
高温干旱下缩节胺通过调节碳和氨基酸代谢提高Bt棉杀虫蛋白含量的生理机制
2023-05-15 08:52邢羽桐滕永康吴天凡刘媛媛陈源陈媛陈德华张祥
中国农业科学 2023年8期
邢羽桐,滕永康,吴天凡,刘媛媛,陈源,陈媛,陈德华,张祥
高温干旱下缩节胺通过调节碳和氨基酸代谢提高棉杀虫蛋白含量的生理机制

扬州大学农学院/江苏省作物遗传生理国家重点实验室培育点,江苏扬州 225009
【目的】探讨高温干旱胁迫下缩节胺(mepiquat chloride,DPC)调控()棉杀虫蛋白含量的生理机制,为高抗虫性棉品种选育及高产高效栽培提供理论依据。【方法】2020—2021年以转抗虫基因抗虫棉品种泗抗3号为材料,采用盆栽法,在人工气候室进行高温干旱胁迫,胁迫开始后立即用20 mg·L-1DPC和清水(对照)喷施。7 d后测定铃壳杀虫蛋白含量、α-酮戊二酸含量、丙酮酸含量以及谷氨酸合酶活性、谷氨酸草酰乙酸转氨酶活性、可溶性蛋白含量、游离氨基酸含量。并进行转录组测序,利用DESeq进行差异基因分析,通过GO富集和KEGG Pathway数据库注释参与DPC调节杀虫蛋白含量的差异表达基因。【结果】与清水对照相比,DPC可显著提高高温干旱条件下棉铃壳中杀虫蛋白含量,提高幅度达4.7%—11.9%。在碳代谢方面,α-酮戊二酸含量、丙酮酸含量提高46%—57%和25%—29%;在氨基酸代谢方面,谷氨酸合酶活性、谷氨酸草酰乙酸转氨酶活性、可溶性蛋白含量、游离氨基酸含量分别提高32%—44%、30%—40%、28%和22%—27%。转录组分析结果表明,DPC处理后上调基因7 542条,下调基因10 449条。GO和KEGG结果显示,差异基因主要涉及氨基酸代谢、碳代谢等生物过程。其中编码6-磷酸果糖激酶和丙酮酸激酶、谷氨酸丙酮酸转氨酶、丙酮酸脱氢酶、柠檬酸合酶、异柠檬酸脱氢酶和α-酮戊二酸脱氢酶、谷氨酸合酶、吡咯啉-5-羧酸脱氢酶、谷氨酸草酰乙酸转氨酶、N-乙酰谷氨酸合成酶、乙酰鸟氨酸脱乙酰酶基因表达显著上调。【结论】高温干旱下,DPC通过调节碳、氨基酸代谢增加丙氨酸、α-酮戊二酸含量,提高天冬氨酸、谷氨酸、丙酮酸和精氨酸合成能力,进而提高棉杀虫蛋白含量。
棉;缩节胺;杀虫蛋白;氨基酸代谢;碳代谢
0 引言
【研究意义】转()基因棉花(棉)是一种将苏云金芽孢杆菌中杀虫蛋白基因,经过改造后转入棉花体内,从而使棉株表达杀虫蛋白,防治鳞翅目害虫的一种棉花。并且杀虫蛋白含量高低决定棉的抗虫效果。因此,种植棉可减少棉田农药的使用,产生良好的经济效益和生态效益。【前人研究进展】研究人员发现在棉生长过程中植株各器官尤其是生殖器官杀虫蛋白表达易受外部不良环境的影响,从而降低了其应用效果。在全球棉花主产国,棉花花铃期常出现极端高温与土壤干旱现象,造成棉杀虫蛋白含量下降和抗虫性显著下降。如夏兰芹等[1]发现高温可导致棉杀虫蛋白含量急剧降低。CHEN等[2]和WANG等[3]则进一步明确37℃和38℃分别是导致叶片和棉蕾中杀虫蛋白表达量显著下降的临界温度。BENEDICT等[4]和SACHS等[5]发现当降水量较少时土壤含水量降低,土壤水分压力会降低植株中杀虫蛋白的含量。MARTINS等[6]和ROCHESTER[7]研究发现干旱胁迫导致棉叶杀虫蛋白表达下降,并最终引起棉铃虫分布发生变化。在棉杀虫蛋白表达调节方面,近年来有研究表明,喷施生长调节剂可提高棉杀虫蛋白含量。董志强等[8]研究发现缩节胺(mepiquat chloride,DPC)能提高细胞膜的稳定性,增加植株抗逆性。郑青松等[9]研究发现DPC有利于缓解盐渍对棉苗生长的抑制。董志强等[8]则研究发现棉花专用生长调节剂B1P1S1(油菜素内酯、DPC、水杨酸各1份)能提高棉盛蕾期上部叶片、花铃期下部叶片和果枝叶中杀虫蛋白含量;ZHANG等[10]进一步证实了单施DPC亦可显著调节杀虫蛋白含量。但目前,DPC对高温与干旱互作胁迫下棉杀虫蛋白含量的影响及相关生理机制还未见系统报道。【本研究切入点】本文将以高温干旱逆境下棉植株为材料,应用转录组分析技术和相关生理测定分析,来明晰DPC对逆境下杀虫蛋白表达的调节效应及其机制。【拟解决的关键问题】利用转录组数据和相关生理数据,分析外源调节剂DPC对高温干旱逆境下棉杀虫蛋白表达的影响及其生理机制,从而获得提升逆境下棉自身抗虫性的调节技术,缓解逆境对抗虫性的负面影响,进而减少棉田农药用量。研究结果对于基因育种提高棉自身杀虫蛋白稳定性具有重要意义,同时也可为生产上抗虫棉的安全应用提供技术指导和理论依据。
1 材料与方法
1.1 试验材料与设计
试验于2020—2021年在扬州大学农学院试验温室进行。以生产上应用较广的转基因抗虫棉常规品种泗抗3号(以下简称为SK-3)为材料。采用盆栽试验,所用盆钵直径40 cm,盆高50 cm,每盆装25 kg过筛的砂壤土(取自大田试验田)。采用育苗移栽方式种植,两年均于4月6日播种,出苗41 d后,选择长势一致的壮苗移栽至盆钵中,每盆1株。其他生长管理措施按大田高产栽培要求进行。
盛花期(7月20日)控制浇水,并标记植株中部果枝内围当日花。7月30日,在人工气候室进行高温干旱胁迫,其中白天(7:00—19:00)温度设为38℃,夜间(19:00—7:00)设为28℃。土壤含水量为最大持水量的50%。于早晨、中午、傍晚使用WET土壤三参数速测仪监测土壤水分,用称重法控制土壤水分,即当监测发现土壤水分低于设计值时,进行定量补水,达到预期水分指标。开始胁迫后立即用 20 mg·L-1DPC和清水(Water)喷施植株。持续胁迫7 d,胁迫结束后,取棉株相同部位棉铃,带回实验室立即用无菌刀将棉花分为铃壳、棉籽、纤维等三部分,分别液氮冷冻后在-80℃条件下保存,待测。
1.2 测定内容和方法
1.2.1 杀虫蛋白含量 应用酶联免疫法(ELISA)测定,试剂盒由中国农业大学提供。测定方法参见文献[11]。
1.2.2 碳、氨基酸代谢相关物质含量 采用文献[12]的方法测定谷氨酸合酶活性、谷氨酸草酰乙酸转氨酶活性、可溶性蛋白含量、游离氨基酸含量。按照Hodges[13]的方法测定α-酮戊二酸的含量。按照Mustroph等[14]的方法测定丙酮酸含量。
1.2.3 铃壳转录组测序
1.2.3.1 cDNA文库的构建及转录组测序 处理7 d后取标记棉铃铃壳用于cDNA文库构建和转录组测序。样品cDNA由上海鹿明生物科技有限公司制备,然后进行转录组测序。所得原始数据经过滤得到有效数据,利用hisat2[15]比对到棉花基因组得到总映射和唯一映射。
1.2.3.2 差异基因的筛选、功能富集及聚类分析 使用DESeq[16](2012)R package对数据进行标准化,并使用nbinom Test函数计算差异比较的值和fold change值,挑选出值小于0.05,差异倍数大于2的差异基因,并进行差异基因的GO基因本体(gene ontology)和KEGG[17](kyoto encyclopedia of gene and genomes,京都基因与基因组百科全书)富集分析,以判定差异基因主要影响的生物学功能或者通路。同时对差异基因进行非监督层次聚类,利用热图的形式展示差异基因在不同样本间的表达模式。
1.3 数据统计分析
采用Excel 2019进行数据处理和绘图。应用SPSS20.0进行数据方差分析,采用Duncan’s多重比较法检验处理间的差异显著性(<0.05)。试验数据为3次重复的均值,用平均值±标准误表示。
2 结果
2.1 DPC对高温干旱胁迫下Bt棉杀虫蛋白含量影响
图1表明,与清水(对照)相比,DPC可显著提高高温干旱条件下棉铃壳中杀虫蛋白含量,两年提高幅度达4.7%和11.9%。

不同小写字母表示处理间差异显著(p<0.05)
2.2 DPC对高温干旱胁迫下Bt棉碳代谢、氨基酸代谢相关酶活性和物质含量影响
与清水(对照)相比,DPC可提高高温干旱胁迫下棉铃壳碳代谢、氨基酸代谢相关酶活性和物质含量(表1)。在碳代谢方面,DPC处理α-酮戊二酸含量、丙酮酸含量分别比清水处理高46%—57%和25%—29%;在氨基酸代谢方面,DPC处理谷氨酸合酶活性、谷氨酸草酰乙酸转氨酶活性、可溶性蛋白含量、游离氨基酸含量分别提高32%—44%、30%—40%、28%和22%—27%。
2.3 DPC处理前后的转录组质量评估
由表2可知,在RNA序列中,过滤后得到的有效数据为48 769 270—53 204 746,Q30比例(错误率<0.1%)在94.72%—95.25%,GC比例均大于43%,说明测序结果质量良好。测序结果与棉花基因组数据库进行比对结果说明,97%以上的测序序列可以映射到基因组,并且唯一映射率达89%以上。
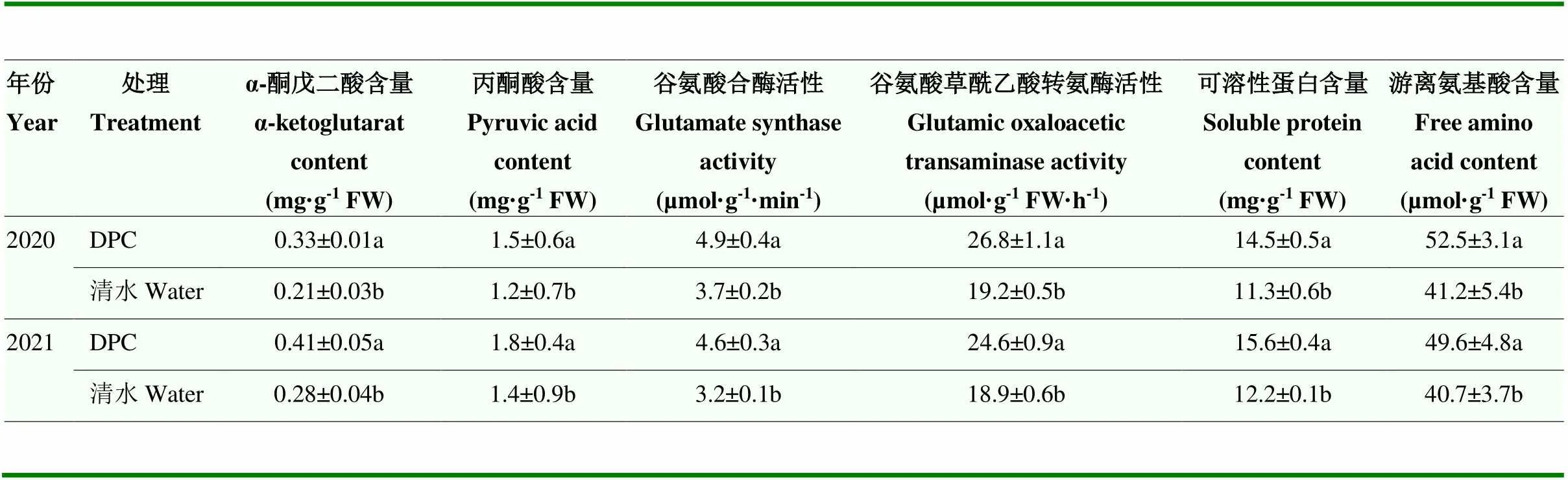
表1 DPC对高温干旱胁迫下Bt棉碳、氨基酸代谢和可溶性蛋白含量影响
每列数据后不同小写字母表示同一年份处理间差异显著(<0.05)
Different small letters after each column of data meant significant difference at<0.05 level among treatments
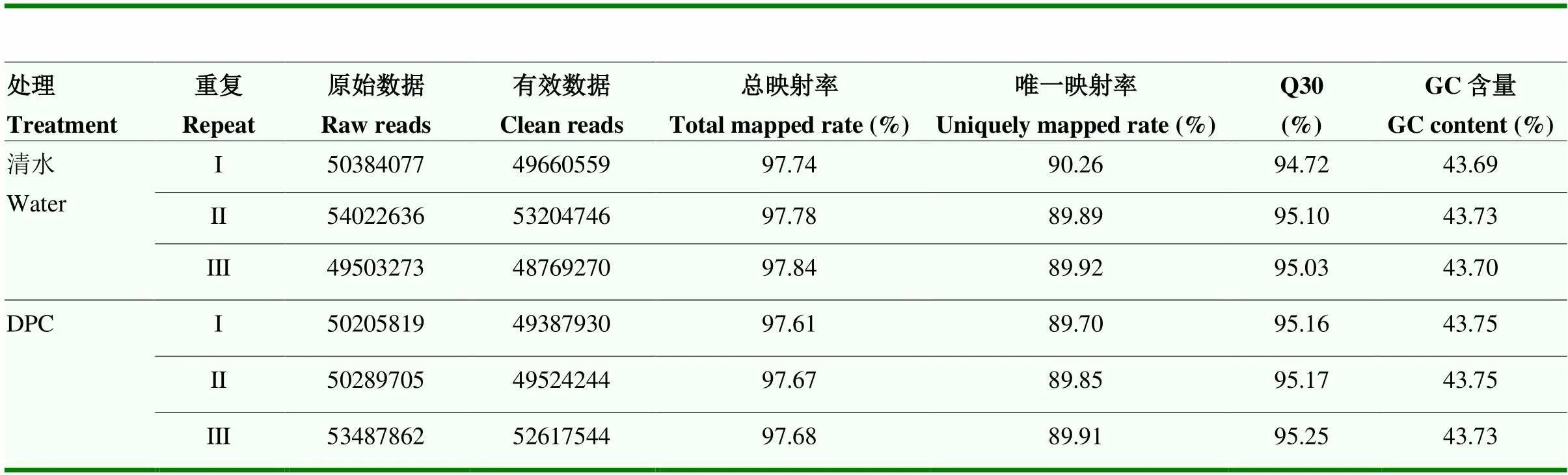
表2 Bt棉转录组数据质量统计
2.4 DPC处理前后的Bt棉差异表达基因分析
以fold change(FC)≥2且value<0.05为筛选标准,DPC和清水处理间差异表达基因数目达17 991条(图2),其中与清水处理(对照)相比,DPC处理后上调基因7 542条,下调基因10 449条。说明DPC处理后棉铃壳中多数基因表达受到了抑制。
2.5 DPC处理前后的Bt棉差异基因功能注释和富集分析
对高温干旱下DPC处理和清水处理差异基因进行基因本体GO功能分析,筛选GO分类的生物过程、细胞组分和分子功能中各值前10的主要基因功能信息见表3。
与清水对照相比,在生物过程分类下,DPC处理上调的差异表达基因主要富集到下列词条:ATP水解耦合质子传输、三羧酸循环、对乙烯的反应、乙烯激活信号通路的负调控、亮氨酸分解代谢过程、脂肪酸β氧化、线粒体中电子传递(细胞色素c到氧)、盐胁迫反应、糖酵解过程、氨基酸跨膜转运。上述大多词条与植株碳和氨基酸代谢密切相关。在细胞组成分类下,上调的差异表达基因主要富集在膜组成部分、高尔基体、内质网膜等。在分子功能分类下,质子转运ATP酶活性,旋转机制、细胞色素C氧化酶活性、氨基酸跨膜转运蛋白活性、脂肪酰辅酶A结合、钴离子结合、乙酰辅酶A C-酰基转移酶活性、泛醌细胞色素C还原酶活性、烯醇辅酶A水合酶活性、蔗糖跨膜转运蛋白活性等9个词条与碳氮代谢关系密切。上述结果表明,DPC主要通过影响棉植株体内生物学过程来调控杀虫蛋白表达,同时也涉及调控其分子功能来影响杀虫蛋白含量,因此通过GO功能分类的富集途径解释DPC提高棉杀虫蛋白含量的调控网络具有较为积极的意义。

图2 DPC和清水处理Bt棉铃壳差异表达基因的火山图

表3 高温干旱下DPC处理 vs 清水处理铃壳中差异表达基因的GO富集
2.6 DPC处理前后的Bt棉差异基因的KEGG生物通路分类及富集分析
在生物体内,不同的基因产物相互协调来行使生物学功能,对差异表达基因的通路(pathway)注释分析有助于了解棉杀虫蛋白合成所涉及的途径,从而进一步了解DPC提高高温干旱逆境下棉杀虫蛋白含量的分子机理。对差异表达基因(7 765个)进行 KEGG功能注释分析,发现它们被富集到194条代谢通路,本文挑选值较小的前20 条 pathway 条目在图中进行展示(图 3),分别属于代谢(19条)和环境信息处理(1条)KEGG分支。而在代谢分支中,5条属于氨基酸代谢,7条属于碳代谢,3条属于全局和总览图,2条脂质代谢,萜类和聚酮的代谢、能量代谢各1条。可见DPC主要是通过调节代谢分支,尤其是碳代谢和氨基酸代谢来调节杀虫蛋白表达。
糖酵解是将葡萄糖降解为丙酮酸并伴随着ATP生成的一系列反应,是生物体内普遍存在的葡萄糖降解的途径。糖酵解的关键酶有3个,即己糖激酶、6-磷酸果糖激酶和丙酮酸激酶,它们催化的反应基本上都是不可逆的[18]。图4表明,DPC处理对棉植株糖酵解途径有明显的影响。编码6-磷酸果糖激酶(6-phosphofructokinase)10个基因(GH_A02G1558、GH_A05G1771、GH_A06G0746、GH_A07G0253、GH_A07G0497、GH_D03G0426、GH_D05G0280、GH_D05G1804、GH_D06G0723、GH_D07G0262)表达全部显著上调,其中GH_A07G0253、GH_D07G0262 log2FC 分别达62.94和29.83;编码丙酮酸激酶(pyruvate kinase)10个基因,其中6个(GH_A03G0315、GH_A08G0889、GH_A09G1595、GH_A10G2189、GH_D03G1659、GH_D09G1539)基因表达显著上调,4个基因表达显著下调,但上调基因数量和上调幅度均大于下调基因。综上,DPC处理后可加速糖酵解途径的运行速率,利于丙酮酸的形成,并且主要是通过上调6-磷酸果糖激酶表达实现。

气泡颜色代表p值大小,p值越小代表富集结果越显著。气泡大小代表差异表达基因的数目,气泡大代表差异基因数目多,气泡小代表差异基因数目少
方格中红色代表差异上调基因;深绿色代表差异下调基因;黄色代表同一个基因不同转录本,既有差异上调也有差异下调基因;浅绿色或紫色代表物种特有基因,非差异显著性基因;白色代表该物种没有的基因或未检测到的基因。下同
柠檬酸循环(三羧酸循环)是需氧生物体内普遍存在的代谢途径,不但为机体提供能量,同时柠檬酸循环是三大营养物质的共同氧化途径:乙酰CoA,不但是糖氧化分解的产物,也是脂肪酸和氨基酸代谢的产物。此外,柠檬酸循环是三大物质代谢联系的枢纽:糖有氧氧化过程中产生的α-酮戊二酸、丙酮酸和草酰乙酸等与氨结合可转变成相应的氨基酸;这些氨基酸脱去氨基又可转变成相应的酮酸而进入糖的有氧氧化途径。同时脂类物质分解代谢产生的甘油、脂肪酸代谢产生的乙酰CoA也可进入糖的有氧氧化途径进行代谢。
糖酵解的产物丙酮酸,在有氧条件下进入线粒体,开始柠檬酸循环,形成水和二氧化碳并释放能量。柠檬酸共有10步反应,其中柠檬酸合酶(citrate synthase)是柠檬酸循环的限速酶。异柠檬酸脱氢酶、α-酮戊二酸脱氢酶系是另外两种关键酶。DPC总体上提高了所有步骤的反应速率(图5)。其中编码丙酮酸脱氢酶E1α亚单位(pyruvate dehydrogenase E1 component alpha subunit)共7个基因,其中6个表达上调,编码丙酮酸脱氢酶E1β亚单位(pyruvate dehydrogenase E1 component beta subunit)共5个基因,其中4个表达上调;编码丙酮酸脱氢酶E2共4个基因,表达全部上调。说明DPC可以加快催化丙酮酸向乙酰辅酶A转变,增加了进入柠檬酸循环的底物乙酰辅酶A的可获得性。
编码柠檬酸合酶(GH_A05G1609、GH_A11G0627、GH_D05G1636、GH_D11G0654)的基因显著上调表达;编码异柠檬酸脱氢酶(isocitrate dehydrogenase)4个基因中3个显著上调表达;编码α-酮戊二酸脱氢酶(2-oxoglutarate dehydrogenase)(GH_A05G3704)基因显著上调表达。综上,DPC提高了高温干旱逆境下棉柠檬酸循环反应速率。

图5 高温干旱下DPC处理对铃壳中柠檬酸(三羧酸)循环通路(ko00020)的影响
柠檬酸循环的中间产物丙酮酸、α-酮戊二酸、草酰乙酸也是合成氨基酸的原料。而谷氨酸在氮素合成代谢中起关键作用,同时也是其他氨基酸的氨基主要供体。谷氨酸合酶(glutamate synthase)可催化谷氨酰胺和α-酮戊二酸形成谷氨酸(Glu)。图6可见,编码谷氨酸合酶基因2个(GH_A12G2700、GH_D12G2727)显著上调,1个(GH_D08G1115)下调;编码谷氨酰胺合成酶(glutamine synthetase)基因6个显著下调2个显著上调,但下调幅度显著高于上调幅度。说明DPC处理促进谷氨酸的形成,并抑制其向谷氨酰胺的转化。此外,高等植物体内脯氨酸首先可以被脯氨酸脱氢酶(PDH)氧化成吡咯啉-5-羧酸(P5C),后者在吡咯啉-5-羧酸脱氢酶的作用下也可生成谷氨酸。而本文中编码吡咯啉-5-羧酸脱氢酶(GH_A03G0786、GH_D03G1048)基因显著上调。综上,DPC处理有利于谷氨酸的积累。

图6 高温干旱下DPC处理对铃壳中丙氨酸、天冬氨酸和谷氨酸代谢通路(ko00250)的影响
谷氨酸草酰乙酸转氨酶(GOT)在植物体内催化草酰乙酸合成天冬氨酸,且催化天冬氨酸与α-酮戊二酸间的氨基转换反应,与蛋白质及氨基酸含量有关[19-20]。编码谷氨酸草酰乙酸转氨酶11个基因,其中9个(GH_A09G0041、GH_A10G1610、GH_D09G0044、GH_D10G1279、GH_D05G0584、GH_A06G0690、GH_A07G0277、GH_D06G0665、GH_D07G0289)基因显著上调,如GH_A10G1610、GH_D10G1279 log2FC 分别达38.44和18.74,仅有2个(GH_A07G276、GH_D07G0288)基因显著下调,上调基因无论是数量还是上调幅度均明显高于下调基因。可见DPC处理可加快草酰乙酸向天冬氨酸的转变,促进天冬氨酸合成。
由图6可知,编码谷氨酸丙酮酸转氨酶(GPT)(GH_A10G0511、GH_D05G1444、GH_D10G0537)基因显著上调表达,加快了丙酮酸向丙氨酸的转化,这利于丙氨酸的积累。
在植物细胞核内,精氨酸的生物合成起始于L-谷氨酸乙酰化作用并涉及连续的8个酶的催化过程,其中第一个酶乙酰谷氨酸合成酶是精氨酸反馈抑制的靶点[21-22]。图7表明,DPC上调了编码N-乙酰谷氨酸合成酶(amino acid N-acetyltransferase,argAB)(GH_A10G0898、GH_D11G2171)基因表达量,其中GH_D11G2171 log2FC达21.30。此外,编码乙酰鸟氨酸脱乙酰酶(acetylornithine deacetylase,argE)(GH_A03G0060、GH_D03G1904)基因也显著上调,这利于鸟氨酸的形成从而也进一步为精氨酸合成提供前体。可见DPC可促进了精氨酸的合成。

图7 高温干旱下DPC处理对铃壳中精氨酶合成通路(ko00220)的影响
3 讨论
3.1 DPC可提高高温干旱胁迫下Bt棉杀虫蛋白含量
前人研究表明,高温干旱可显著抑制棉杀虫蛋白合成,导致其含量下降。而缩节胺可调节作物碳、氮代谢,且能提高细胞膜的稳定性,增加植株抗逆性[23]。本研究发现,在昼夜38℃/28℃、土壤含水量为最大持水量的50%胁迫条件下,喷施20 mg·L-1DPC可显著提高棉铃壳中杀虫蛋白含量,这与前人研究结果基本一致[8,10]。此外,结合笔者前期研究结果[24],本文发现高温干旱逆境下DPC处理后棉铃壳中杀虫蛋白含量绝对值与正常温度和土壤水分条件下的含量基本相似,说明施用外源调节剂可以降低甚至抵消本文逆境环境条件对棉抗虫性的负面影响。因此,在大田生产中,若遇高温干旱天气可采用喷施DPC的方法提高棉自身抗虫性,从而减少农药使用,提高棉花生产经济效益和生态效益。
3.2 高温干旱胁迫下Bt棉响应DPC处理的转录组水平差异
植物根系吸收硝态氮(NO3-),通过硝酸还原酶和亚硝酸还原酶将NO3-还原成NH3,再经谷氨酰胺合成酶和谷氨酸合酶同化为谷氨酸,后者与来自碳代谢中间物的各种碳骨架(α-酮戊二酸、草酰乙酸)之间转氨形成各种氨基酸。而蛋白质生物合成即把mRNA分子中碱基排列顺序转变为蛋白质或多肽链中的氨基酸排列顺序的过程[25-26]。本文认为基因作为一种外源基因,其翻译表达的杀虫蛋白合成也应依托棉花自身原有的代谢体系,可能并不涉及特有的酶。
本文采用RNA-seq技术对DPC处理后的棉基因表达谱进行比较发现,差异表达基因(7 765个)被富集到194 条代谢通路,其中值较小的前20 条 pathway 条目(图3)主要属于代谢KEGG分支(19条)。进一步分析发现在上述代谢分支中,5条属于氨基酸代谢,7条属于碳代谢,3条属于全局和总览图,2条脂质代谢,萜类和聚酮的代谢、能量代谢各1条。因此,本文认为DPC主要是通过调节代谢分支,尤其是碳代谢和氨基酸代谢来调节杀虫蛋白表达。这也验证了前人关于棉碳氮代谢能力与杀虫蛋白含量密切相关的结论[27]。这可能是由于外源基因的高效表达,必然影响宿主的生长和代谢,合理地调节宿主细胞的代谢负荷与外源基因高效表达的关系,是提高外源基因表达水平不可缺少的一个环节。本文中高温干旱胁迫使得棉细胞代谢损伤,必然影响外源基因的表达,而DPC可以合理地调节棉细胞的代谢负荷,从而利于外源基因高效表达。
进一步分析发现,在糖酵解途径中,DPC处理中6-磷酸果糖激酶和丙酮酸激酶基因表达显著上调,加速了糖酵解途径的运行速率,利于丙酮酸的形成(图4)。随后谷氨酸丙酮酸转氨酶基因也显著上调表达(图6),这加快了丙酮酸向丙氨酸的转化,最终利于丙氨酸的积累。
此外,糖酵解的产物丙酮酸,在有氧条件下可进入线粒体,经丙酮酸脱氢酶作用转变为乙酰辅酶A从而进入柠檬酸循环(图5)。其中柠檬酸合酶(citrate synthase)是柠檬酸循环的限速酶,异柠檬酸脱氢酶、α-酮戊二酸脱氢酶系是另外2种关键酶。DPC处理下编码丙酮酸脱氢酶基因总体表达上调,催化丙酮酸向乙酰辅酶A转变加快,进入柠檬酸循环的底物乙酰辅酶A的可获得性增加。同时编码柠檬酸合酶、异柠檬酸脱氢酶和α-酮戊二酸脱氢酶基因显著上调表达。综上,DPC提高了高温干旱逆境下棉柠檬酸循环反应速率,利于α-酮戊二酸(表1)、草酰乙酸的积累,这为后期天冬氨酸和谷氨酸合成提供了充足的底物。
谷氨酸合酶是催化谷氨酰胺的酰胺上氨基转移到α-酮戊二酸的酮基上,生成谷氨酸的氧化还原酶,而谷氨酰胺合成酶主要是催化依赖ATP的谷氨酰胺合成。本研究发现编码谷氨酸合酶基因(GH_A12G2700、GH_D12G2727)和吡咯啉-5-羧酸脱氢酶(GH_A03G0786、GH_D03G1048)基因总体上也显著上调,这与谷氨酸合酶活性显著提高(表1)结果相一致,但编码谷氨酰胺合成酶基因显著下调。说明DPC处理不仅促进高温干旱逆境下棉铃壳中谷氨酸的形成,同时抑制其向谷氨酰胺的转化,两者共同促进了谷氨酸的积累。这与前人发现的谷氨酸合酶对杀虫蛋白合成起主要作用结果相一致[28]。
谷氨酸草酰乙酸转氨酶(GOT)是植物体内氮代谢合成的关键酶。DPC总体上促进GOT基因显著上调(图6),其中GH_A10G1610、GH_D10G1279 log2FC 分别达38.44和18.74,其活性显著提高(表1)。说明DPC处理加快草酰乙酸向天冬氨酸的转变,促进天冬氨酸合成。在精氨酸合成过程中,本研究发现DPC上调了精氨酸的生物合成过程中关键酶N-乙酰谷氨酸合成酶(argAB)基因表达量,其中GH_D11G2171 log2FC达21.30,以及编码乙酰鸟氨酸脱乙酰酶(argE)基因表达量,这均利于精氨酸的合成。
综上所述,DPC处理后显著提高高温干旱胁迫下棉碳代谢能力,这不仅为后期氨基酸的生物合成提供碳骨架,也为杀虫蛋白的合成提供了充足的能量。同时,DPC利于丙氨酸、天冬氨酸、谷氨酸、精氨酸等的合成,这为后期杀虫蛋白的合成提供充足底物。结合游离氨基酸含量增加结果(表1),本研究认为DPC处理总体而言提升了棉植物氨基酸合成能力。氨基酸是合成蛋白质的基本单位,其含量的多少和组分的比例影响蛋白质的合成效率[29]。杀虫蛋白由18种共1 176个氨基酸及酰胺组成,其中天冬氨酸占氨基酸总数量比例最大(13.31%),其次为谷氨酸(11.98%)、精氨酸(7.65%)、亮氨酸(7.49%)、缬氨酸(7.32%)[30]。在本文DPC处理下,天冬氨酸、谷氨酸、精氨酸表达量与杀虫蛋白含量变化一致,但亮氨酸、缬氨酸却与之相反。这可能是由于杀虫蛋白合成需要较多的天冬氨酸、谷氨酸、精氨酸,它们比例的增加更加促进杀虫蛋白的合成,这也解释了前人喷施外源氨基酸可提高棉杀虫蛋白含量的结果[31-32]。因此,生产中棉农可以通过采取喷施外源生长调节剂等措施,调节棉碳氮代谢强度来提高杀虫蛋白含量。
4 结论
喷施20 mg·L-1DPC可显著提高高温干旱胁迫下棉铃壳中杀虫蛋白含量。其主要是通过提高糖酵解途径、柠檬酸循环的运行速率,增加丙酮酸、α-酮戊二酸、草酰乙酸等含量。同时显著上调编码谷氨酸合酶基因、吡咯啉-5-羧酸脱氢酶基因,下调编码谷氨酰胺合成酶基因以及上调谷氨酸草酰乙酸转氨酶、N-乙酰谷氨酸合成酶等编码基因,从而利于谷氨酸、天冬氨酸、精氨酸积累。最终通过改变棉铃壳中的氨基酸含量和比例来调节杀虫蛋白合成。
[1] 夏兰芹, 郭三堆. 高温对转基因抗虫棉中Bt杀虫基因表达的影响. 中国农业科学, 2004, 37(11): 1733-1737.
XIA L Q, GUO S D. The expression of Bt toxin gene under different thermal treatments. Scientia Agricultura Sinica, 2004, 37(11): 1733-1737. (in Chinese)
[2] CHEN D H, YE G Y, YANG C Q, CHEN Y, WU Y K. The Effect of high temperature on the insecticidal properties of Bt cotton. Environmental and Experimental Botany, 2005, 53(3): 333-342.
[3] WANG J, CHEN Y, YAO M H, LI Y, WEN Y J, CHEN Y, XIANG Z, CHEN D H. The effects of high temperature level on square Bt protein concentration of Bt cotton. Journal of Integrative Agriculture, 2015, 14(10): 1971-1979.
[4] BENEDICT J H, SACHS E S, ALTMAN D W, DEATON W R, KOHEL R J, RING D R, BERBERICH S A. Field performance of cottons expressing transgenic Cry1A insecticidal proteins for resistance toand(Lepidoptera: Noctuidae). Journal of Economic Entomology, 1996, 89(1): 230-238.
[5] SACHS O, KATZMAN A, ABU-JOHAR E, EIDELMAN M. Treatment of adolescent blount disease using taylor spatial frame with and without fibular osteotomy: Is there any difference? Journal of pediatric orthopedics, 2015, 35(5): 501.
[6] MARTINS C M, BEYENE G, HOFS J L, KRÜGER K, VYVER C VD, SCHLÜTER U, KUNERT K. Effect of water-deficit stress on cotton plants expressing thetoxin. Annals of Applied Biology, 2008, 152(2): 255-262.
[7] ROCHESTER I J. Effect of genotype edaphic, environmental conditions, and agronomic practices on Cry1Ac protein expression in transgenic cotton. Journal of Cotton Science, 2006, 10(4): 252-262.
[8] 董志强, 舒文华, 翟学军, 张保明, 刘芳, 宋国琦. 棉株不同器官中几种内源激素的变化及相关关系. 核农学报, 2005, 19(1): 62-67.
DONG Z Q, SHU W H, ZHAI X J, ZHANG B M, LIU F, SONG G Q. The change and relationship of several endogenesis hormone in different organs of cotton plant. Acta Agriculturae Nucleatae Sinica, 2005, 19(1): 62-67. (in Chinese)
[9] 郑青松, 刘友良. DPC浸种提高棉苗耐盐性的作用和机理. 棉花学报, 2001, 13(5): 278-282.
ZHENG Q S, LIU Y L. Effects of soaking seeds in dpc increasing the salinity tolerance in cotton (L.) seedlings and its mechanism. Cotton Science, 2001, 13(5): 278-282. (in Chinese)
[10] ZHANG X, TIAN Q F, ZHAO C, MA Y X, WEI C H, CHEN Y, CHEN D H. Exogenous hormones affect Bt protein content of two Bt cotton cultivars. Agronomy Journal, 2019, 3(6): 3076-3083.
[11] 陈松, 吴敬音, 何小兰, 黄骏麒, 周宝良, 张荣铣. 转基因抗虫棉组织中Bt毒蛋白表达量的ELISA测定. 江苏农业学报, 1997, 13(3): 154-156.
CHEN S, WU J Y, HE X L, HUANG J Q, ZHOU B L, ZHANG R X. Quantification using ELISA ofinsecticidal protein expressed in the tissue of transgenic insect-resistant cotton. Jiangsu Journal of Agricultural Sciences, 1997, 13(3): 154-156. (in Chinese)
[12] 张宪政. 作物生理研究法. 北京: 中国农业出版社, 1992: 139-142.
ZHANG X Z. The Research Method of Crop Physiology. Beijing: China Agriculture Press, 1992: 139-142. (in Chinese)
[13] HODGES M. Enzyme redundancy and the importance of 2-oxoglutarate in plant ammonium assimilation. Journal of Experimental Botany, 2002, 53(370): 905-916.
[14] MUSTROPH A, ALBRECHT G. Tolerance of crop plants to oxygen deficiency stress: fermentative activity and photosynthetic capacity of entire seedlings under hypoxia and anoxia. Physiologiaplantarum, 2003, 117(4): 508-520.
[15] KIM D, LANGMEAD B, SALZBERG S L. HISAT: a fast spliced aligner with low memory requirements. Nature methods, 2015, 12(4): 357-360.
[16] ANDERS S, HUBER W. Differential expression of RNA-Seq data at the gene level-the DESeq package. EMBL, 2013.
[17] KANEHISA M, ARAKI M, GOTO S. KEGG for linking genomes to life and the environment. Nucleic Acids Research, 2008, 36: 480-484.
[18] WATANABE C K, Hachiya T, Takahara K, Kawai-Yamada M, UCHIMIYA H, UESONO Y, TERASHIMA I, NOGUCHI K. Effects of AOX 1a deficiency on plant growth, gene expression of respiratory components and metabolic profile under low-nitrogen stress in. Plant and Cell Physiology, 2010, 51(5): 810-822.
[19] SENER G, SEHIRLI O, TOZAN A, VELIOĞLUO A, GEDIK N, OMURTAG G Z. Ginkgo biloba extract protects against mercury (II)-induced oxidative tissue damage in rats. Food and chemical toxicology, 2007, 45(4): 543-550.
[20] CELIK I, TULUCE Y, ISIK I. Influence of subacute treatment of some plant growth regulators on serum marker enzymes and erythrocyte and tissue antioxidant defense and lipid peroxidation in rats. Journal of biochemical and molecular toxicology, 2006, 20(4): 174-182.
[21] XU Y, GLANSDORFF N, LABEDAN B. Bioinformatic analysis of an unusual gene-enzyme relationship in the arginine biosynthetic pathway among marine gamma proteobacteria: implications concerning the formation of N-acetylated intermediates in prokaryotes. BMC Genomics, 2006, 7: 4.
[22] 程功, 徐建中, 张伟国. L-精氨酸生物合成机制及其代谢工程育种研究进展. 微生物学通报, 2016, 43(6): 1379-1387.
CHENG G, XU J Z, ZHANG W G. Progress in biosynthesis and metabolic engineering of L-arginine producer. Microbiology China, 2016, 43(6): 1379-1387. (in Chinese)
[23] 张特, 赵强, 李广维. 缩节胺对棉花生长发育影响研究进展. 江苏农业科学, 2021, 49(18): 14-18.
ZHANG T, ZHAO Q, LI G W. Progress in the effects of DPC on the development of cotton. Jiangsu Agricultural Sciences, 2021, 49(18): 14-18. (in Chinese)
[24] ZHANG X, TIAN Q, ZHAO Z, DONG Z, CHEN Y, CHEN D. Analysis of differentially expressed proteins affecting insecticidal protein content in Bt cotton under high-temperature and water deficit stress using label-free quantitation. Journal of Agronomy and Crop Science, 2021, 207: 1-11.
[25] HERNÁNDEZC A E, SÁNCHEZ E, PRECIADOR P, GARCÍAB M L, PALOMOG A, ESPINOZAB A. Nitrate reductase activity, biomass, yield, and quality in cotton in response to nitrogen fertilization. Phyton-International Journal of Experimental Botany, 2015, 84(2): 454-460.
[26] ZHOU H, ZHOU Y, ZHANG F, GUAN W X, SU Y, YUAN X X, XIE Y J. Persulfidationof nitrate reductase 2 is involved in l-cysteine desulfhydrase-regulated rice drought tolerance. International Journal of Molecular Sciences, 2021, 22(22): 12119.
[27] LIU Z Y, WANG G X, ZHANG Z N, ZHANG C X, LI H J, WU T F, ZHANG X, CHEN D H. Recovery characteristics of Cry1Ac endotoxin expression and related physiological mechanisms in Bt transgenic cotton squares after high-temperature stress termination. Agronomy, 2022, 12(3): 668.
[28] 刘媛媛. 高温与干旱对Bt棉杀虫蛋白含量的影响及其生理机制[D]. 扬州: 扬州大学, 2021.
LIU Y Y. The effect of high temperature and drought on the content of insecticidal protein in Bt cotton and its physiology mechanism[D]. Yangzhou: Yangzhou University, 2021. (in Chinese)
[29] 张祥, 王剑, 彭盛, 芮秋治, 李丽楠, 陈媛, 陈源, 陈德华. 温度和土壤水分与Bt棉铃壳中杀虫蛋白表达关系及其氮代谢生理机制. 中国农业科学, 2018, 51(7): 1261-1271.
ZHANG X, WANG J, PENG S, RUI Q Z, LI L N, CHEN Y, CHEN Y, CHEN D H. Relationship between temperature, soil moisture, and insecticidal protein content in Bt cotton boll shell and the mechanism of nitrogen metabolism. Scientia Agricultura Sinica, 2018, 51(7): 1261-1271. (in Chinese)
[30] SCHNEPF H E, WONG H C, WHITELEY H R. The amino acid sequence of a crystal protein fromdeduced from the DNA base sequence. The Journal of Biological Chemistry, 1985, 260(10): 6264-6272.
[31] TAMBEL L I M. ZHOU M Y, CHEN Y, ZHANG X, CHEN D H. Amino acids application enhances flowers insecticidal protein content in Bt cotton. Journal of Cotton Research, 2019, 2: 7.
[32] YANG Z, YOU S S, JI C D, YIN M Z, YANG W T, SHEN J. Development of an amino acid-functionalized fluorescent nanocarrier to deliver a toxin to kill insect pests. Advanced Materials, 2016, 28(7): 1375-1380.
Mepiquat Chloride Increases theProtein Content Through Regulating Carbon and Amino Acid Metabolism ofCotton Under High Temperature and Drought Stress
Agricultural college, Yangzhou University/Key Laboratory of Crop Genetics and Physiology of Jiangsu Province, Yangzhou 225009, Jiangsu
【Objective】The effects of mepiquat chloride (DPC) on the insecticidal protein contents in() cotton shell under high temperature and drought stress were investigated to provide a theoretical reference for thecotton breeding as well as high-yield and high-efficiency cotton cultivation.【Method】The study was undertaken on thecotton cultivar Sikang 3 during the 2020 and 2021 growing seasons at the Yangzhou University Farm, Yangzhou, China. The potted cotton plants were exposed to high temperature and drought stress, and 20 mg·L-1DPC and water (CK) were sprayed to cotton plants. Seven days after treatment, the insecticidal protein content, α-ketoglutarate content, pyruvic acid content, glutamate synthase activity, glutamic oxaloacetic transaminase activity, soluble protein content, free amino acid content in boll shell were analyzed, and the transcriptome sequencing was performed. DESeq was used for differential gene analysis. The GO and KEGG pathway databases were used to analyze the differentially expressed genes involved in regulating the insecticidal protein content through DPC.【Result】Compared with the water treatment (CK), the insecticidal protein contents under DPC treatment increased by 4.7%-11.9%. In terms of carbon metabolism, the contents of α-ketoglutarate and pyruvic acid were increased by 46%-57% and 25%-29%, respectively. In terms of amino acid metabolism, the activities of glutamate synthase and glutamic oxaloacetic transaminase, and the contents of soluble protein and free amino acid were increased by 32%-44%, 30%-40%, and 28%, 22%-27%, respectively. The transcriptome analysis revealed that there were 7 542 upregulation genes and 10 449 downregulation genes for DPC vs water. The GO and KEGG analysis showed that the differentially expressed genes were mainly involved in biological process such as amino acid metabolism and carbon metabolism. The genes coding 6-phosphofructokinase, pyruvate kinase, glutamic pyruvate transaminase, pyruvate dehydrogenase,citrate synthase, isocitrate dehydrogenase, 2-oxoglutarate dehydrogenase,glutamate synthase, 1-pyrroline-5-carboxylate dehydrogenase, glutamic oxaloacetic transaminase, amino-acid N-acetyltransferase, and acetylornithine deacetylase were all significantly up-regulated.【Conclusion】 Under the stress of high temperature and drought, the DPC treatment increased the contents of α-ketoglutarate and pyruvic acid, and improved the synthesis ability of aspartic acid, glutamic acid, pyruvate and arginine, then enhanced the insecticidal protein contents in boll shell by regulating the carbon and amino acid metabolism.
cotton; mepiquat chloride; insecticidal protein; amino acid metabolism; carbon metabolism
10.3864/j.issn.0578-1752.2023.08.004
2022-08-08;
2022-12-05
国家自然科学基金(31901462)、江苏省高等学校自然科学基金(22KJA210005)、江苏省自然科学基金(BK20191439)、江苏省高校优势学科建设工程、江苏高校品牌专业建设工程(PPZY2015A060)
邢羽桐,E-mail:xingyt0601_yzu@163.com。通信作者张祥,E-mail:zhangxiang@yzu.edu.cn。通信作者陈德华,E-mail:cdh@yzu.edu.cn
(责任编辑 杨鑫浩,岳梅)
猜你喜欢
牡丹江医学院学报(2021年5期)2021-12-05
今日农业(2019年12期)2019-08-15
食品安全导刊(2018年30期)2019-01-28
心肺血管病杂志(2018年11期)2018-12-18
中国中医药信息杂志(2016年7期)2016-12-01
中国医药生物技术(2015年4期)2015-12-26
川北医学院学报(2015年5期)2015-12-05
现代检验医学杂志(2015年5期)2015-02-06
中国质量与标准导报(2014年4期)2014-03-11
河南医学研究(2014年3期)2014-02-27
