
外源水杨酸对低温胁迫下谷子幼苗生长的影响
2023-05-06 13:59闫锋
江苏农业科学 2023年7期
闫 锋
(黑龙江省农业科学院齐齐哈尔分院,黑龙江齐齐哈尔 161006)
谷子是一种起源于我国的古老粮食作物,在我国已经有8 000年左右的种植历史,对我国北方旱作农耕文明影响深远[1]。谷子具有适应性广、耐瘠薄等特点,是典型的环境友好型作物,也是一种重要的战略储备作物[2-3]。黑龙江地处高纬度,由于独特的地理环境导致春季气温变化大,倒春寒频发[4-5],低温冷害已成为导致谷子产量低且品质差的主要原因,如何缓解春季低温对谷子幼苗的伤害是谷子生产中亟待解决的问题。因此,研究谷子苗期耐低温机制及栽培技术,提高谷子苗期耐低温能力,对该地区谷子的生产具有重要意义。低温可抑制植物细胞代谢,从而使活性氧自由基(ROS)大量积累,加剧细胞膜脂过氧化损伤[6-7]。植物内源激素可以调控植物的生长及对逆境的应答[8],水杨酸作为一种广泛存在于植物组织中的内源激素,在植物的抗寒、抗旱、盐胁迫、重金属胁迫等非生物胁迫方面起着重要的调节作用[9]。外源激素通过改变内源激素水平调节植物生理代谢,利用外源激素对作物预先处理,被广泛利用于作物抗逆栽培中[10]。
前人研究发现,可以通过增加水杨酸在细胞中的含量,促进植物在低温逆境中的光合作用,进而提高植物叶片的光合效率[11-13];提高抗氧化酶活性,减缓细胞膜脂过氧化作用,稳定膜的结构与功能,增强细胞膜的抗逆能力,以适应低温逆境的变化[14-16];提高脯氨酸、可溶性蛋白、可溶性糖等渗透调节物质的含量,抵御低温冷害[17-19]。虽然前人关于水杨酸对缓解低温胁迫作用的机制做了大量研究,但关于谷子方面的研究仍鲜见报道。本试验在前期研究的基础上,以冷敏感品种鲁谷7号为试材,探讨不同浓度外源水杨酸对谷子遭受低温冷害的缓解作用,并明确其调控作用,筛选出最适外源水杨酸浓度,以期为研究水杨酸缓解谷子低温冷害的作用机制提供参考。
1 材料与方法
1.1 供试材料
选用前期试验筛选出的冷敏感谷子品种鲁谷7号为研究材料,该材料由中国农业科学院作物科学研究所陆平研究员提供。水杨酸购自天津市致远化学试剂有限公司。
1.2 试验设计
试验于2022年春季在黑龙江省农业科学院齐齐哈尔分院实验室进行,将种子浸入温水中催芽 12 h,露白后每个处理挑取100粒饱满、均匀一致的种子播种于小塑料盆中(每盆播种5粒,20盆作为1个处理),待谷苗长到3叶1心时,用浓度为0(T0)、50(T1)、100(T2)、150(T3)、200 mg/L(T4)的水杨酸溶液对谷子幼苗叶片进行叶面喷施处理,施药处理后2 d将谷子幼苗放入人工气候箱进行低温处理,胁迫温度为12 ℃/6 ℃ (昼12 h/夜12 h),以 25 ℃ 喷施清水为对照,每个处理设3次重复,处理时间为7 d。
1.3 测定项目与方法
低温胁迫7 d时从各处理随机选取10株谷苗进行根长、叶长(顶二叶)、苗鲜质量(地上部)等相关形态指标的测定。低温胁迫处理0、3、5、7 d时,从各处理随机选取5株叶片剪碎混合后进行相关生理生化指标的测定,分别参照刘萍等的方法采用酸性茚三酮法测定脯氨酸含量,采用硫代巴比妥酸法测定丙二醛(MDA)含量,采用氮蓝四唑法测定超氧化物歧化酶(SOD)活性,采用愈创木酚法测定过氧化物酶(POD)活性[20]。低温胁迫处理0、3、5、7 d时,从各处理随机选取5株,使用CI-340型光合作用测定仪测定谷子倒二叶中部位置的蒸腾速率、净光合速率、气孔导度。
1.4 数据处理与分析
采用SPSS 20.0进行显著性分析,用Excel 2019软件进行数据统计和作图。
2 结果与分析
2.1 外源水杨酸对低温胁迫下谷子幼苗生长的影响
由表1可知,低温胁迫处理7 d时除了T2处理下苗鲜质量与对照差异不显著外,其余各处理下谷子幼苗的根长、叶长及苗鲜质量均显著低于对照。不同水杨酸浓度处理下谷子幼苗的根长、叶长及苗鲜质量均高于T0处理,随着水杨酸溶液浓度上升,谷子幼苗各形态指标呈先上升后下降的变化规律,说明一定浓度范围内的外源水杨酸可以缓解低温冷害对谷子幼苗造成的伤害,但随着浓度的增加缓解作用逐渐减弱。所有水杨酸处理中以T2处理下谷子幼苗的根长、叶长及苗鲜质量最大,比T0处理分别增加了26.56%、29.76%、36.36%,说明T2处理下低温对谷子生长的抑制作用最小。

表1 外源水杨酸处理对低温胁迫7 d 后谷子幼苗生长的影响
2.2 外源水杨酸对低温胁迫下谷子幼苗光合指标的影响
2.2.1 水杨酸对低温胁迫下谷子幼苗气孔导度的影响 由图1可知,胁迫0 d时各处理间气孔导度差异不显著,处理3、5、7 d时各处理气孔导度均显著低于对照,随着胁迫时间延长谷子幼苗的气孔导度呈逐渐降低的变化趋势,且均在处理7 d时达到最低,说明低温胁迫逐渐加重。除胁迫0 d外,各水杨酸浓度处理下的气孔导度均高于T0处理,且差异性显著,随着水杨酸处理浓度增加,谷子幼苗的气孔导度表现为先上升后下降。在胁迫3、5、7 d时,除对照和T0处理外,以T2处理下气孔导度最高,T4处理最低,其中T2 处理的气孔导度均显著高于其他喷施处理,缓解低温胁迫效果最好。

2.2.2 水杨酸对低温胁迫下谷子幼苗蒸腾速率的影响 由图2可知,胁迫0 d时各处理的蒸腾速率与对照差异不显著,随着胁迫时间延长谷子幼苗叶片的蒸腾速率逐渐下降,且均在处理7 d时达到最低,与对照相比均显著降低,说明低温胁迫逐渐加重。随着水杨酸处理浓度增加,谷子幼苗叶片的蒸腾速率先上升后下降,T2处理下的蒸腾速率最高,在胁迫3、5、7 d时比T0处理分别高22.86%、21.21%、34.48%。

2.2.3 水杨酸对低温胁迫下谷子幼苗净光合速率的影响 由图3可知,胁迫0 d时各处理与对照净光合速率差异不显著,随着胁迫时间的延长,各处理谷子幼苗的净光合速率逐渐降低,且均在胁迫7 d时达到最低,各处理均与对照差异显著,说明低温胁迫作用逐渐加重。水杨酸处理组在胁迫5、7 d时的净光合速率均显著高于T0处理,随着水杨酸处理浓度增加,净光合速率先上升后下降,说明一定浓度的水杨酸溶液能缓解低温胁迫对光合作用的抑制。在胁迫3、5、7 d时,除对照和T0处理外,均以T2处理下净光合速率最高,T4 处理最低,其T2处理的净光合速率比T0处理分别高34.96%、50.96%、71.76%。

2.3 外源水杨酸对低温胁迫下谷子幼苗生理指标的影响
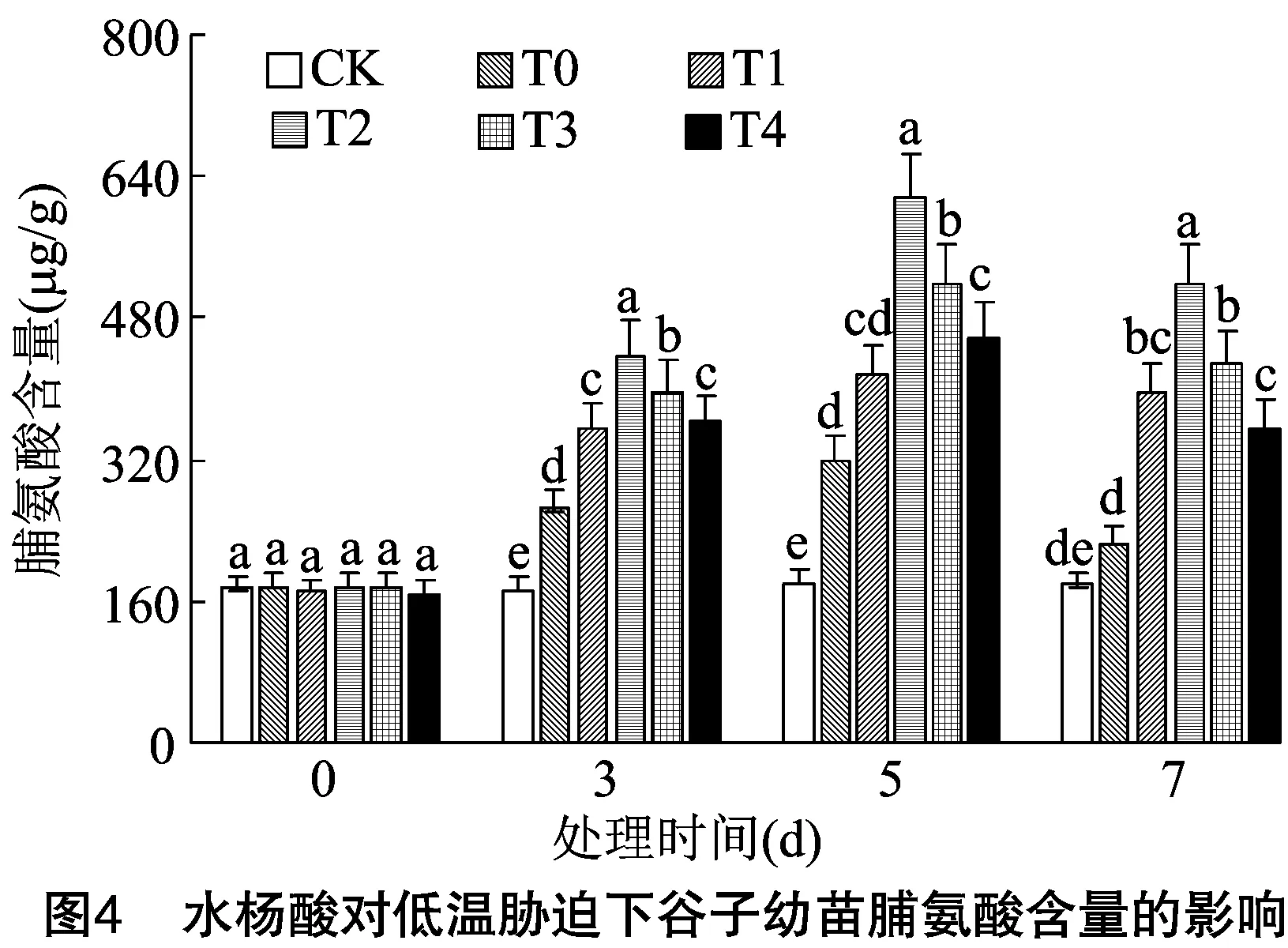
2.3.1 对渗透调节物质含量的影响 由图4可知,低温胁迫0 d时各处理与对照脯氨酸含量差异不显著;在胁迫3、5、7 d时,除胁迫7 d的T0处理外,其余各处理的脯氨酸含量均显著高于对照,随着胁迫时间的推移谷子幼苗中脯氨酸含量均表现为先上升后下降,在胁迫5 d时达到最高。与对照和T0处理相比,喷施不同浓度水杨酸溶液均可使脯氨酸含量显著升高,表明在低温胁迫下谷子叶片内部迅速合成脯氨酸,且经过水杨酸处理可显著提高脯氨酸含量,缓解了低温对谷子幼苗的伤害。所有处理中,T1和T4处理在低温胁迫3、5、7 d时差异均不显著,T1和T3处理在胁迫7 d时差异不显著。低温胁迫3、5、7 d时,均以T2处理下脯氨酸含量最高,比T0处理分别高66.10%、93.50%、129.77%。
2.3.2 对膜脂过氧化程度的影响 由图5可知,低温胁迫3、5、7 d时,各处理MDA含量均与对照差异显著,随着胁迫时间的推移,谷子幼苗叶片中的MDA含量逐渐上升,并在胁迫7 d时达到最高。随着水杨酸处理浓度增加,谷子幼苗叶片中MDA含量先下降后上升,与T0相比,除了T4处理在低温胁迫7 d外,其余处理均可使MDA含量显著降低;所有处理中,T3和T4处理在低温胁迫3、5、7 d时差异均不显著,T1、T3、T4处理在胁迫3 d时差异不显著,T1、T3处理在胁迫5 d时差异不显著。低温胁迫3、5、7 d时,除对照处理外,均以T2处理下MDA含量最低,在胁迫3、5、7 d时T2比T0处理分别降低24.07%、33.08%、33.28%。

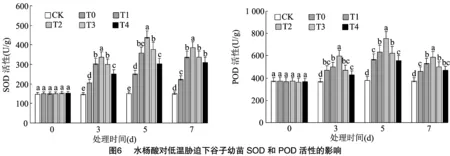
2.3.3 对抗氧化酶活性的影响 由图6可知,低温胁迫0 d时各处理与对照SOD、POD活性差异均不显著,随着胁迫时间推移,SOD与POD活性均表现为先上升后下降,各处理的SOD与POD活性均在胁迫5 d时达到最高且与对照差异显著。不同浓度水杨酸处理下的SOD与POD活性差异显著性有所不同,在胁迫3 d时,T0处理的POD活性与T1、T3、T4处理差异不显著,在胁迫5、7 d时,T0处理的POD活性与T4处理差异不显著,在胁迫3、5、7 d时T0处理的SOD活性与其余各处理均差异显著。在胁迫3、5、7 d时,均以T2处理的抗氧化酶活性最高。
3 讨论
3.1 水杨酸对低温胁迫下谷子幼苗生长的影响
水杨酸可提高植物对多种生物胁迫及非生物胁迫的抵抗能力[21]。李颖等的研究表明,低浓度水杨酸处理可以有效缓解低温对扁蓿豆发芽、幼苗生长的抑制作用[22]。张荣萍研究认为,水杨酸预处理可使巨胚稻幼苗抗寒性提高,对幼苗地上部分和根系生长具有一定的促进作用[23]。本研究表明,在低温胁迫7 d后,除T2处理下苗鲜质量与对照差异不显著外,其余各处理在低温胁迫后谷子幼苗的根长、叶长、苗鲜质量均低于对照,说明低温对谷子幼苗生长具有显著的抑制作用。谷子幼苗根长、叶长及苗鲜质量随着水杨酸处理浓度的增加表现为先上升后下降,说明一定浓度范围内外源水杨酸处理可以有效缓解低温对谷子幼苗的胁迫,但随着浓度的增加缓解作用逐渐减弱,呈现出一定的剂量效应,这与王星剑等的研究结果[24]一致。
3.2 水杨酸对低温胁迫下谷子幼苗光合指标的影响
对植物光合作用以及植物内部光合原件进行研究,发现低温胁迫影响植物生长,并直接影响内部光合原件的结构与活性,间接影响光合作用,光合作用的强弱也可以通过光合参数反映出来。杨朴丽等研究认为,诺丽幼苗遭遇低温冷害后,其叶片各项光合指标与对照相比显著降低[25]。孙玉珺等对玉米的研究表明,外源水杨酸能提高低温胁迫下玉米幼苗的净光合速率[26]。本研究表明,除胁迫0 d外,经低温胁迫处理后谷子幼苗的各项光合指标均显著低于对照,喷施不同浓度水杨酸溶液后,T1~T4处理各光合指标均高于T0处理,说明一定浓度范围内的水杨酸溶液能缓解低温胁迫下谷子幼苗光合指标的降低,该结果与周艳丽等对甜瓜的研究结果[11]相一致。
3.3 水杨酸对低温胁迫下谷子幼苗细胞膜的影响
低温胁迫下植物细胞膜会发生各种膜脂过氧化反应,导致细胞 MDA 含量增加[27]。辛慧慧等研究发现,棉花幼苗经过低温胁迫后,叶片中MDA含量与对照相比显著增加,施用外源水杨酸后,丙二醛的产生受到抑制[28]。本研究表明,各水杨酸浓度处理的谷子幼苗经低温处理后,在胁迫3、5、7 d时,叶片中MDA含量均低于T0处理,这与前人的研究结果相一致。产生这一现象的原因可能是低温胁迫导致谷子幼苗细胞膜脂过氧化,促进了丙二醛的大量产生,外源水杨酸的施用保护了细胞膜的稳定性,维持了细胞结构的完整性。研究表明,植物细胞内的脯氨酸在植物受到低温胁迫时参与抵御作用,脯氨酸含量的增加可以在一定程度上维持细胞膜的渗透势,保持水分平衡[29]。本研究表明,与对照相比,除胁迫0 d外,低温胁迫下不同水杨酸浓度处理均能提高脯氨酸含量,随着时间推移脯氨酸含量均表现为先上升后下降,这可能与谷子叶片逐渐适应低温环境有关。
3.4 外源水杨酸对低温胁迫下谷子幼苗抗氧化系统的影响
植物在受到低温胁迫时,细胞中的活性氧自由基大量积累,造成细胞损伤,植物可以通过 SOD、POD和 CAT 三者协同作用清除大量自由基,从而使细胞内氧自由基维持在较低水平[30]。本试验结果表明,除胁迫0 d外,各水杨酸处理能够显著提高低温胁迫下谷子叶片的 SOD、POD活性,且抗氧化酶活性随水杨酸浓度增加呈现先上升后下降的趋势。这与秦文斌等的研究结果[31-32]一致,说明适宜浓度外源水杨酸的施用能够提高低温胁迫下谷子幼苗的抗氧化酶活性,维持谷子幼苗对抗活性氧自由基的能力,缓解低温带来的过氧化伤害,提高幼苗对低温的适应能力。
4 结论
低温胁迫处理7 d时除了T2处理下苗鲜质量与对照差异不显著外,其余各处理下谷子幼苗的根长、叶长及苗鲜质量均显著低于对照。除胁迫0 d外,随着胁迫时间推移谷子幼苗中脯氨酸含量均表现为先上升后下降;MDA含量表现为持续升高;抗氧化酶SOD、POD活性均较对照显著增强,且均表现为先上升后下降的变化趋势;各光合指标均显著低于对照,且随着时间推移各光合指标持续降低。除胁迫0 d外,与T0处理相比,T1~T4处理下的MDA含量均降低,根长、叶长、苗鲜质量及脯氨酸含量升高,抗氧化酶活性增强,以100 mg/L水杨酸处理对谷子幼苗低温胁迫的缓解效果最好。
猜你喜欢
中老年保健(2022年3期)2022-11-21
当代陕西(2021年21期)2022-01-19
科学与财富(2019年15期)2019-10-21
发酵科技通讯(2018年2期)2018-07-06
湖南林业科技(2017年1期)2017-02-06
中国药物应用与监测(2015年5期)2015-12-11
化工进展(2015年6期)2015-11-13
结核与肺部疾病杂志(2015年3期)2015-07-18
现代农业(2015年5期)2015-02-28
应用化工(2014年11期)2014-08-16
