
基于转录组学和代谢组学解析氮调控花生结瘤固氮的机理
2023-05-27 22:32:09刁瑞宁杨莎张佳蕾王建国彭振英崔利刘珂珂李新国万书波
山东农业科学 2023年4期
刁瑞宁 杨莎 张佳蕾 王建国 彭振英 崔利 刘珂珂 李新国 万书波


关键词:花生;结瘤;固氮酶活性;转录组学;代谢组学
花生作为一种重要的油料作物,在热带、亚热带和温带地区广泛种植。近年来,为了确保花生高产,农民在生产过程中往往过量施用氮肥,导致氮肥供应与作物需求严重不同步,不仅造成资源浪费,而且污染环境,严重威胁我国农业生产的可持续发展。
氮素是作物生长发育必需的三大营养元素之一,与作物的产量和品质密切相关。豆科作物通过与土壤根瘤菌相互作用形成一个特殊器官——根瘤,表现出较强的固氮能力,对其生长和产量形成至关重要,也是可持续农业生产的有效氮源。然而,豆科作物的生物固氮受土壤中氮素有效性的严格调控,当氮素过多时对结瘤和固氮具有强烈的抑制作用,形成“氮阻遏”效应。
氮素对豆科作物结瘤固氮的调控是一个复杂的过程。结瘤是一个耗能的过程,过多的根瘤同样不利于作物生长发育,因此豆科作物通过AON(autoregulation of nodulation)途径负反馈调节根瘤数量,减少能量消耗,维持碳氮平衡,而氮素可以通过AON途径抑制豆科作物结瘤。研究表明,氮素供给促进了豆科作物根系生长及对氮的吸收和同化,增加了碳消耗,因此减少了光合同化产物向根瘤的分配,从而抑制了根瘤生长发育及固氮酶活性。根瘤的硝酸盐代谢也是硝态氮抑制根瘤固氮的重要原因。研究表明,硝态氮能提高根瘤中硝酸还原酶和亚硝酸还原酶活性,硝酸还原酶能将硝酸盐还原为亚硝酸盐并在胞质中积累,而亚硝酸盐是类菌体固氮酶活性的抑制剂,因此积累的亚硝酸盐会直接抑制固氮酶活性;Stephens等研究也认为添加KNO3或KNO2明显抑制根瘤的固氮酶活性。
目前在氮素调控豆科作物结瘤和固氮的分子机制方面已经取得了一定的进展,但大多数研究是以大豆、百脉根、蒺藜、苜蓿等模式作物为试验材料,关于花生的研究较少,而且不同的豆科作物中调控结瘤固氮的分子机制存在差异,因此本试验开展关于氮素调控花生结瘤固氮机理的研究,以期为高效利用花生生物固氮体系提供理论依据。
1材料与方法
1.1供试材料
试验于2019-2020年在山东省农业科学院饮马泉试验基地进行,供试花生品种为花育22号。采用盆栽试验,花盆规格为高30cm、直径25cm,栽培基质为石英砂与蛭石按3:1(体积比)比例混合。
1.2试验设计
试验设CK(不施氮)和N(施纯氮90kg/hm2)两个处理,每个处理30盆,随机排列,重复3次。花生于5月10日播种,播种前用活性良好的根瘤菌菌液拌种,每盆播3穴,每穴1粒,出苗后选留2株壮苗;施底肥,氮肥按试验设计,每盆施尿素1.74g,其他肥料按Ca0 150kg/hm2、P 180 kg/hm2、K180kg/hm2施用,每盆分别施Ca0 1.33g、KH2P044.62g。所有盆栽均放在可移动大棚下进行避雨,定期定量浇水。
1.3测定指标与方法
1.3.1根瘤数量统计出苗后35天取样,每个处理随机取样5盆。将取样植株根系清洗干净,统计每株花生的根瘤数量。
1.3.2固氮酶活性测定出苗后35天取样,每个处理随机取3盆花生。将根瘤摘下洗干净并擦干表面水分,装入广口瓶并盖上带孔的胶塞,连接好玻璃导管及橡胶导管,密封;用注射器从密闭的广口瓶中抽出10mL空气,然后注入10mL乙炔气体,室温放置2h;抽取混合气体,用气象色谱仪测定乙烯峰面积,固氮酶活性用每克根瘤每小时产生的乙烯量(nmol)表示。
1.3.3转录组学分析
出苗后35天取样,每个处理随机取样3盆。取样后将根系用蒸馏水冲洗干净,擦干水分,剪碎混匀后用锡箔纸包好,迅速放人液氮。利用TRIzol法提取RNA,经过质量检测后利用试剂盒(NEBNext⑧UltrarM RNA Li-brary Prep Kit for Illumina,NEB,美国)进行文库构建,主要步骤参考说明书。建好的文库应用Il一lumina HiSeq平台进行测序,整個测序过程由武汉迈特维尔生物科技有限公司完成。
高通量测序后的原始序列经过过滤、筛选,获得可以用于后续分析的Clean Reads。利用HI-SAT2将Clean Reads与四倍体栽培花生参考基因组进行序列比对,然后以log2FC≥1且FDR<0.05为条件对获得的基因进行筛选,得到差异表达基因。参照GO(Gene Ontology)数据库将差异表达基因进行功能注释分类,参考KEGG(KyotoEncyclopedia of Genes and Genomes)数据库进一步分析差异表达基因参与的代谢途径。
1.3.4代谢组学分析代谢组学分析材料同转录组学测序材料,测序分析由武汉迈特维尔生物科技有限公司完成。将花育22号根系(含根瘤)粉碎并研磨至粉末状,称取100mg溶解于1.2mL70%甲醇提取液中,放人4℃冰箱过夜,离心(转速12000r/min,10min)后取上清液,过0.22um微孔滤膜,用超高效液相色谱(UPLC)和串联质谱(MS/MS)分析仪检测代谢物,然后基于MWDB(Metware Database),根据二级谱信息进行物质定性,并利用三重四级杆质谱的多反应检测(multi-ple reaction monitoring,MRM)模式进行定量分析。利用软件Analyst 1.6.3处理得到的质谱数据,用MultiaQuant软件进行色谱峰的积分和校正;然后进行主成分分析和聚类分析,再以VIP≥1、log2FC≥1为条件筛选出差异代谢物,其中VIP为变量重要性投影(variable importance in projec-tion)。最终利用KEGG数据库和GO数据库对差异代谢物进行注释,获得代谢物参与的代谢路径。
1.3.5qRT-PCR验证取1.3.3中提取的RNA,利用试剂盒进行反转录,然后进行定量PCR试验。每个样品设置3次生物学重复。所用引物如表1所示。PCR反应体系:SYBR Green MasterMix10uL,引物F、R各0.5uL,cDNAluL,用RNase Free H20补至20uL。具体操作按照荧光定量PCR说明书进行。以actin作为内参基因,采用2-△△法对qRT-PCR结果进行计算,用t—test对数据进行显著性分析。
1.3.6生长素类化合物含量测定出苗后35天取样,每个处理随机取样3盆。取样后将根系用蒸馏水冲洗干净,擦干水分,剪碎混匀后用锡箔纸包好,迅速放人液氮速冻。由武汉迈特维尔生物科技有限公司(http://www. metware. cn/)基于LC-MS/MS平台对样品进行生长素类化合物含量的测定。
1.4数据处理与统计分析
基因相对表达量、根瘤数与固氮酶活性数据用SPSS22.0软件统计分析,用Microsoft Excel2016软件制图。
2结果与分析
2.1转录组测序数据质量评估
经分析(表2)发现,每个样品产生44. 03~52.29M的Raw Reads,去除低质量数据后得到42.32~51.19M的Clean Reads,GC含量在43.87%~44.29%之间,没有AT/GC分离现象,Q20和Q30的碱基比分别达到97%和93%以上,充分证明样本测序数据的质量较好。将得到的Clean Reads进一步与栽培花生基因组进行比对,质控后的测序数据中94%以上的序列可以比对到参考基因组中,79%以上的序列被定位到唯一的区域,由此说明,测序数据的比对效果较好,可以进行后续分析。
2.2样本间基因表达差异分析
本试验中,以log2FC≥1且FDR<0.05为条件筛选对照(CK)和氮处理(N)中的基因,如图1A所示。经统计,共获得3123个差异表达基因,其中上调表达的有1538个,下调表达的有1585个(图1B)。
经具体分析,从中筛选出13个与氮调控花生结瘤固氮相关的基因(表3),包括编码生长素信号转导(生长素应答因子)、鸟氨酸脱羧酶、天冬酰胺合成酶、谷氨酰胺合成酶、谷氨酸合酶以及苹果酸脱氢酶等基因:施氮后,2个编码生长素信号转导的基因、2个鸟氨酸脱羧酶基因、2个天冬酰胺合成酶基因上调表达,其余7个基因均下调表达。
2.3转录组测序结果的qRT-PCR验证
为了验证转录组信息的可信度,随机选取6个差异表达基因进行qRT-PCR验证,结果(图2)显示,N处理后LOC112715771上调表达,其余5个基因均下调表达,与RNA-seq分析结果一致,证明转录组测序结果可靠。
2.4差异表达基因GO功能注释和KEGG富集分析
对得到的差异基因进行GO功能注释,可以阐明样本差异在基因功能上的体现。本试验对CK与N处理中的差异基因进行GO功能注释,可以从宏观上展示花生根部响应氮素处理的基因的功能分布特征。基于前期筛选出来的差异基因,选取了富集分析结果中q value最低的50个GO-Term,绘制富集条目直方图(图3)。差异基因的功能注释为3大类:生物过程(biologicalprocess)、细胞组成(cellular component)和分子功能(molecular function)。N处理中的差异基因主要参与的生物过程有:细胞过程(cellularprocess)、代谢过程(metabolic process)、對刺激的响应(response to stimulus)、生物调节(biologicalregulation)等;在细胞组成中,差异基因主要富集在细胞组分(cell part)、细胞器(organelle)等;在分子功能中,催化活性(catalytic activity)、转录调节活性(transcription regulator activity)等所占比例较高。
生物体内的不同基因产物通过相互作用行使生物学功能,对差异表达基因的通路注释分析有助于进一步解读基因的功能,因此,本研究对CK与N处理的差异基因进行了KEGG通路富集分析。我们选取了差异表达基因富集的前20条KEGG通路进行展示,从图4可以看出,N处理中差异基因富集的主要通路有次生代谢物的生物合成(biosynthesis ofsecondary metabolites)、植物激素信号转导(plant hor-mone signal transduction)、苯丙素的生物合成(pheny-lpropanoid biosynthesis) .MAPK信号途径(MAPK sig-naling pathway-plant)、甘油磷脂代谢(glycerophospho-lipid metabolism)、氮代谢(nitrogen metabolism)等。
2.5样本中差异代谢物的筛选
本试验以log2FC≥1且VIP≥1为条件,对CK与N处理中检测到的代谢物进行筛选,得到201种差异代谢物,其中87种代谢物下调,114种代谢物上调。这些差异代谢物中有32种氨基酸及其衍生物(amino acids and derivatives)、24种酚酸(phenolic acids)、10种核苷酸及其衍生物(nu-cleotides
and
derivatives)、40种黄酮类(fla-vonoids)、2种醌类(quinones)、7种木脂素和香豆素(lignans
and coumarins)、27种其他类(others)、1种鞣质(tannins)、14种生物碱(alkaloids)、4种萜类(terpenoids)、11种有机酸(organic acids)、29种脂质(lipids),其中,黄酮类代谢物所占比重最高,约为19.9%,除2种代谢物下调外,38种黄酮类代谢物上调(图5)。综上所述,氮处理后,黄酮、氨基酸、酚酸、木脂素和香豆素等代谢物在花生根和根瘤中呈上调趋势,而脂质、核苷酸和有机酸等代谢物则呈下调趋势。
2.6转录组学和代谢组学联合分析
为了进一步了解同一生物过程的代谢物与基因之间的关联,进行代谢组学和转录组学联合分析,共得到60条显著富集的KEGG通路(图6)。差异基因主要富集的KEGG通路包括苯丙素生物合成(phenylpropanoid biosynthesis)、植物激素信号转导(plant hormone signal transduction)、次生代谢物的生物合成(biosynthesis of secondary me-tabolites)、甘油磷脂代谢(glycerophospholipid me-tabolism)、氮代谢(nitrogen metabolism)等;差异代谢物主要富集的KEGG通路包括异黄酮生物合成(isoflavonoid biosynthesis),次生代谢物生物合成(biosynthesis of secondary metabolites),萜、哌啶和吡啶生物碱的生物合成(tropane,piperidine andpyridine alkaloid biosynthesis),精氨酸生物合成(arginine biosynthesis),氨基酸生物合成(biosyn-thesis of amino acids)等。
2.7氮处理对花生根系生长素类化合物含量的影响
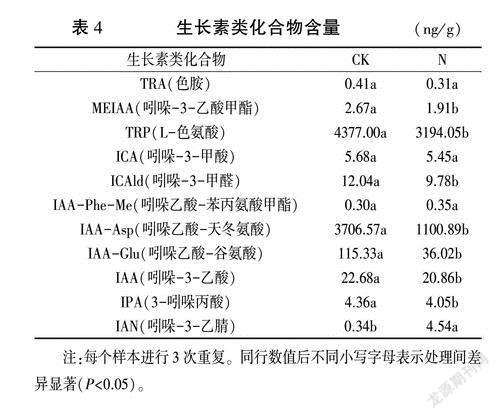
前期试验中发现生长素及其信号转导参与花生结瘤过程,因此对氮素处理后花生根部生长素类化合物及其含量进行了测定,经LC-MS/MS分析共筛选出11种生长素类化合物(表4),N处理后,TRA、ICA含量降低,但差异不显著;IAA -Phe-Me和IAN含量增加,IAN增加显著;其余7种化合物含量降低且差异显著。
2.8氮处理对花生根瘤数和根瘤固氮酶活性的影响
与对照相比,氮处理条件下花育22号的根瘤数量显著减少(图7A),根瘤的固氮酶活性也显著降低(图7B),表明施氮会抑制花生结瘤和固氮
3讨论
花生能通过根瘤与根瘤菌共生固氮,但施氮量过高时会抑制其固氮效应。为研究氮素调控花生结瘤固氮机理,本研究对施氮和不施氮花生根系进行转录组分析,鉴定出3123个差异表达基因,富集在生物过程、细胞组成和分子功能三大类,其中与氮调控根瘤固氮有关的基因13个;经液相色谱串联质谱法分析,鉴定出酚酸、黄酮、氨基酸等12大类201种差异代谢物,其中黄酮类代谢物占比最高,约占19.9%。通过转录组学和代谢组学的联合分析,进一步探究了氮素调控花生结瘤固氮的主要途径,具体结果如下。
3.1氮素对生长素信号转导途径的调控
生长素在植物整个生命周期中发挥重要作用,能通过调控生长素响应基因的表达参与根瘤的形成。在生长素信号转导的早期阶段Aux/IAA(auxin/indole-3-aceticacid)家方笑、ARF(auxin re-sponse factor)家族、SAUR(small auxin upregulatedRNA)家族和生长素响应GH3(gretchenhagen 3)家族对生长素的刺激有响应。本研究中发现,氮处理后编码生长素信号转导(生长素应答蛋白IAA27)的基因表达量上调。因此,外源氮会使花生根中IAA基因表达量增加,抑制生长素信号转导,这一研究结果与在苜蓿中的研究结果相反,可能是因为苜蓿与花生的根瘤类型不同。另外,外施氮肥后3个ARF基因表达下调,而ARF蛋白对根瘤菌侵染具有直接反应,因此,ARF基因表达下调可能会影响根瘤菌侵染。综上所述,外施氮肥会通过影响生长素响应基因AuxIAA和ARF的表达来影响生长素信号转导和根瘤菌侵染,进而影响花生结瘤。
3.2氮素对根瘤固氮信号分子的调控
有报道称,腐胺、天冬酰胺是调节根瘤固氮的信号分子。鹰嘴豆和黑豆根瘤中腐胺的浓度与固氮能力呈正相关,鸟氨酸脱羧酶(ODC)在植物中催化精氨酸合成腐胺。本研究发现2个编码鸟氨酸脱羧酶的基因在氮处理中表达量上升。因此,外源氮促进花生根瘤中精氨酸向腐胺的转化,降低了花生根瘤中固氮酶活性,这与鹰嘴豆、豌豆和黑豆的研究结果相反,但与大豆的研究结果相似,表明腐胺介导的固氮酶活性调节可能与作物种类有关。Bacanamwo和Har-per的研究表明大豆植株的固氮酶活性与天冬酰胺及其代谢物(天冬氨酸和谷氨酸)的含量有关,施用外源氮后,根瘤中谷氨酸和天冬氨酸的含量会增加。本研究中,外施氮肥后编码天冬酰胺合成酶(AS)的基因表达上调,可能会使天冬酰胺含量增加,而天冬酰胺负调控固氮活性。综上所述,外施氮肥促进了根瘤中腐胺和天冬酰胺等信号分子的合成,这会在一定程度上抑制根瘤的固氮酶活性。
3.3氮素对根瘤固氮能量的调控
豆科作物与根瘤菌的交互需要根瘤菌与寄主植物之间的协调沟通,这种沟通使根瘤菌能够被容纳在根瘤中,在此处大气中的N,被还原为氨,为植物提供了一种可以被同化的氮源:反过来,植物也向根瘤菌提供固定碳。在定型根瘤中,大部分固定的NH3通过类菌体和共生体擴散到植物细胞质中,通过谷氨酰胺合成酶/谷氨酸合成酶(GS/GOGAT)循环将其迅速同化为谷氨酰胺(Gln),然后谷氨酰胺被用作氮供体,在定型根瘤中用于嘌呤和脲类的生物合成。本试验中,氮处理后编码GS和GOGAT的基因表达下调,这可能会直接降低根瘤的固氮作用。
根瘤的碳代谢是根瘤固氮的主要能量来源。蔗糖可通过淀粉合酶转化为淀粉用于贮存,并通过蔗糖合酶催化为葡萄糖和果糖,进一步参与糖酵解等代谢途径。草酰乙酸可以通过苹果酸脱氢酶转化成苹果酸,苹果酸通过转运蛋白进入共生体中,从而维持共生体正常的呼吸和共生固氮。Tesfaye等研究表明,苜蓿中苹果酸脱氢酶过表达可导致苜蓿根瘤中苹果酸浓度升高。本研究中,施氮花生根系中编码苹果酸脱氢酶和NAD依赖的苹果酸酶基因表达量下调,这可能会使根瘤中苹果酸浓度降低。因此,外源氮对花生根瘤固氮的抑制作用可能是由于苹果酸浓度降低,根瘤菌固氮能量不足所致。
4结论
本研究基于转录组学和代谢组学综合分析了施氮对花生结瘤固氮的调控,发现氮素通过调控生长素响应基因的表达来影响生长素信号转导,进而影响花生结瘤:花生根瘤中腐胺、天冬酰胺等物质的合成可能与氮素抑制固氮酶活性有一定的相关性:而花生根瘤中苹果酸浓度降低,根瘤固氮能量不足可能是氮素抑制固氮酶活性的另一原因。因此,花生种植过程中要合理施用氮肥,提高花生与根瘤菌的共生固氮效率。
猜你喜欢
儿童时代·快乐苗苗(2022年10期)2022-12-09 08:53:44
山东冶金(2022年1期)2022-04-19 13:40:14
上海金属(2021年4期)2021-07-28 12:33:22
上海金属(2020年6期)2021-01-04 12:25:52
中国中药杂志(2016年21期)2017-02-16 14:10:12
中国中药杂志(2016年21期)2017-02-16 13:04:42
中国中药杂志(2016年22期)2017-02-13 16:45:32
儿童时代·快乐苗苗(2016年4期)2016-11-07 01:09:26
学苑创造·A版(2016年7期)2016-07-06 18:11:05
儿童时代·快乐苗苗(2015年8期)2015-11-12 23:16:31
