
BMP2重组慢病毒转染兔BMSCs后细胞矿化的实验研究
2023-05-08 06:50宁寅宽刘林志陈贤平
天津医药 2023年5期
宁寅宽,刘林志,陈贤平
目前在组织工程研究领域,骨髓间充质干细胞(bone mesenchymal stem cells,BMSCs)是比较理想的种子细胞之一,骨形态发生蛋白(bone morphogenic proteins,BMP)已被证实能调控BMSCs 向成骨方向分化,BMP2 是目前研究最为广泛、诱导成骨活性最强的BMP[1-2]。天然BMP2 在体内无法实现缓释,不能持续诱导BMSCs 的成骨分化,如何获得持续、高效及稳定表达的内源性BMP2是目前研究的热点[3]。细胞矿化与普通的地质矿化的最大区别是无机相结晶严格受生物体分泌的有机质控制,骨骼中的羟基磷灰石形成作为细胞矿化最普遍的表现形式,亦受到广泛关注[4-5]。既往学者们缺乏对BMSCs 生物矿化现象的深入研究。本研究通过将BMP2重组慢病毒载体导入兔BMSCs,观察BMP2 重组慢病毒转染细胞向成骨方向分化后细胞矿化的能力,初步尝试使用扫描电镜能谱分析技术研究BMSCs 生物矿化现象,为干细胞基因治疗和临床转化提供理论依据。
1 材料与方法
1.1 试剂与仪器 低糖DMEM、胰蛋白酶、胎牛血清(美国HyClone 公司);流式一抗小鼠抗兔CD44、CD45、CD29(美国Antigenix公司),同型对照小鼠抗兔IgG1(eBioscien公司),流式二抗PerCP 标记(Jackson 公司);BMP2 单克隆抗体(美国Bioworld 公司),山羊抗小鼠IgG/辣根过氧化物酶标记、DAB显色试剂盒、二抗FITC 标记IgG(北京中杉金桥公司),Ⅰ型胶原单克隆抗体(武汉博士德),茜素红(美国Sigma);碱性磷酸酶(ALP)测试盒(南京建成生物工程研究所),总RNA提取试剂盒(北京天根生化科技有限公司),HiFi-MMLV 逆转录cDNA 合成试剂盒(上海英俊生物技术有限公司),2×Es Taq MasterMix PCR 试剂盒(北京康为世纪生物技术公司),DNA Marker、2×Tap PCR Mix(北京艾德莱生物科技有限公司),倒置荧光显微镜(日本Olympus公司),场发射扫描电镜(荷兰飞利浦公司),X 射线能谱分析仪(英国牛津公司),BMP2 重组慢病毒构建和鉴定(上海吉凯基因化学技术有限公司)。
1.2 方法
1.2.1 兔BMSCs 的获取及鉴定 清洁级雄性6 月龄新西兰大白兔1 只,体质量1.2 kg,购自桂林医学院动物实验中心,实验动物生产许可证号:SCXK(桂)2013-0001,按照实验动物伦理学标准执行。采用密度梯度离心及贴壁培养法获取第5代兔BMSCs,无菌条件下穿刺兔双侧股骨干骺端,获取骨髓约5 mL,离心后弃上清液(1 500 r/min,5 min),用比重为1.073 g/mL 的percoll 分离(2 500 r/min,20 min)兔BMSCs,取界面处细胞层,离心后洗涤,以2×105个/cm2的密度接种于25 cm2塑料培养瓶,72 h后更换培养液,弃掉未贴壁细胞,以后每1 d 更换细胞培养液1 次。细胞生长到80%汇合时1∶2传代培养,传至第5 代进行后续实验研究。取第5 代细胞制成细胞悬液,按操作说明分别标记CD44/CD45、CD29-PerCP,流式细胞仪上机检测,实验重复3次。
1.2.2 兔BMSCs的分组及转染 实验分为3组:空白对照组(未转染);Lv-EGFP组[阴性对照组,转染仅携带增强绿色荧光蛋白(enhanced green fluorescent protein,EGFP)基因的慢病毒);Lv-BMP2/EGFP 组(实验组,转染携带BMP2 和EGFP 基因的慢病毒)。第5代兔BMSCs以5×104个/孔接种于6孔板,细胞培养汇合至80%左右,根据前期实验数据以感染复数(MOI=100)转染细胞,转染24 h后换液,48 h后在倒置荧光显微镜下观察细胞形态并计算细胞转染效率(绿色荧光蛋白细胞数/视野内细胞总数×100%),实验重复3次。
1.2.3 免疫组织化学染色检测BMP2表达 分别随机取转染72 h后细胞,胰酶消化细胞爬片,浸入40 g/L多聚甲醛,按SP免疫组织化学染色法操作,DAB 显色,封固,在倒置显微镜观察。
1.2.4 RT-PCR 检测BMP2 基因的表达 转染后72 h 提取3×106个细胞总RNA,根据RNA 提取试剂盒操作步骤,通过凝胶电泳进行质量检测,使用分光光度计进行纯度浓度测定。按照MMLV逆转录cDNA合成试剂盒说明书合成第1条链cDNA。设计引物:GAPDH 引物(正义链5'-CCAGAA⁃CATCCCTGCCTC-3' ,反 义 链5'-TAGCCAAATTCGTTGT⁃CATACCA-3'),BMP2 引物(正义链5'-ACTACCAGAAAC⁃GAGTGGGAA-3',反义链5'-GCATCTGTTCGGAAAACCT-3')。按照2×Taq PCR MasterMix 试剂盒说明书进行PCR 扩增。PCR产物通过琼脂糖凝胶电泳鉴定,调电泳仪各参数进行电泳(U:220 V;I:140 mA;T:30 min),电泳结束,凝胶成像系统分析目的基因与对应内参光密度比值确定基因相对表达量,相对表达量=目的基因条带OD 值/对应内参照基因条带OD 值。反应条件:95 ℃预变性10 min,94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸30 s,进行35 个循环,72 ℃终延伸5 min。测量结果重复3次。
1.2.5 Western blot 检测BMP2 蛋白表达 转染后48 h,根据实验分组分别取2×106个细胞,裂解液提取细胞总蛋白,按BCA 蛋白定量试剂盒检测样品蛋白含量。取30 µg 上样,10%SDS-PAGE 电泳后转模,5%脱脂奶粉4 ℃封闭过夜,加一抗BMP2 单克隆抗体室温下摇床震荡孵育1 h,与辣根过氧化物酶标记二抗孵育1 h 进行抗体杂交,ECL 发光孵育1 min,曝光5 min,暗室内压片,显影定影,凝胶成像仪对蛋白印迹进行成像,以内参光密度值为标准,计算每组蛋白相对表达量,测量结果重复3次。
1.2.6 兔BMSCs的成骨分化检测 6孔板内预先放置消毒好的塑料盖玻片,按实验分组,以1×105个/孔接种6孔板进行塑料细胞爬片培养。分别于第14、21天,随机取爬片,按试剂盒说明书,行SP法检测Ⅰ型胶原蛋白表达、茜素红染色检测矿化结节形成。分别于第7、14、21天,各组随机取3孔,按ALP试剂盒说明操作,实验重复3次,计算ALP含量。
1.2.7 扫描电镜和能谱分析观测矿化结节表面微观形貌及其元素构成 第21天随机取实验组塑料细胞爬片,固定后梯度丙酮逐级脱水,临界点干燥,表面进行喷镀,在Quanta 200 FEG场发射环境行扫描电镜观察和主要元素能谱分析。X射线能谱分析仪测定微区主要元素(碳、氮、氧、钙、硫、磷)的质量百分比和原子个数百分比,测定结果重复3次。计算钙磷比(Ca/P)=钙元素质量百分比∶磷元素质量百分比,并打印出能谱分析图。
1.3 统计学方法 采用SPSS 25.0软件进行数据分析。计量资料以±s表示,多组之间比较采用单因素方差分析,组间多重比较采用LSD-t检验,P<0.05为差异有统计学意义。
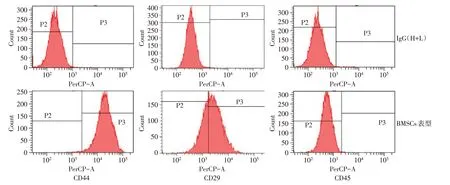
Fig.1 The characteristics of cell surface markers in BMSCs图1 BMSCs细胞表面标志物表达特征
2 结果
2.1 BMSCs 细胞表面标志物表达特征 流式细胞仪分析显示第5 代兔BMSCs 的CD44、CD29 表达呈阳性,CD45 表达呈阴性,CD44、CD29、CD45 阳性细胞分别占92.20%±1.12%,76.30%±0.78%,0.26%±0.11%,BMSCs 的细胞表面标志物符合BMSCs 的特征,见图1。
2.2 BMSCs 转染后细胞形态 转染48 h 后,Lv-BMP2/EGFP组、Lv-EGFP 组细胞在倒置荧光显微镜下观察到荧光,目的基因BMP2和EGFP报告基因成功修饰入细胞基因组,转染效率分别为95.32%±1.05%和95.18%±0.82%。空白对照组未观察到荧光。各组BMSCs 细胞生长呈长梭形,形态均一,单个细胞轮廓较清楚,胞浆丰富,核仁明显,细胞呈涡旋状、放射状密集排列,见图2。
2.3 BMP2 免疫组织化学染色 Lv-BMP2/EGFP 组BMSCs 细胞胞浆呈阳性表达,可见棕黄色染色,而Lv-EGFP组及空白对照组细胞呈阴性,见图3。
2.4 BMP2 mRNA 表达情况 Lv-BMP2/EGFP 组、Lv-EGFP 组及空白对照组BMP2 mRNA 相对表达量分别为0.474±0.008,0.126±0.027 和0.095±0.007(n=3,F=458.641,P<0.01),Lv-BMP2/EGFP 组BMP2 mRNA 水平高于Lv-EGFP 组及空白对照组(P<0.05),且基因条带结果符合扩增片段长度,见图4。
2.5 BMP2 蛋白表达情况 Lv-BMP2/EGFP 组、Lv-EGFP 组及空白对照组BMP2 蛋白相对表达量分别为0.751±0.057,0.272±0.013,0.305±0.007(n=3,F=186.426,P<0.01),Lv-BMP2/EGFP 组BMP2 蛋白表达量高于Lv-EGFP 组及空白对照组(P<0.05),见图5。
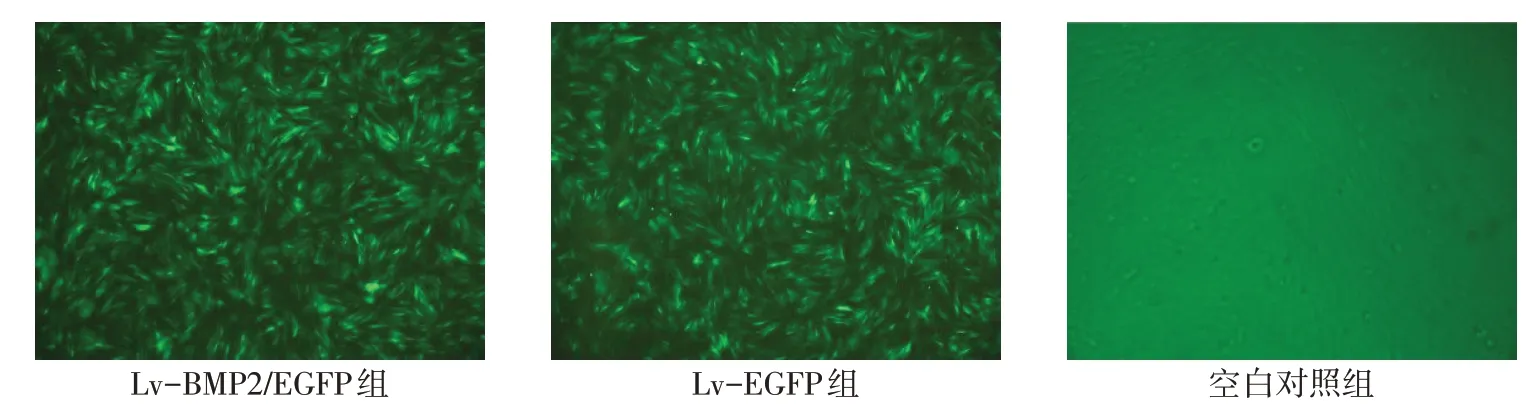
Fig.2 The cell morphology of BMSCs under fluorescence microscope(×100)图2 BMSCs在倒置荧光显微镜下细胞形态(×100)
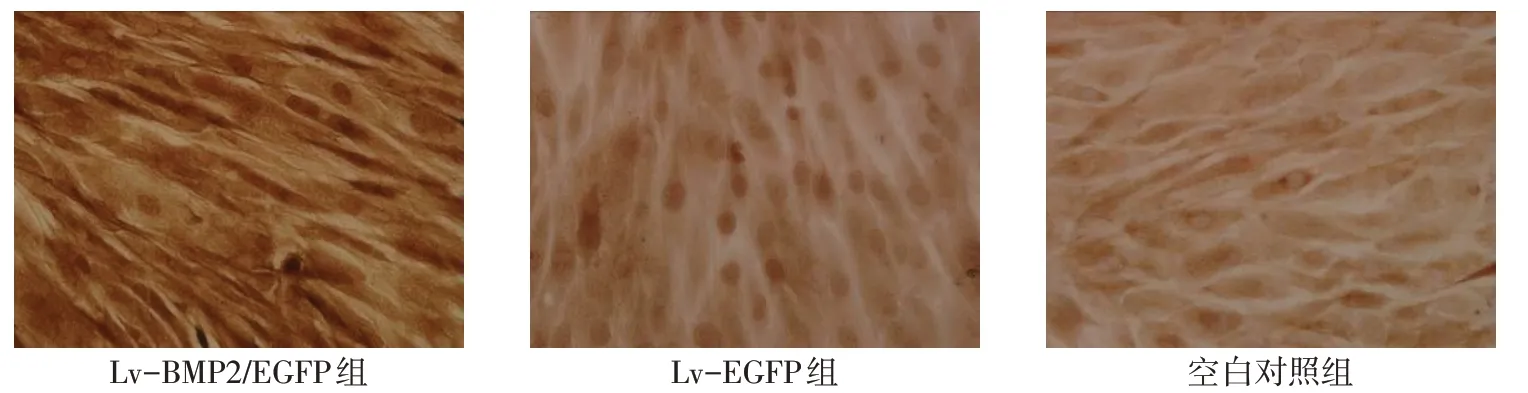
Fig.3 The SP immunochemical staining of BMP2 expression(DAB stanining,×200)图3 免疫组织化学染色SP法检测BMP2表达(DAB染色,×200)
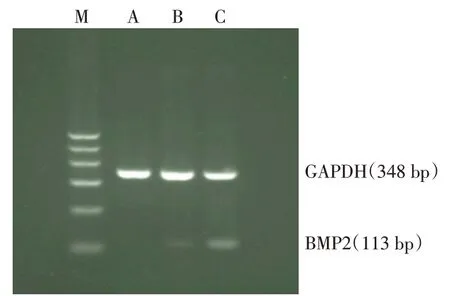
Fig.4 The BMP2 gene expression in BMSCs measured by RT-PCR图4 RT-PCR检测BMSCs中BMP2基因表达水平

Fig.5 The protein expression of BMP2 measured by Western blot assay图5 Western blot检测BMP2蛋白表达
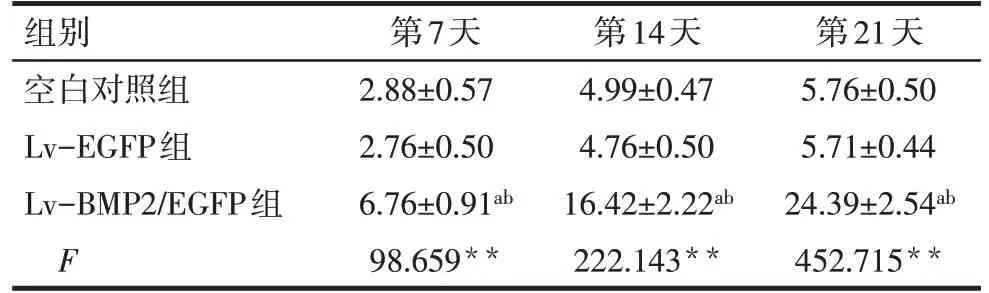
2.6 BMSCs的成骨方向分化 Lv-BMP2/EGFP组第14 天塑料细胞爬片在显微镜下观察显示细胞胞浆出现棕黄色染色颗粒,Ⅰ型胶原SP法免疫组织化学染色呈阳性表达。第21天茜素红染色呈现阳性,见形貌各不相同的矿化结节,矿化结节染色后呈现红色。而Lv-EGFP 组及空白对照组Ⅰ型胶原染色和茜素红矿化结节染色均呈现阴性。见图6、7。细胞经转染第7、14、21天,Lv-BMP2/EGFP组ALP水平较Lv-EGFP组及空白对照组升高,见表1。

Fig.6 The I collagen immunohistochemical staining of BMSCs(×100)图6 BMSCsⅠ型胶原免疫组织化学染色(×100)
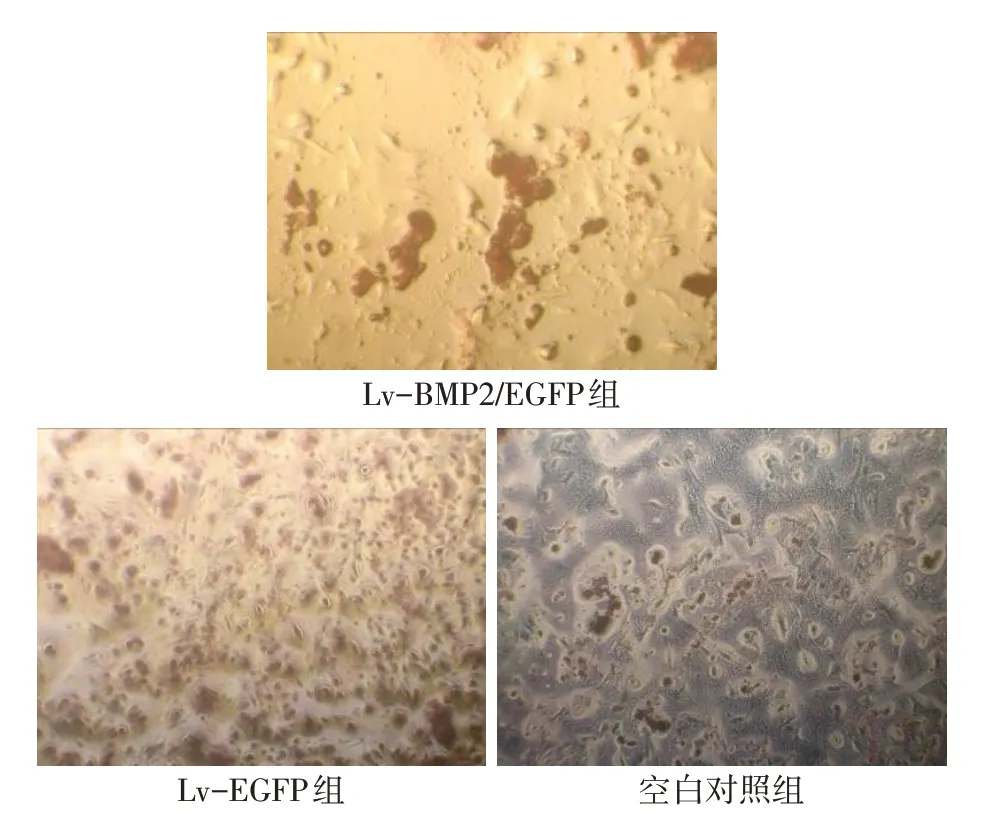
Fig.7 The alizarin red S staining of BMSCs(×50)图7 BMSCs茜素红染色(×50)
Tab.1 Comparison of ALP levels at different time points between the three groups表1 各组不同时间点ALP水平比较(n=9,U/gprot,±s)
Tab.1 Comparison of ALP levels at different time points between the three groups表1 各组不同时间点ALP水平比较(n=9,U/gprot,±s)
**P<0.01;a与空白对照组比较,b与Lv-EGFP组比较,P<0.05。
组别空白对照组Lv-EGFP组Lv-BMP2/EGFP组F第7天2.88±0.57 2.76±0.50 6.76±0.91ab 98.659**第14天4.99±0.47 4.76±0.50 16.42±2.22ab 222.143**第21天5.76±0.50 5.71±0.44 24.39±2.54ab 452.715**
2.7 矿化结节观察和元素能谱分析 在扫描电镜下可见矿化结节相互连接成片状,细胞重叠生长,基质分泌旺盛,矿化结节散布在重叠细胞中并稍凸出于细胞层之上,形貌似含沙浆较多的水泥,结构呈现疏松、粗糙特点,见图8。电镜扫描微区得出能谱分析图谱,见图9,对应主要元素见表2。矿化结节微区Ca/P为1.52±0.13。
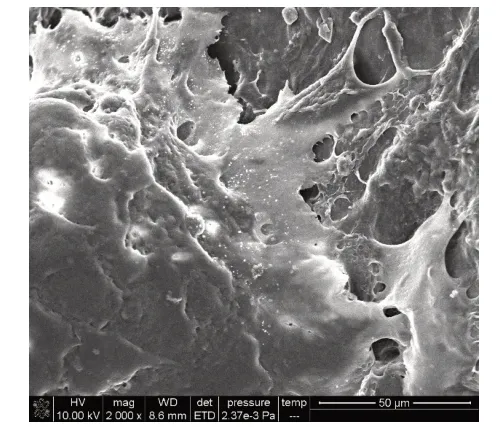
Fig.8 The microscopic morphology of the mineralized nodule surface observed by SEM图8 扫描电镜观察矿化结节表面微观形貌
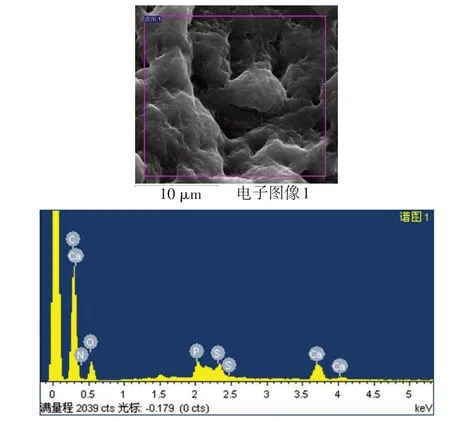
Fig.9 The energy spectrum analysis of the scanned area on the surface of the mineralized nodule图9 矿化结节表面扫描区域能谱分析图
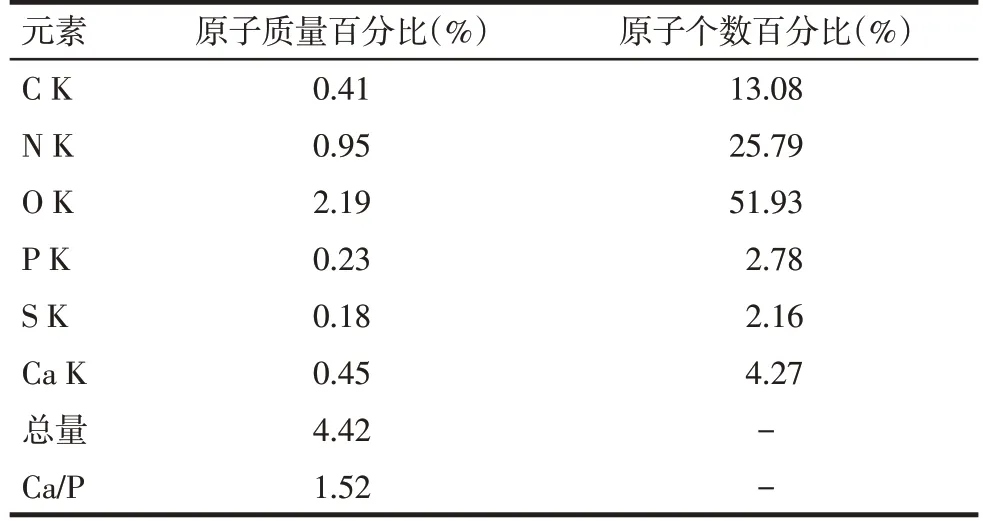
Tab.2 The elemental analysis of the scanned area on the surface of the mineralized nodule表2 矿化结节表面扫描区域元素分析
3 讨论
BMSC 由于其能较稳定地保持干细胞生物学特征,为多种病毒载体导入目的基因进行基因治疗奠定了基础[6]。BMP2 已被证实能单独诱导BMSCs 向成骨方向分化,但天然BMP2 获取困难,价格昂贵,且局部发挥生物作用有限,因此利用病毒载体导入目的基因,持续稳定的表达功能蛋白是解决该问题的有效方法[7]。现有研究发现,腺病毒载体系统在导入目的基因后会因病毒载体导入时间的持续延长,出现外源基因丢失的现象,从而降低了目的基因表达效果[8]。慢病毒载体系统弥补了该缺陷,在目的基因表达持续时间上更具有优势。Milone 等[9-10]研究发现慢病毒可高效感染分裂中的BMSCs,成功把携带的目的基因导入细胞,使目的基因在细胞中持续、稳定、高效表达,从而进行基因治疗。本课题组前期研究也表明慢病毒载体系统介导的BMP2基因相对于腺病毒载体系统,在表达持续时间上更具有优越性[11]。本研究以慢病毒为载体系统,通过基因转染技术,将携带BMP2基因和EGFP基因载体整合进入BMSCs 中,使其在细胞内能够持续稳定地表达。EGFP作为一种示踪标记的报告基因,其成功表达不仅示踪标记了携带BMP2 基因细胞的准确位置,还可直接体现出细胞的转染效率[12]。本研究通过检测BMP2 mRNA和蛋白的表达,证实了BMP2重组慢病毒载体转染BMSCs 并诱导细胞成骨方向分化的可行性。
目前,病毒的转染和BMSCs 成骨分化及其观察的相关技术在国内外已较为成熟,现被普遍认可的主要有ALP、Ⅰ型胶原蛋白、成骨特异性蛋白(骨形态生发蛋白、骨桥蛋白等)、相关成骨分化调控因子(骨钙素泌量、成骨转录因子等)为成骨早期的标志物,而矿化结节形成可作为成骨晚期的标志物[13-14]。本研究通过检测骨化标志性产物ALP、Ⅰ型胶原及矿化结节验证BMSCs 的成骨分化能力,但这些生物化学检测方法只能间接反映细胞的成骨方向分化。因此,本研究同时利用扫描电镜和能谱分析技术,直接对细胞成骨分化晚期的矿化结节进行观察,对其钙、磷等元素含量进行测定,结果显示矿化结节微区Ca/P为1.52±0.13,接近羟基磷灰石的钙磷比1.67,说明矿化结节表面有钙、磷沉积物产生,可能为磷酸钙盐。而硫、氮、氧、碳元素为有机物主要元素,可能为细胞基质产生的细胞胶原成分及细胞蛋白复合物的有机成分。
BMSCs向成骨分化后的细胞矿化是其主导骨形成的前提与基础[4]。在细胞矿化过程中,其最基本的生物学特征是骨基质合成、分泌及成熟,而矿化结节是成骨细胞分化的成熟标志,同时也是成骨细胞行使成骨功能的主要形态学表现[15]。细胞矿化的表现是细胞外基质的矿化及无机钙、磷盐的沉积,在整个BMSCs 矿化结节形成过程中,细胞和基质起着非常重要的作用[16]。本研究在电镜下观察发现BMSCs成骨分化晚期,细胞会重叠生长,分泌大量细胞基质,矿化结节则凸出于细胞层之上,出现磷酸钙盐堆积,结构呈现疏松、粗糙,说明BMSCs 成骨分化后细胞和细胞基质在细胞矿化过程中发挥重要作用。发生细胞矿化的细胞通过大量分泌细胞基质,被自身大量分泌的细胞基质包裹围绕,从而使局部细胞基质浓缩集中成沉淀的磷酸钙盐离子,使磷酸钙盐离子结晶导致矿化结节形成。其形成机制可能为矿化后的BMSCs 细胞内高表达ALP,从而促进细胞基质的矿化,且其分泌的Ⅰ型胶原蛋白是构成矿化结节的蛋白支架,Ⅰ型胶原蛋白支架与钙、磷无机盐亲和,形成磷酸钙盐矿化结节。BMSCs 发生细胞矿化的分子调控机制可能是由于细胞矿化过程中编码细胞外基质成熟的基因表达增加,而与细胞增殖相关的基因表达下降,且与细胞内ALP活性、骨钙素合成及细胞外基质促进羟基磷灰石沉积的基因表达相关[17]。BMSCs成骨方向分化后的矿化过程实质上为钙、磷无机盐在细胞基质内沉积的过程,整个矿化过程始终伴随着钙、磷等元素的动态变化[18]。
综上所述,BMP2重组慢病毒转染兔BMSCs 后,BMP2基因整合入细胞基因组中,高效表达BMP2功能蛋白,诱导细胞向成骨方向分化并发生细胞矿化,在细胞矿化过程中通过细胞基质对无机盐离子的浓集、沉淀,最终形成无机磷酸钙盐的矿化结节,其钙磷比值非常接近人和动脉骨骼中的主要无机磷酸钙盐羟基磷灰石的成分比值,说明其存在一定联系,需要后续深入研究。
猜你喜欢
建材发展导向(2022年24期)2022-12-22
口腔医学(2021年10期)2021-12-02
环境卫生工程(2021年4期)2021-10-13
看世界·学术下半月(2020年7期)2020-09-10
广东农业科学(2017年10期)2018-01-25
中国医学创新(2017年22期)2017-11-15
中华老年口腔医学杂志(2016年2期)2017-01-15
中国病理生理杂志(2015年8期)2015-12-21
天津护理(2015年4期)2015-11-10
甘肃农业科技(2015年12期)2015-04-22
