
美洲南瓜Dof基因家族鉴定与表达模式分析
2023-05-27 13:02:53孙守如胡德菊常静依王永马长生朱磊
中国瓜菜 2023年5期
关键词:荧光定量PCR
孙守如 胡德菊 常静依 王永 马长生 朱磊
摘 要:DNA结合单锌指(DNA binding with one finger,Dof)是植物所特有的转录因子,主要参与调控植物的生长发育、抗逆和品质等生命活动过程。为了鉴定美洲南瓜Dof基因家族成员和探究其表达模式,从全基因组水平筛选和分析了美洲南瓜CpDof家族成员、基因结构、蛋白保守基序和亲缘关系等,同时,通过转录组和荧光定量技术比较了不同成员在美洲南瓜中的时空表达与果实发育表达特性。结果表明,在美洲南瓜中存在58个Dof基因并广泛分布于全基因组;功能域Motif 1存在于所有的Dof基因中;其启动子序列中含有光响应、激素响应和防御应激等顺式作用元件;Dof基因在美洲南瓜中具有组织表达特异性和时空表达特异性。基于转录组和荧光定量验证结果,表明CpDof7、CpDof30和CpDof42等基因与美洲南瓜356和296果实发育密切相关。以上结果为进一步克隆和解析南瓜属Dof基因功能提供了有力支撑。
关键词:美洲南瓜;DNA结合单锌指;基因家族;荧光定量PCR;果实发育
中图分类号:S642.6 文献标志码:A 文章编号:1673-2871(2023)05-016-13
Identification and expression pattern analysis of Dof gene family in Cucurbita pepo
SUN Shouru HU Deju CHANG Jingyi WANG Yong MA Changsheng ZHU Lei
(1. Henan Cucumber Crop Planting Innovation and Utilization Engineering Technology Research Center, Zhengzhou 450002, Henan, China; 2. Henan International Joint Laboratory of Horticultural Plant Biology/Henan Agricultural University, Zhengzhou 450002, Henan, China)
Abstract: DNA binding with one finger(Dof)is a transcription factor unique to plants, mainly involved in regulating plant life processes, such as growth and development, stress resistance, and quality. In order to identify the members of the Dof gene family and explore its expression pattern analysis in pumpkin (Cucurbita pepo), this study screened and analyzed the members, gene structure, protein conservative motifs and genetic relationships of CpDof family from the whole genome, and compared the spatiotemporal expression and fruit development expression characteristics of different members through transcriptome and fluorescence quantitative techniques. The results showed that there were 58 Dof genes and widely distributed all over the different chromosomes; The functional domain Motif 1 exists in all Dof genes; The promoter sequence contains cis acting elements such as light response, hormone response, and defense against stress; Dof gene has tissue expression specificity and spatiotemporal expression specificity in Cucurbita pepo. Based on the results of transcriptome and fluorescence quantitative verification, CpDof7, CpDof30 and CpDof42 genes are closely related to the fruit development of pumpkin 356 and 296. This provides strong support for further cloning and analysis of the Dof gene function in Cucurbita species.
Key words: Cucurbita pepo; DNA binding with one finger; Gene family; Fluorescence quantitative PCR; Fruit development
美洲南瓜(Cucurbita pepo)是葫蘆科南瓜属重要栽培种之一,原产于北美洲南部,并广泛分布于世界各地[1]。美洲南瓜于19世纪中叶由欧洲引入中国,并在南北方广泛栽培[2]。近些年来,随着中国设施栽培的迅速发展,美洲南瓜的种植面积逐渐扩大,目前是仅次于黄瓜的早春茬瓜类蔬菜[3]。美洲南瓜种植技术简单,适合集约化栽培与管理,且易获得高产,因此,研究和利用美洲南瓜可以为我国园艺产业的多样性和种业发展提供保障。
DNA结合单锌指(DNA binding with one finger,Dof)家族是植物所特有的转录因子家族之一,其DNA富含一个独特的单锌指结构域,所以它也被称为Dof基因家族[4]。Dof蛋白的两个主要功能性结构域是N端的DNA结合域和C端的转录调控域[5],N端是由52个高度保守的氨基酸残基组成C2-C2型单锌指结构域,并且该结构中还有4个保守的Cys残基和1个Zn2+共价结合[6]。C端的转录调控域氨基酸序列多变,保守性较低,进而导致了Dof蛋白的功能具有多样性。研究发现,如果锌指结构中的Cys残基或Zn2+离子发生改变,均会导致Dof蛋白丧失活性[7]。
Dof转录因子在植物生长发育、产量和抗性等方面均具有调控作用,是重要的明星转录因子之一。1993年,玉米的ZmDof1基因是第一个被鉴定到的Dof家族成员,它的主要功能是参与调控C4植物的光合作用[8]。目前Dof基因家族已经在拟南芥[9]、大豆[10]、白菜[11]、番茄[12]和黄瓜[13]等多个物种中被鉴定出来。Dof蛋白的功能具有多样性,主要涉及到植物的种子萌发、开花调控、非生物胁迫和生长发育等。如在拟南芥中,鉴定出来CDF1(Cycling Dof Factor1)和CDF2等多个Dof基因通过抑制CO(CONSTANS)基因表达来调控开花期[14-17];在拟南芥突变体dag1(Dof Affecting Germingation1)的种子中,发现和赤霉素合成相关的基因表达量显著上调,进一步推测该基因通过抑制赤霉素的生物合成来调控种子的萌发[18]。Dof家族成员还参与了植物对多种非生物胁迫的响应,拟南芥的AtDof5.8基因与ANAC069启动子的AAAG序列特异结合,响应高盐、干旱和ABA胁迫[19];在拟南芥中,过表达番茄的SlCDF1或SlCDF3基因,都可以提高植株的耐干旱性和耐盐胁迫性[20]。Dof蛋白也参与了植物的多种生长发育过程,过表达的OsDof12水稻株系在长日照条件下开花期显著提前[21-22],并且过表达OsDof12转基因株系不仅减少水稻主支和侧支的数目,降低植株高度,还使水稻的叶片变短、小穗长度变小,说明OsDof12参与水稻株型的建成[23]。并且有研究表明,水稻的OsDof3基因能够影响种子糊粉层中赤霉素响应的相关基因来参与调控种子萌发[24],OsDof24和OsDof25则能够调节水稻种子中贮藏蛋白谷蛋白GluB-1基因的表达[25]。在大豆中,过表达GmDof11显著增加了转基因植株的分枝数、主茎节数和百粒质量,并提高了种子含油量及大豆的产量[26-27]。以上研究结果为筛选和鉴定美洲南瓜Dof基因家族及其相关研究提供了重要思路。
美洲南瓜是重要的瓜类蔬菜作物之一,在世界各地广泛栽培,深受消费者喜爱,在瓜果蔬菜生产中也占有重要的地位。Dof基因家族在植物生长发育的各个方面和对逆境的响应过程中都有参与。然而,目前在美洲南瓜中还未有任何与Dof基因相关的研究与報道。因此,笔者以美洲南瓜为研究材料,从全基因组水平鉴定和筛选了美洲南瓜Dof基因家族成员,还分析了其在染色体上的位置、蛋白理化性质、基因结构、保守基序和进化关系等信息,同时利用转录组数据重点筛选了果实发育相关基因,并利用实时荧光定量技术进行了验证,为进一步研究美洲南瓜Dof基因家族的功能提供了参考依据。
1 材料与方法
1.1 美洲南瓜Dof基因家族成员的筛选与鉴定
从葫芦科基因组数据库(http://www.cucurbitgenomics.org/organism/14)中根据Dof基因家族的功能域,初步筛选获得了美洲南瓜的全部Dof候选基因,同时利用拟南芥的Dof成员进行了基因组数据库的同源比对分析和筛选。然后,利用在线网站NCBI的CD-Search(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)对候选Dof蛋白序列的保守结构域进行验证,将下载的文件放在软件Tbtools中进行可视化处理,剔除重复冗余及不含Dof结构域的不完整序列,最终获得美洲南瓜Dof基因家族的全部候选基因,并根据其在染色体上的位置依次命名。通过Tbtools软件中的Show Genes on Chromosome和Advanced Circos插件对Dof基因进行染色体位置和共线性的可视化分析与绘图。
1.2 基因的结构和保守基序分析及系统进化树的构建
依据美洲南瓜Dof基因家族成员的mRNA全长序列、CDS序列、氨基酸序列、外显子数量等基本数据,利用软件Tbtools中的“Protein Paramter Calc”插件分析该基因家族蛋白的理论等电点、分子质量、不稳定系数和疏水性等理化性质;利用MEME(http://meme-suite.org/tools/meme)分析基因家族成员的保守基序,其中参数设置如下:保守基序数量为15个,motif的长度为6~50 aa,最后将motif分析后的数据进行可视化处理[28]。
将葫芦科作物美洲南瓜、西瓜、甜瓜和黄瓜(http://www.cucurbitgenomics.org/)中的Dof蛋白质序列进行同源比对分析,并利用软件MEGA7中的最大似然法(maximum likehood,ML)构建葫芦科Dof蛋白的系统进化树,最后利用iTOL(https://itol.embl.de/)对构建的进化树进行美化与调整。
1.3 启动子顺式作用元件分析
从美洲南瓜基因组数据库中截取Dof基因家族每个成员的启动子序列(ATG上游2000 bp的序列),导入文本并整理成FAST文件,将该文件提交至PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)网站进行启动子顺式作用元件分析。根据分析结果,对其进行筛选排序,并利用Tbtools中的“Simple Bio Sequence Viewer”模块进行可视化处理与分析。
1.4 基于转录组的美洲南瓜Dof家族成员的表达分析
根据Aliki Xanthopoulou等[29]的转录组数据,调取Dof基因家族在美洲南瓜雌花、雄花、根、茎、种子等不同组织和授粉后不同时间(day after pollination,DAP)果实Dof的FKAM值。将提取的数据按照组织表达和果实发育时期表达进行分类,并利用软件Tbtools的“Heat Map”模块绘制Dof基因的表达热图,按照蓝色、黄色和红色对不同表达倍数进行赋值,其中,深蓝色代表表达量较低,而深红色代表表达量较高。
1.5 美洲南瓜幼果Dof基因的实时荧光定量(qRT-PCR)表达验证
美洲南瓜材料引自中国国家农作物種质资源平台—蔬菜种质资源子平台,296和356自2018年经过多代自交均培育至高代自交系,由河南农业大学园艺学院瓜类课题组保存,试验于2023年2—3月在河南农业大学瓜类课题组实验室进行。
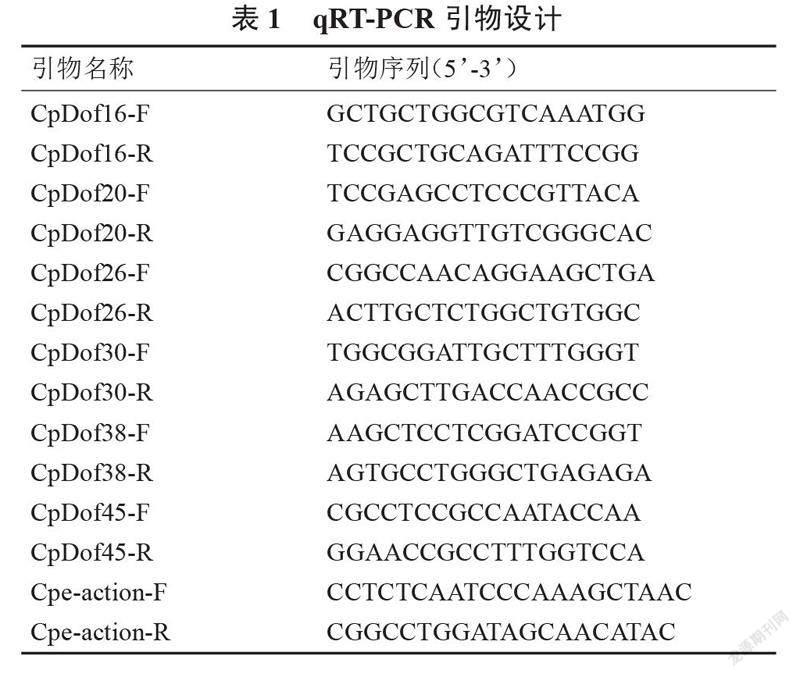
以 296和356为材料,采用随机区组法对授粉10 d后的幼果进行取样,每个样品3次重复,收集约0.5 g 的果肉用于后续 RNA 提取。样品经液氮研磨成粉末后,使用Quick RNA isolation Kit试剂盒(北京,华越洋)进行总RNA的提取。第一链cDNA采用MonScriptTM RTIII All-in-One Mix with dsDNase试剂盒(苏州,莫纳)合成。内参基因Cpe-action由河南农业大学瓜类课题组提供,利用Bio-Rad CFX96 TouchTM仪器进行实时荧光定量PCR检测。
20.0 μL Real-time PCR体系如下:1.6 μL cDNA模板,上、下游引物各0.4 μL(10 μmol·L-1)(表1),10.0 μL Taq Pro Universal SYBR qPCR Master Mix(南京,诺唯赞)和7.6 μL ddH2O。PCR程序为:95 ℃ 180 s;95 ℃ 10 s,60 ℃ 40 s,共40个循环;95 ℃ 10 s,65 ℃ 5 s,95 ℃ 5 s。每个样品3次重复。采用Office 2010 Excel对定量结果进行数据统计与分析,采用2-ΔΔCt法计算基因的相对表达量,并利用SPSS 22.0进行差异显著性分析。
2 结果与分析
2.1 美洲南瓜Dof基因家族成员的鉴定及其理化性质分析
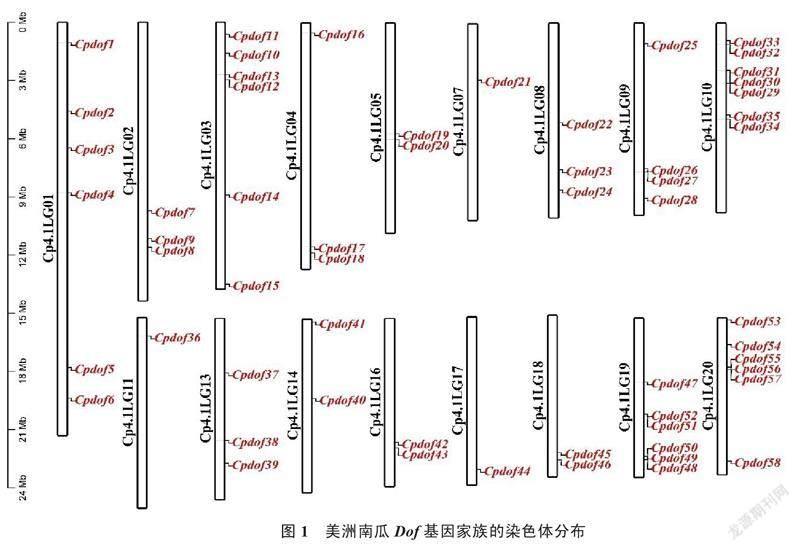
经过功能域分析和同源比对分析,初步筛选获得61个候选Dof基因。根据它们的保守结构域、基因完整性和相似性等进行剔除,最终在全基因组共鉴定到58个美洲南瓜Dof基因。通过全基因组信息的比对和分析,结果显示,除LG06、LG12和LG15外,其他染色体上均有CpDof基因家族成员的分布,这说明Dof基因家族成员在美洲南瓜全基因组上的分布较为广泛,且有可能在全基因组发挥作用。根据它们在染色体上的位置,依次命名为CpDof1~CpDof58(图1);其中,10号染色体分布的Dof家族基因最多,共7个;LG07、LG11和LG17染色体上仅检测到1个Dof基因。从图2可以看出,大部分美洲南瓜Dof基因在不同染色体上均具有较高的共线相关性,而LG06、LG12、LG15和LG17未检测到。其中,LG17上的CpDof44是全基因组唯一一个与其他染色体无任何共线性关系的Dof基因。
为了明确美洲南瓜58个Dof蛋白的理化性质,对其分子质量、等电点和亲水性等进行了计算和分析(表 2)。结果表明,美洲南瓜Dof蛋白的氨基酸数目为108~490 aa,分子质量介于12.31~53.80 kD之间,最小和最大值分别对应的是CpDof26和CpDof18;其等电点介于5.28~9.93之间,其中共有14个蛋白的等电点小于7,为酸性蛋白,43个蛋白的等电点大于7,为碱性蛋白;仅有1个为中性蛋白,对应的是CpDof25;不稳定系数为38.06~72.29,除CpDof33、CpDof51和CpDof52这3个蛋白的不稳定系数小于40,为稳定蛋白,剩余55个蛋白的不稳定系数均大于40,表明大部分CpDof为不稳定蛋白;其脂肪系数为36.67~66.39,最小值和最大值分别对应CpDof15和CpDof48;该家族成员蛋白的亲水系数均为负数,介于-0.960~-0.226之间,表明58个Dof蛋白均属于疏水性蛋白。
2.2 美洲南瓜Dof家族成员的进化树构建
为了解美洲南瓜Dof基因与其他葫芦科作物Dof基因的同源性,构建了包含美洲南瓜(Cp)、西瓜(Cl)、甜瓜(Cm)和黄瓜(Cs)4个物种的Dof蛋白质序列进化树(图3)。参考葫芦科其他物种的分组方式,将其划分为A到I共9个亚群。其中,A亚群含有的Dof蛋白数量最多,为33个;C亚群含有的CpDof蛋白最多,为12个,占比41.38%;I亚群含有Dof蛋白数量最少,共5个蛋白,其中有2个是CpDof蛋白,占比为40.00%。相较于其他瓜类,美洲南瓜的CpDof在9个亚群中均含有多个成员。
2.3 美洲南瓜Dof基因家族保守基序和基因结构分析
依据聚类和进化树构建结果(图4-a),对美洲南瓜不同亚群Dof家族成员的功能域和基因结构进行了分析。结果表明,CpDof蛋白序列中共发现15个保守基序(图4-b)。58个CpDof家族成员中均包含Motif 1(代表Dof结构域),因此,Motif 1为核心保守结构域。保守基序在蛋白/基因的功能中起着重要作用,它也是基因分类和功能分化的基础;进化树中属于同一亚群的保守基序的类型、数量和分布结构类似,表明亚群内的CpDof亲缘关系更相近。如图所示,Motif 3为C亚家族所有成员特有的基序,Motif 5为F亚家族所有成员特有的基序,由此可见,同一亚群内的成员可能在瓜类作物中有着相似的功能。
对其基因结构进行分析发现,在B、C、D和F亚家族中,多数CpDof基因含有2个外显子;在A、E、G、H和I亚家族中,多数CpDof基因只包含有CDS而无UTR(图4-c)。每个亚家族的基因结构大部分相似,但是同一亚家族中也出现少数成员的结构差异,表现为内含子与外显子的数量和长度不等。例如,D亚家族CpDof49和CpDof56内含子和外显子的数量与其他成员明显不同。另外,F亚家族的基因总碱基数相对较大(图4-c),G亚家族则较小,这说明Dof家族的不同成员在进化中存在序列的分化。
2.4 美洲南瓜Dof家族成员的启动子顺式调控元件
为了深入了解CpDof家族成员的结构和功能,对CpDof的启动子进行了顺式调控元件的分析(图5)。结果显示,大部分启动子区域均含有光响应元件,还有参与低温、伤口、防御和应激等多种胁迫响应的顺式调控元件,这说明该家族的启动子具有与胁迫相关的调控功能;同时也包含了参与水杨酸、脱落酸以及茉莉酸甲酯(MeJA)反应的顺式作用元件等。其中,CpDof4、CpDof35和CpDof46的启动子区域均含有多个脱落酸相关的顺式作用元件。
2.5 美洲南瓜Dof基因家族的表达模式分析
为了解CpDof基因家族成员的生物学功能和表达特性,利用Aliki Xanthopoulou在2021年已公布的转录组数据,分析了美洲南瓜CpDof基因在雌花、雄花、根、茎、种子等不同组织中的表达情况(图6)。结果表明,部分Dof基因如CpDof9、CpDof10、CpDof14和CpDof23等8个基因在所有的美洲南瓜组织中的表达量都比较低。其次,CpDof基因具有组织表达特异性,如在雄花中CpDof5、CpDof26和CpDof43基因的表达量明显高于雌花和其他组织;在叶柄中表达量较高的基因有CpDof42和CpDof47;在种子中CpDof40和CpDof41的表达量较高;在主根和侧根中表达量高的均集中在CpDof1、CpDof13、CpDof19等基因上。此外,部分CpDof在美洲南瓜的叶片中具有特异性表达,如CpDof39、CpDof42、CpDof54等在美洲南瓜嫩叶中的表达量高,但在老叶中的表达量显著下调;CpDof7、CpDof32和CpDof46等基因在嫩叶中表达量低,在老叶中的表达量明显上调,它们可能与叶片的衰老发育机制相关。
嫩瓜是美洲南瓜的重要产品器官,因此,重点分析了CpDof在开花授粉后2~30 DAP果实中的特异表达情况(图7):在幼果时期(2~7 DAP),CpDof31、CpDof36、CpDof49和CpDof52基因表达量相对较高,而CpDof28、CpDof45和CpDof46基因表达量相对较低。其中,CpDof16、CpDof19、CpDof26和CpDof45基因在果实发育前期(2~20 DAP)的表达量较低,但在果实发育后期(30 DAP)的表达量却明显上调;以上结果为进一步筛选和分析果实发育相关基因奠定了基础。
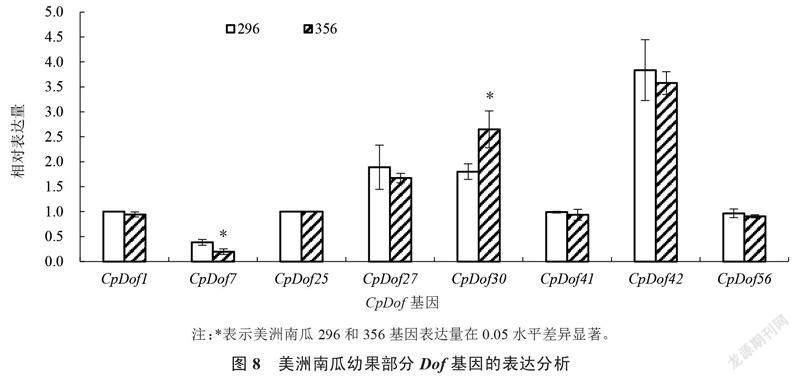
2.6 Dof基因在美洲南瓜幼果中表达特征分析
在转录组分析的基础上,对授粉后10 d的美洲南瓜高代自交系296和356幼果进行取样,通过qRT-PCR技术分析了8个Dof基因在美洲南瓜果实中的表达情况。如图8所示,CpDof27、CpDof30和CpDof42的相对表达量较高,而CpDof1、CpDof7、CpDof25、CpDof41和CpDof56等5个基因相对表达量较低。其中,相对表达量最低和最高的分别是CpDof7和CpDof42。CpDof7和CpDof30在2个不同美洲南瓜果实中的表达量差异显著,CpDof7在296中的表达量显著高于356,而基因CpDof30的表达却相反。推测这2个Dof基因可能与美洲南瓜果实的差异发育相关。自交系材料的测定结果与转录组数据相比较(图7),8个基因的表达模式基本吻合。
3 讨论与结论
笔者从美洲南瓜的基因组数据库中共鉴定到58个CpDof家族成员,该家族的成员数量与烟草相近[30]。葫芦科作物中西瓜、甜瓜和黄瓜的Dof家族成员分别为36个、34个和36个[13,31-32],而美洲南瓜的CpDof成员数约为其他葫芦科物种的2倍,这可能与南瓜属特有的全基因组染色体加倍事件有关。在美洲南瓜基因结构分析中,其家族成员间内含子和外显子差异较大,外显子数量在1~3之间。有46%的CpDof没有内含子,基因结构相对简单;这与拟南芥、水稻、小麦和黄瓜等植物的研究结果相类似[13,33-35],表明Dof基因家族的結构相对保守。同一亚群内大多数成员的保守基序分布模式相似,而笔者发现CpDof同一亚群成员的保守基序类型、数量和顺序高度相似。其中,特征功能域Motif 1存在于所有美洲南瓜CpDof中,这与拟南芥和水稻的研究结果相吻合[33]。美洲南瓜CpDof基因结构的分析将有助于更好地鉴定和分类研究其功能。基因的启动子区域响应元件分析是预测其功能的重要手段。通过对CpDof序列上游2000 bp的启动子区域进行分析,发现它们可能在激素响应和生长发育等方面发挥作用,其中,光响应元件是典型特征元件之一。前人研究表明,在拟南芥中,过表达Dof蛋白OBP3株系,通过改变其对光响应进而影响拟南芥的生长[36];拟南芥的Dof转录因子COG1通过光敏色素A(PhyA)和光敏色素B(PhyB)负调控光敏信号[5]。由此说明,Dof家族内的成员可能与光调控元件有密切关系。此外,Dof还通过参与多种激素途径,从而协同调控植物的生长发育,如烟草中Dof转录因子NtBBF1(rolb domain B factor 1)通过结合到基因rolB启动子的ACTTTA区域来调控rolB基因的组织特异性表达和生长素诱导表达[37],从而促进根的生长[38]。通过启动子顺式元件的分析将为进一步研究CpDof参与光信号和激素相关途径的潜在功能奠定基础。
通過对前人转录组数据的进一步分析,发现CpDof基因在不同组织中的表达具有特异性,在美洲南瓜果实发育的过程中,大部分CpDof基因在不同时期的表达差异较小。通过美洲南瓜自交系材料的幼果对部分CpDof基因的表达量进行验证,证明了该转录组数据的可靠性。CpDof7和CpDof42基因的相反表达模式,分别说明它们负向和正向调控了果实的早期发育。在不同的美洲南瓜材料中,CpDof7和 CpDof30的表达量呈现显著差异,推测是由种质不同或果实发育(如果形大小和果实成分等)的差异造成的。如在猕猴桃果实中发现了乙烯响应的C2H2-型锌指蛋白AdDof3,可通过转录激活AdBAM3L启动子正向调控猕猴桃果实中的淀粉含量[39]。Dof转录因子可以直接激活糖代谢和糖转运相关基因的表达,从而影响火龙果果实的糖分积累[40]。由此表明Dof对果实品质有一定的调控作用,但具体的功能机制需进一步研究。
笔者通过对美洲南瓜基因组、转录组数据和qRT-PCR分析,在全基因组水平上筛选鉴定了58个美洲南瓜Dof家族基因(CpDof1~CpDof58)。这些家族成员不但在染色体上的分布呈现聚集现象,且大部分个体成员在染色体间具有较高的相关性。美洲南瓜Dof基因的启动子序列含有光响应、激素响应和防御应激与胁迫响应等多种顺式作用元件,且同一亚族内成员间的结构和功能域高度相似。此外,美洲南瓜Dof家族基因的表达具有组织表达特异性和时空表达特异性,其中,CpDof7和CpDof30在自交系296和356幼果中的差异显著。本研究结果为深入挖掘美洲南瓜Dof基因及其作用机制研究提供了理论依据。
参考文献
[1] 李金峰,刘佳,侯敏,等.美洲南瓜枯萎病菌分离鉴定及其生物学特性[J].中国瓜菜,2020,33(3):23-28.
[2] 朱海生,刘建汀,温文旭,等.美洲南瓜α-微管蛋白CpTUA基因的分离及其作为内参基因的应用[J].中国细胞生物学学报,2018,40(12):2040-2050.
[3] 王朝阳,程永安,张恩慧,等.西葫芦花蕾纵横径长度与小孢子发育关系研究[J].西北农林科技大学学报(自然科学版),2012,40(5):92-98.
[4] 张华珍,吴昊,李香花,等.一个低氮诱导表达的水稻Dof转录因子OsDof-13的分离和转化[J].分子植物育种,2007,5(4):455-460.
[5] DIAZ I,VICENTE-CARBAJOSA J,ABRAHAM Z,et al.The GAMYB protein from barley interacts with the DOF transcription factor BPBF and activates endosperm-specific genes during seed development[J].The Plant Journal,2002,29(4):453-464.
[6] YANAGISAWA S.Dof domain proteins:plant-specific transcription factors associated with diverse phenomena unique to plants[J].Plant & Cell Physiology,2004,45(4):386-91.
[7] CAVALAR M,MOLLER C,OFFERMANN S,et al.The interaction of DOF transcription factors with nucleosomes depends on the positioning of the binding site and is facilitated by maize HMGB5[J].Biochemistry,2003,42(7):2149-2157.
[8] YANAGISAWA S,IZUI K.Molecular cloning of two DNA-binding proteins of maize that are structurally different but interact with the same sequence motif[J].Journal of Biological Chemistry,1993,268(21):16028-16036.
[9] LIJAVETZKY D,CARBONERO P,VICENTE-CARBAJOSA J,et al.Genome-wide comparative phylogenetic analysis of the rice and Arabidopsis Dof gene families[J].BMC Evolutionary Biology,2003,3(1):17.
[10] GUO Y ,QIU L.Genome wide analysis of the Dof transcription factor gene family reveals soybean-specific duplicable and functional characteristics[J].PLoS One,2013,8(9):e76809.
[11] MA J,LI M Y,WANG F,et al.Genome-wide analysis of Dof family transcription factors and their responses to abiotic stresses in Chinese cabbage[J].BMC Genomics,2015,16(1):33.
[12] CAI X F,ZHANG Y Y,ZHANG J H,et al.Advances in research on function of the Dof gene family in plant[J].Plant Physiology Journal,2013,49(1):1-12.
[13] WEN C L,CHENG Q,ZHAO L Q,et al.Identification and characterisation of Dof transcription factors in the cucumber genome[J].Scientific Reports,2016,6:23072.
[14] IMAIZUMI T,SCHULTZ T F,HARMON F G ,et al.FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis[J].Science,2005,309(5732):293-297.
[15] IMAIZUMI T,KAY S A.Photoperiodic control of flowering:not only by coincidence[J].Trends in Plant Science,2006,11(11):550-558.
[16] FORNARA F,PANIGRAHI K C S,GISSOT L,et al.Arabidopsis DOF transcription factors act redundantly to reduce CONSTANS expression and are essential for a photoperiodic flowering response[J].Developmental Cell,2009,17(1):75-86.
[17] SONG Y H,SMITH R W,TO B J,et al.FKF1 conveys timing information for CONSTANS stabilization in photoperiodic flowering[J].Science,2012,336(6084):1045-1049.
[18] GABRIELE S.The Dof protein DAG1 mediates PIL5 activity on seed germination by negatively regulating GA biosynthetic gene AtGA3ox1[J].The Plant Journal,2010,61(2):312-323.
[19] HE L,SU C,WANG Y C,et al.ATDOF5.8 protein is the upstream regulator of ANAC069 and is responsive to abiotic stress[J].Biochimie,2015,110:17-24.
[20] CORRALES A R,NEBAUER S G,CARRILLO L,et al.Characterization of tomato cycling Dof factors reveals conserved and new functions in the control of flowering time and abiotic stress responses[J].Journal of Experimental Botany,2014,65(4):995-1012.
[21] IWAMOTO M,HIGO K,TAKANO M.Circadian clock- and phytochrome-regulated Dof-like gene,Rdd1,is associated with grain size in rice[J].Plant Cell & Environment,2010,32(5):592-603.
[22] LI D J,YANG C H,LI X B,et al.Functional characterization of rice OsDof12.[J].Planta,2009,229(6):1159-1169.
[23] QI W,LI D,LI D,et al.Overexpression of OsDof12 affects plant architecture in rice (Oryza sativa L.)[J].Frontiers in Plant Science,2015,6:833.
[24] WASHIO K.Functional dissections between GAMYB and Dof transcription factors suggest a role for protein-protein associations in the gibberellin-mediated expression of the RAmy1A gene in the rice aleurone[J].Plant Physiology,2003,133(2):850-863.
[25] ZHANG Y.Functional analysis of Dof transcription factors controlling heading date and PPDK gene expression in rice[D].Holland:Leiden University,2015.
[26] 張雪,尹悦佳,范贝,等.植物Dof转录因子的结构特点及功能研究进展[J].作物杂志,2016(2):14-20.
[27] 王志坤,SEBASTIAN A,常健敏,等.转GmDof11基因高油转基因大豆的鉴定及主要农艺性状调查[J].作物杂志,2014(2):39-42.
[28] SAITOU N,NEI M.The neighbor-joining method:a new method for reconstructing phylogenetic trees[J].Molecular Biology & Evolution,1987(4):406.
[29] XANTHOPOULOU A,MONTERO-PAU J,BELEN P,et al.A comprehensive RNA-Seq-based gene expression atlas of the summer squash (Cucurbita pepo) provides insights into fruit morphology and ripening mechanisms[J]. BMC Genomics,2021,22(1):341.
[30] 赵振祥,崔露莹,周东波,等.普通烟草Dof转录因子家族的全基因组鉴定及分析[J].中国烟草科学,2022,43(4):70-78.
[31] ZHOU Y,CHENG Y,WAN C,et al.Genome-wide characterization and expression analysis of the Dof gene family related to abiotic stress in watermelon[J].Peer Journal,2020,8(1/2):e8358.
[32] 张焕欣,李国权,杨惠栋,等.甜瓜Dof家族全基因组鉴定与表达分析[J].园艺学报,2019,46(11):2176-2187.
[33] 葛敏,吕远大,李坦,等.玉米Dof转录因子家族的全基因组鉴定与分析[J].中国农业科学,2014,47(23):4563-4572.
[34] LIU Y,LIU N N,DENG X,et al.Genome-wide analysis of wheat DNA-binding with one finger (Dof) transcription factor genes:evolutionary characteristics and diverse abiotic stress responses[J].BMC Genomics,2020,21(1):549-562.
[35] 牟藝菲.小麦OPR和LOX基因家族的鉴定及其抗逆功能分析[D].陕西杨凌:西北农林科技大学, 2019.
[36] UMEMURA Y,ISHIDUKA T,YAMAMOTO R,et al.The Dof domain,a zinc finger DNA-binding domain conserved only in higher plants,truly functions as a Cys2/Cys2 Zn finger domain[J].The Plant Journal for Cell and Molecular Biology,2004,37(5):741-749.
[37] WARD J M,CUFR C A,DENZEL M A.The Dof transcription factor OBP3 modulates phytochrome and cryptochrome signaling in Arabidopsis[J].The Plant Cell,2005,17(2):475-485.
[38] PARK D H,LIM P O,KIM J S,et al.The Arabidopsis COG1 gene encodes a Dof domain transcription factor and negatively regulates phytochrome signaling[J].The Plant Journal,2003,34(2):161-171.
[39] 张爱迪.AdDof3转录调控采后猕猴桃果实淀粉降解[D].杭州:浙江大学,2018.
[40] 曾闰曦.Dof转录因子调控火龙果果实糖代谢及转运的分子机制[D].武汉:华南农业大学,2020.
猜你喜欢
中国现代医生(2017年28期)2017-11-17 21:50:10
肉类研究(2017年8期)2017-11-16 12:06:16
中国医学创新(2017年25期)2017-11-14 13:57:39
安徽农学通报(2017年19期)2017-11-08 13:08:23
医学信息(2017年20期)2017-10-10 16:15:35
上海预防医学(2017年4期)2017-05-18 17:11:50
中国现代医生(2017年2期)2017-04-06 14:02:38
江苏农业科学(2016年11期)2017-03-21 08:53:24
今日健康(2016年12期)2016-11-17 19:21:34
江苏农业科学(2016年2期)2016-04-11 15:38:48
