9组竹子阻抗试验皆呈现含水率越高阻抗越低的规律特性。
根据不同含水率和长度的竹子试验现象及其阻抗变化特性曲线总结以下几点规律:
① 含水率越大,竹子放电和着火现象越剧烈,竹子阻抗变化越明显;
② 竹子阻抗大小与竹子含水率呈反比,表现为含水率越大,竹子阻抗越小;
③ 竹子长度变化对竹子阻抗变化无明显相关性。
比较松树和竹子两者的阻抗变化特性发现,在实验过程中,松树的阻抗大于竹子的,松树阻抗最大值超过了400 kΩ,而竹子阻抗最大值为62 kΩ,松树阻抗的最小值也大于竹子阻抗的最小值,含水率不同固然是其中的一个原因,另一个原因为两者内部结构不同。
4 树木阻抗变化机制与等效电路
4.1 树线故障树木阻抗变化4个阶段
树线故障下树木放电与树木阻抗的变化过程较为复杂,树木为复杂的有机生命体,内部含有大量鲜活细胞和有机物质构成的不同的组织和结构,这些组织和结构执行着不同的功能和作用,支撑着树木不断生长,所以,树木阻抗变化特性也并不只是单一因素影响,根据试验过程和现象,树木阻抗变化主要分为以下几个阶段:
① 树木接触导线阶段。树木首先接触导线的部位是树木最表面的木栓部,木栓部细胞排列非常紧密,大多为死细胞,其密度和硬度较大,因此短时间内木栓部无法被击穿,此时树木与导线之间的接触电阻极大,大量带电粒子正在高速运动,但尚未形成电子崩和稳定的电流通路。此时温度逐渐升高,能量逐渐增强,木栓部正在被慢慢侵蚀,木栓部的部分物质被能量改变成为固体颗粒或气体形态挥发出来。此时泄漏电流较小,故障相电压变化不明显,树木阻抗较高。
② 树线放电阶段。在电场强度的作用下,树木与导线接触点处的能量不断增强,温度不断升高,在某一时刻,大量带电粒子形成电子崩和稳定电流通路,在树线接触点形成稳定的电弧。木栓部被电弧击穿后,电弧作用至韧皮部,由于韧皮部含水率极大且含有通透率较高的运输结构——筛管,因此电流沿着筛管流通。此时泄漏电流逐渐增大,故障相电压下降明显,树木阻抗逐渐减小。
③ 树木着火阶段。当树线放电发生后,树木温度上升速度逐渐加快,树木温度不断升高,树木内部的水分在高温的作用下被挥发出来。树木内部的有机物质在温度和电弧的双重作用下逐渐气化,通过木射线和木栓部薄弱点挥发出来,并且部分有机物质达到燃点,火焰开始燃烧,树木顶部火焰燃烧至导线,火焰沿着树木径向不断蔓延。当火焰燃烧过后,有机物质在火焰作用下被炭化,形成焦炭通道,焦炭通道内部含有空气间隙,在电场强度作用下,间隙被击穿,电流通路依然存在。此时泄漏电流幅值较高,故障相电压下降明显,树木阻抗继续减小。
④ 树线分离阶段。由于树木顶部温度最高,火焰最先开始燃烧,并且该部位较小,在火焰燃烧一段时间后,该部位被炭化或烧蚀殆尽,使得树木与导线分离,若该空气间隙无法使电弧熄灭,则电弧持续,树线故障继续存在,直到该空气间隙使得电弧不再重燃,树线故障结束。此时泄漏电流变为0,故障相电压恢复正常。
树线故障的4个阶段变化如图7所示。

(a) 树木接触导线

(c) 树木着火

(d) 树线分离
4.2 树木内部结构对树木阻抗变化影响机制
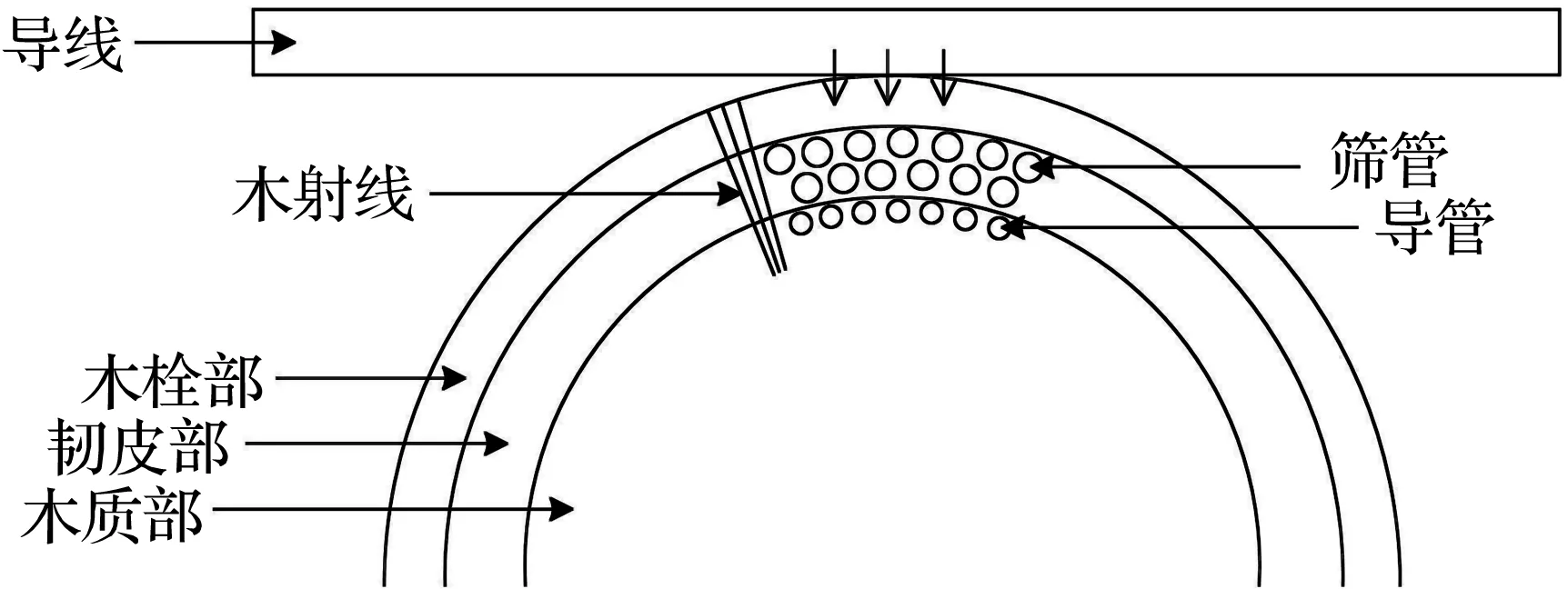
树木的茎沿横向由外向内主要分为木栓部、韧皮部、木质部和髓[23]。木栓部是树木的茎最外面一层,由最外侧的死细胞和内侧的活细胞紧密排列而成,多数细胞呈方形,结构紧凑,主要作用是保护内部结构和支撑树干;韧皮部位于木栓部与木质部之间,韧皮部的含水率较大,营养物质较多,筛管生长、发育在该部位,筛管主要作用是运输根部吸收的水分和养料至树冠,韧皮部的主要作用是向外向内生成新的细胞和组织结构,所以韧皮部多为新生细胞且该部位水分流动性强;木质部为树木茎中占比最大的部位,靠近韧皮部的部分含有导管,导管的主要作用是将树叶经光合作用转化的营养物质和水分运输至根部和其他部位,木质部的主要作用是运输和机械支撑;髓是树木茎最中心的部位,主要作用为支撑和固定。
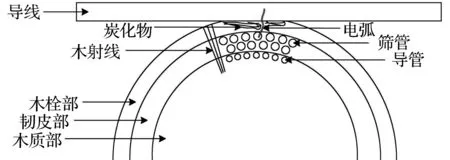
树线故障内部结构示意图如图8所示,图8(a)为树木接触导线起始阶段,图8(b)为树线放电及炭化后阶段,图8(c)为树木内部的电流通路示意图。
树木接触导线初始阶段会出现2种情况:第一种情况,树木与导线的接触点避开了木射线联通外界的开口处,这种情况下导线接触的树木部位是坚硬的木栓部,木栓部的密度较大,导电性低,且没有横向通道,使得此时测得的初始树木阻抗较大,在电弧的冲击与强场强的作用下,木栓部受温度升高与能量作用逐渐分解炭化,导致电流通路转变为含水率高、导电性强的韧皮部。除此之外,木栓部的不同部位也具有差异性,更厚、更坚硬的木栓部部位可能导致树木初始阻抗更大。第二种情况,树木与导线的接触点为木射线联通外界的开口处,木射线是直接将树木的木质部、韧皮部和木栓部相互联通的部位,主要作用为横向运输营养物质,该部位的含水率与导电性相对于木栓部较大,导致初始阶段电流通路就经过木射线流向韧皮部,此时测得的初始树木阻抗较小,在试验过程中树线放电现象也更加剧烈,着火时间更加迅速。树线故障初始接触部位的不同反映的是初始树木阻抗大小的不同以及树线故障早期现象剧烈程度的不同。
(a) 树木接触导线阶段
(b) 树线放电及炭化阶段
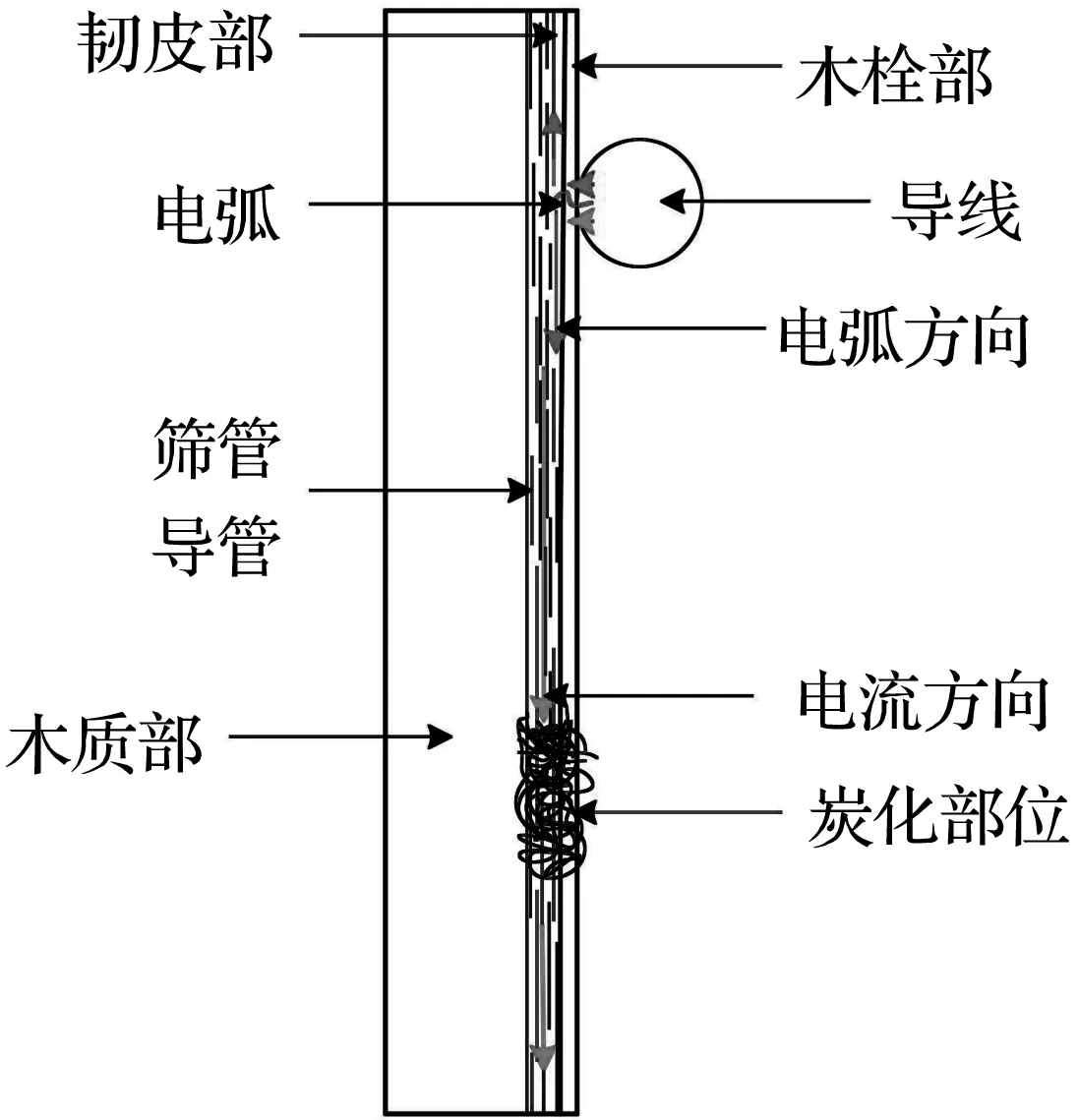
(c) 树木内部电流通路
当电弧击穿木栓部后,电流流向经韧皮部的筛管和木质部的导管沿树木径向流动,根据试验分析,树木的电导性强弱受树木含水率影响较大,水分子与各种导电离子在树木内部充当导电载体,一棵鲜活树木的含水率一般在60%左右,树木内部所有的细胞、组织和结构都含有大量的水,尤其是韧皮部和木质部中的筛管和导管更是运输水与营养物质的主要部位,该部位的含水率较树木其他部位更大,导致该部位的阻抗较其他部位更小。树木阻抗的主要测量部位实际就是树木内部的韧皮部与木质部的筛管与导管这一导电通路的阻抗。木质部较韧皮部的体积更大,含水率更小,越向树木结构中心密度越高,电导率越低,导致其阻抗较韧皮部更大。
电流流经树木内部这一过程中会使树木温度逐渐升高,最高温度能够升至500 ℃以上,在电流冲击与高温的共同作用下,树木枝干不同部位发生着火现象,最终会引起树木部分部位炭化,同时使得树木内部的有机物质发生变化,影响了树木的电导率,当炭化部位连接在一起时,会形成炭化通路。使用LCR电桥测试仪测量炭化部分的树木阻抗约为炭化前的5~10倍,阻抗增大有以下2种原因:一是树木纤维与其他有机物试验后含水率大幅下降,使得电导率减小;二是炭化通路内部含有大量空气间隙,电流通路被阻断。
木栓部与韧皮部在树木结构中的比例较小,影响横向宽度占比最大的是木质部,根据图8(c)树木内部电流通路中可以看出,实际测量树木阻抗的大小与树木木质部没有关系,影响实际测量树木阻抗大小的是木栓部与韧皮部,即树木单位横截面的大小与树木阻抗没有直接关系。
3.1与3.2试验研究证明:松树与竹子都满足含水率越高,树木阻抗越小的变化特性。木栓部的含水率与密度影响着初始树木阻抗的大小,韧皮部与木质部的含水率影响着树木实际测量阻抗的变化特性,水分蒸发与有机物的炭化情况影响着树线故障过程中树木阻抗的变化,即
Za=f(W)。
(7)
松树阻抗随含水率变化的拟合关系为
Zas=1.951×106e-8.426W,
(8)
竹子阻抗随含水率变化的拟合关系为
Zaz=6.488×109e-28.85W,
(9)
式中:Za为树木阻抗;W为含水率;Zas为松树阻抗;Zaz为竹子阻抗。式(8)的拟合系数为0.995 1,式(9)的拟合系数为0.995。
当含水率为一定数值的条件下,树木内部结构影响着树木阻抗的大小与变化特性,具体表现为:若树木与导线接触点为木射线向外界的开口处,则测量的初始树木阻抗相对较小;若接触点没有在开口处,则测量的初始树木阻抗相对较大,当木栓部密度较大或含水率较小时,测量的初始树木阻抗较大,反之较小;当电流流入韧皮部后,韧皮部充当电流通路,韧皮部的含水率越大,测量的树木阻抗越小,反之越大;测量的实际树木阻抗与木质部没有直接关系。
4.3 树线故障树木阻抗等效电路
根据试验现象得到不同树木部位的阻抗值不同,不同部位对树木暂态阻抗变化关系密切,树木横向截面示意图如图9(a)所示。基于树木结构建立了树木阻抗等效电路,如图9(b)所示,简化的树木阻抗等效电路如图9(c)所示。
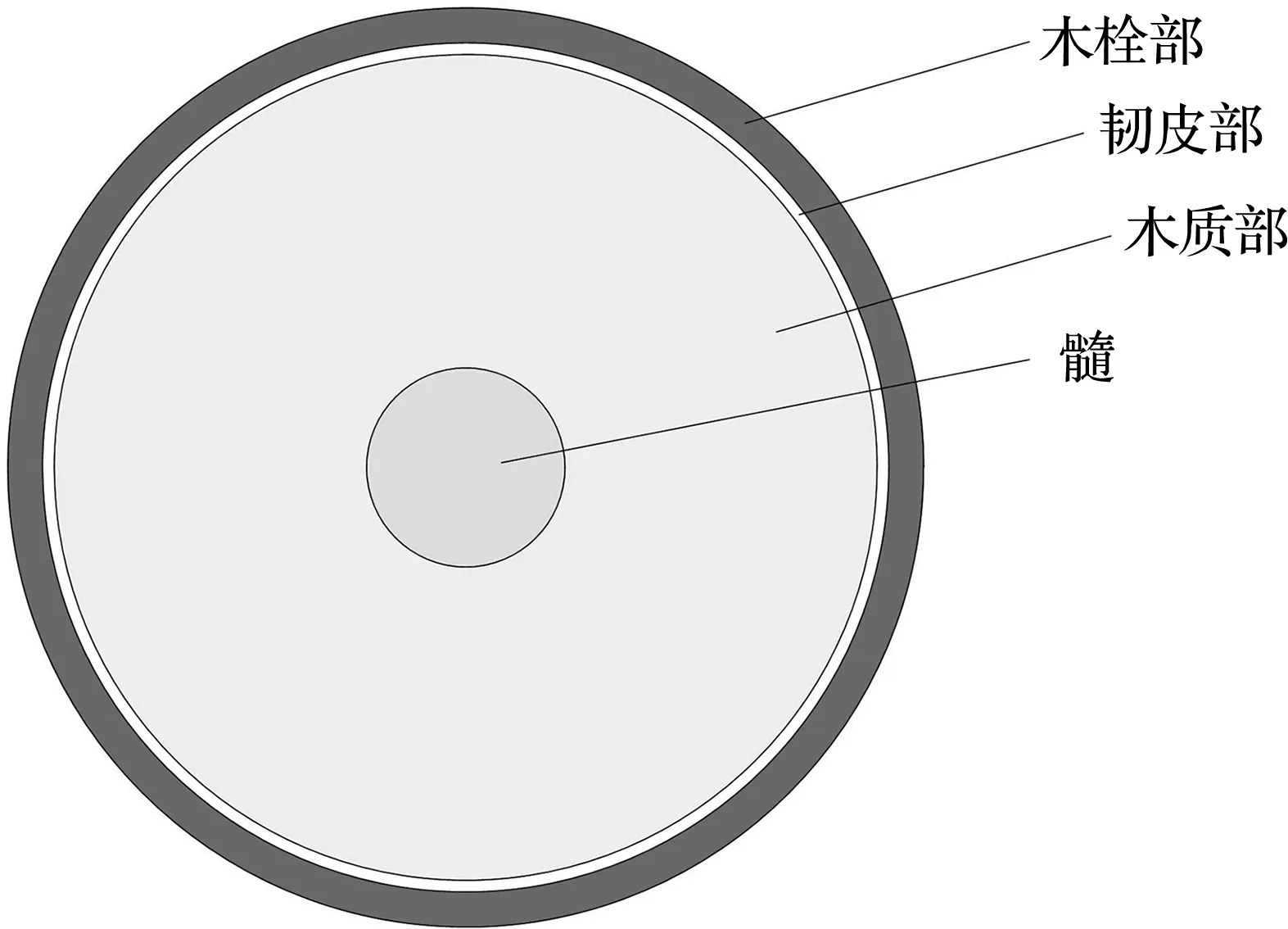
(a) 树木横向截面示意图
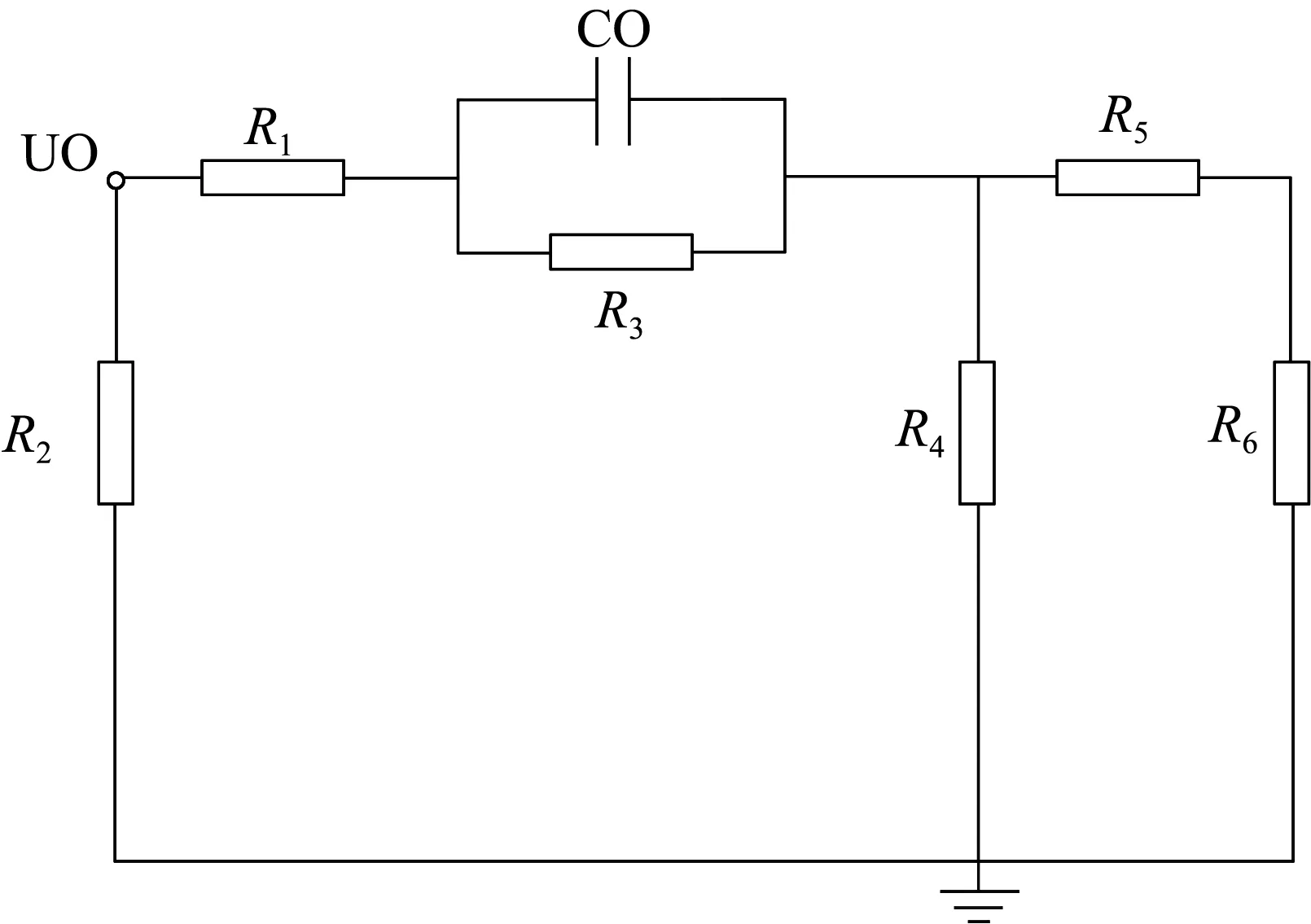
(b) 树木阻抗等效电路
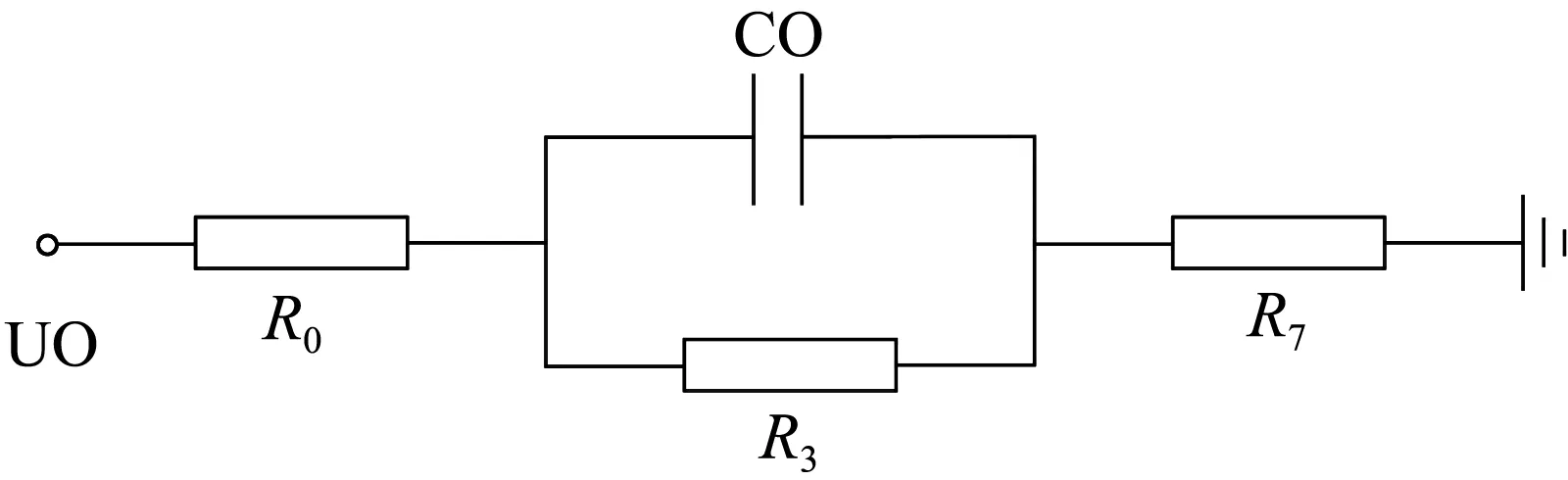
(c) 树木阻抗简化等效电路
图9(b)中:U0为故障相电压;R1、R2为树木木栓部等效电阻;C0为间隙等效电容;R3为间隙等效电阻;R4为树木韧皮部等效电阻;R5为树木木射线等效电阻;R6为树木木质部等效电阻。
图9(c)中:U0为故障相电压;R0=R1//R2为木栓部简化等效电阻;C0为间隙等效电容;R3为间隙等效电阻;R7=R4//(R5+R6)为韧皮部与木质部简化等效电阻。
树木阻抗等效电路是依据不同树木部位的性质和试验测量数据建立的,能够定性地反映不同树木部位之间的电阻大小关系和性质。树木木栓部等效电阻取值范围为100~1 000 000 Ω,测量木栓部与韧皮部之间的阻抗呈容性,间隙等效电容取值为1~10 nF,间隙等效电阻取值为10~100 000 Ω,树木韧皮部等效电阻取值为10~200 000 Ω,树木木射线等效电阻取值为100~500 000 Ω,树木木质部等效电阻取值为100~1 000 000 Ω。影响不同部位等效电阻的最主要因素是含水率,含水率越高,等效电阻越小。
(10)
式中:R1、R2、R3、R4、R5、R6、C0为树木不同部位电阻和电容;Zt为树木等效阻抗;W为树木不同部位阻抗受含水率的影响。
将式(10)简化,得到等效树木暂态阻抗为
(11)
将(11)式化简为
(12)
韧皮部等效电阻与木射线等效电阻、木质部等效电阻为并联关系,且前者等效电阻小于后两者等效电阻。根据前文分析,木栓部被击穿后,树木内部的主要电流通路为韧皮部,因此,木射线和木质部等效电阻对树线故障下的树木暂态阻抗影响较小,将两者等效电阻进行简化。将式(12)简化后,得到实际测量等效树木暂态阻抗为
Zt=f(W(R7))=W(R7)。
(13)
含水率W与树木等效电阻R之间的关系表达式参照式(8)、(9)的拟合公式。
分析树木阻抗等效电路可得:树线故障下木栓部阻抗对树木与导线之间的接触阻抗影响最大;韧皮部阻抗对树线故障下树木阻抗的变化特性影响最大;木质部阻抗对树线故障下树木阻抗没有直接影响;影响上述三者电阻变化的主要因素是含水率。
5 结论
本文搭建了树线故障试验研究平台,选取了易发生树线故障的2种典型树木,在对地电容电流一定条件下,对2种树木的暂态阻抗变化特性进行了试验研究,分析了松树和竹子阻抗的试验现象、影响因素和变化特性,初步建立了树木阻抗等效电路和模型,结论如下:
① 总结了松树和竹子在树线故障下试验现象的规律,树线故障下都会出现冒烟、放电、着火以及炭化现象,树木温度从导线接触点沿径向向两端不断升高,树木炭化后的焦炭通道内部具有空气间隙,可使得树木内部产生局部放电;
② 含水率对树木阻抗的变化特性影响极大,含水率越大,树木阻抗越小,树木阻抗与树木的长度无明显相关性。树线故障下树木的初始阻抗与导线接触点木栓部电阻有关系,接触点阻抗越大,树木初始阻抗越大;
③ 基于树木阻抗变化特性建立了树木阻抗模型,将树木试验现象与阻抗变化分为4个阶段,基于树木结构建立了树木阻抗等效电路,韧皮部等效电阻最小,木栓部等效电阻最大,树木内部结构对树木阻抗变化有着密切关系,影响不同部位阻抗大小的主要因素是含水率;
④ 树线故障树木暂态阻抗变化特性试验研究为后续更深入的研究提供试验基础,为防治树线故障提供试验和理论依据。
猜你喜欢
智能建筑电气技术(2022年2期)2022-02-06
西江月(2021年2期)2021-11-24
动漫星空(兴趣百科)(2020年12期)2020-12-12
小学阅读指南·低年级版(2020年11期)2020-11-16
制造技术与机床(2019年9期)2019-09-10
小天使·一年级语数英综合(2018年4期)2018-06-22
小天使·一年级语数英综合(2017年11期)2017-12-05
儿童时代·幸福宝宝(2017年7期)2017-02-14
电子制作(2017年22期)2017-02-02
少儿科学周刊·儿童版(2015年1期)2015-07-07
