
低剂量电离辐射诱导眼晶状体上皮细胞中差异mRNA表达谱的鉴定
2023-04-29 15:37:14王萍范莉陆雪等
辐射防护 2023年2期
王萍 范莉 陆雪等


关键词:低剂量电离辐射;眼晶状体浑浊;生物信息学;增殖
中图分类号:Q345 文献标识码:A
眼晶状体一直被认为是对辐射最敏感组织之一,流行病学对辐射诱导眼晶状体混浊的队列研究发现,长期受到低剂量辐射的人群,比如核工业工作者[1] 、医院介入心脏病学家[2] 、X 射线工作者[3] 、宇航员和飞行员[4] 等,会增加眼晶状体混浊的发生风险, 且大多发生在皮质或者后囊下。2011 年,主要基于流行病学的研究结果,国际放射防护委员会( ICRP) 推荐眼晶状体剂量阈值为0. 5 Gy。然而,近年美国关于放射技术人员的队列研究结果显示,低剂量(<0. 1 Gy) 和低剂量率(通常<0. 005 Gy/ h)照射后白内障发生风险显著增加[5] 。我国在阳江高本底地区居民的流行病学研究发现,当眼晶状体累积剂量达0. 1 Gy 时,患皮质及后囊下浑浊的风险升高,且发生皮质性浑浊的剂量阈值估值为0. 14 Gy[6] 。这提示对于容易接触低剂量辐射的人群,诱导晶状体浑浊的剂量限值可能要比ICRP 推荐的0. 5 Gy 低。但是,低剂量电离辐射诱导眼晶状体浑浊的机制目前还未完全了解,只提出了一些潜在的途径,包括氧化应激或电离辐射诱导的DNA 损伤可能修复缓慢[7] ,导致细胞增殖、分化异常;DNA 损伤与修复可能存在逆剂量率效应[8-9] ;端粒长度增加和端粒酶活性降低等也可能与白内障发生相关[10-11] 以及遗传背景不同会显著改变低剂量电离辐射诱导白内障的发生风险等[12-14] 。相比于低剂量, 较高剂量(>2 Gy)辐射诱导白内障的机制已经比较明确,主要是晶状体上皮细胞基因组损伤,包括氧化应激、DNA 损伤、细胞间通讯、细胞极性改变等[15] 。为此,我们首先通过生物信息学对低剂量(0. 2 Gy)和较高剂量(2 Gy)γ 射线照射人晶状体上皮细胞的差异基因表达谱和相关分子通路进行分析,并对与细胞增殖相关的PIM1、HMGB1、BRE 和JUN差异表达基因进行初步验证,为进一步探索差异表达基因在晶状体上皮细胞中的功能提供实验依据,有助于揭示低剂量电离辐射诱导眼晶状体浑浊的分子机制。
1 材料与方法
1. 1 细胞培养、照射及样本收集
正常人晶状体上皮细胞SRA01/ 04 培养在含10% 胎牛血清的MEM 培养基( HyClone,SH30024. 01)或含10%胎牛血清的DMEM 高糖培养基(Gibco,C11965500BT)中,正常人晶状体上皮细胞HLE-B3 培养在含10%胎牛血清的EMEM 培养基(ATCC,80615210)中,培养条件为饱和湿度、37 ℃ 、5%CO2的培养箱。待SRA01/ 04 细胞长至80% 融合,分别进行0. 2 Gy137Cs 和2 Gy60Co γ射线照射,其中0. 2 Gy 剂量率为9. 73 mGy/ min,2 Gy 剂量率为1 Gy/ min,照射后继续培养48 h后,收集细胞, 用预冷的PBS 洗一遍, 加1 mLTrizol 裂解,置于-80 ℃ 冰箱保存,用于基因芯片分析。采用相同剂量率对SRA01/ 04 和HLE-B3两种细胞分别进行0、0. 05、0. 075、0. 1、0. 2、0. 5Gy137Cs γ 射線照射和2 Gy60Co γ 射线照射,照射后24 h、48 h 收集细胞,用于验证及分析不同剂量照射后不同时间晶状体上皮细胞差异基因转录水平的变化。
1. 2 总RNA 提取
细胞加Trizol 充分裂解后,加入200 μL 三氯甲烷,剧烈震荡15 s,室温静置2 ~ 3 min,于4 ℃ ,12 000 rpm 离心15 min;将上层水相吸至新EP 管中,再加入等体积异丙醇,轻轻混匀后室温静置10min,4 ℃ ,12 000 rpm 离心10 min,留底部白色沉淀,加入700 μL 无水乙醇、200 μL 焦碳酸二乙酯(DEPC)水,7 500 rpm 离心5 min,留底部沉淀,适当风干, 加入20 ~ 30 μL DEPC 水, 得到RNA溶液。
1. 3 mRNA 芯片分析
采用Affymetrix 芯片进行分析(北京中康博生物科技有限公司)。具体实验过程为:样本总RNA通过质控后,依次进行Poly A 加尾、片段化标记、芯片杂交、芯片洗染、扫描等过程。使用GeneChip3000 7G 扫描仪捕获荧光信号,并通过GCOS 软件将信号转化,从而获得每个探针的信号值,生成CEL 文件,进行数据标准化,筛选变化倍数大于2(FC>2)的差异表达基因。
1. 4 mRNA 功能的生物信息学分析
利用基因本体学(Gene Ontology, GO)数据库和京都基因与基因组大百科全书 ( Kyoto Encyclopedia of Genes and Genomes, KEGG) 对差异表达的基因进行功能和信号通路的显著性分析。
1. 5 实时荧光定量PCR( Real-time PCR) 检测
采用PrimeScript RT reagent Kit With gDNAEraser( 日本TaKaRa) 将样本总RNA 逆转录成cDNA,应用ABI7500 快速实时PCR 系统检测mRNA 表达量。以GAPDH 作为内源性内参基因进行归一化处理,以2-ΔΔCT法计算照射组与未照射组间mRNA 表达水平的变化。所有样品检测均设置3 个平行样,至少重复3 次。PCR 引物列于表1。
1. 6 统计学处理
采用SPSS 25. 0 软件进行统计学分析,数据以x±s表示。在数据符合正态分布和方差齐性条件下,多组之间比较使用方差分析,若方差不齐选择秩和检验。p<0. 05 为差异具有统计学意义。
2 结果
2. 1 不同剂量γ 射线照射SRA01 / 04 细胞后差异表达基因及生物学功能与通路分析
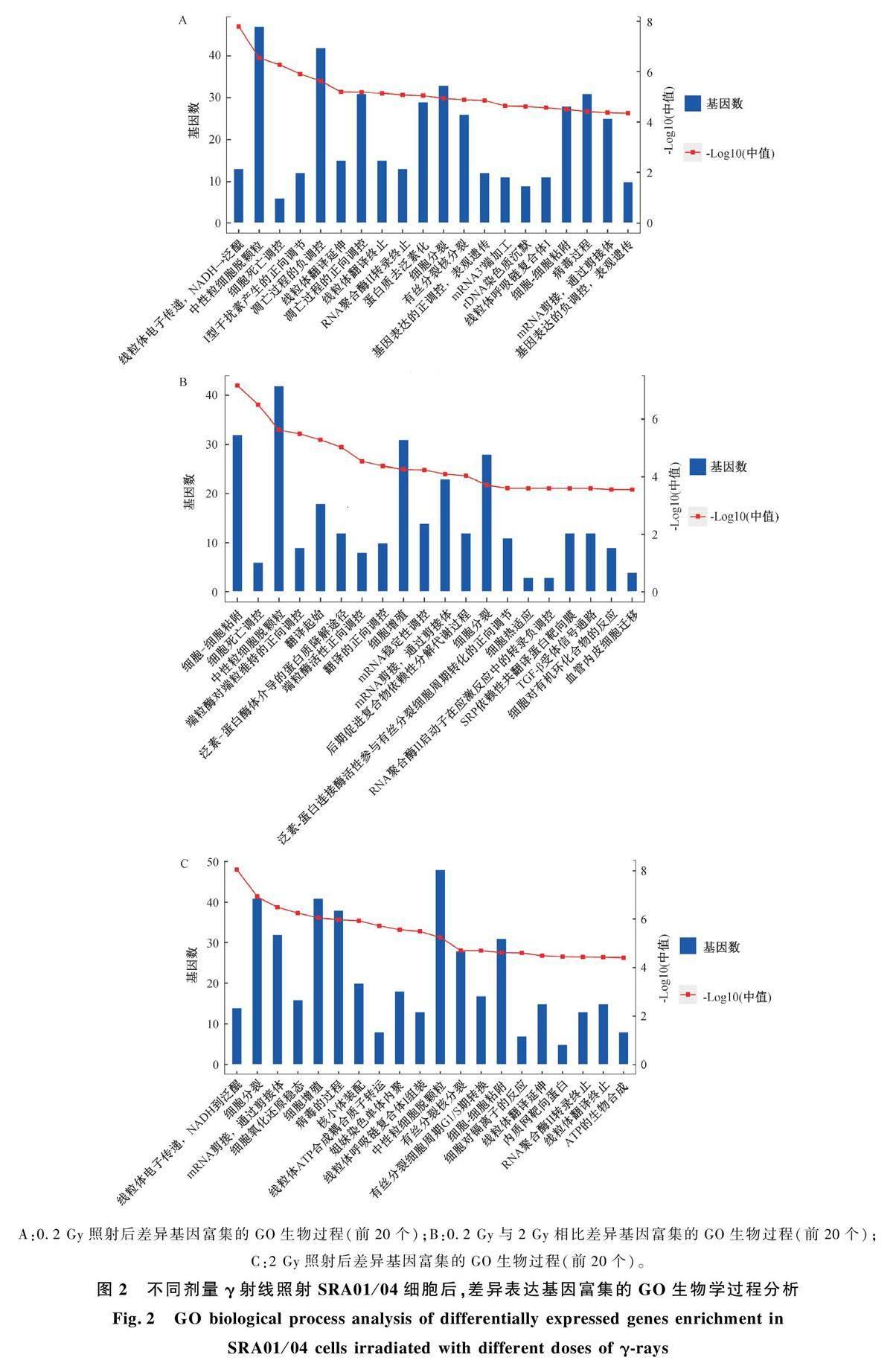
通过芯片分析筛选出了高、低剂量照射后48 h 变化倍数大于2 的基因,各照射组之间相互比较筛选出的共同表达的基因和单独表达的基因如图1 所示。发现较高剂量(2 Gy)照射后差异表达基因要比0. 2 Gy 照射后基因数增多,2 Gy 照射后差异表达基因共3 970 个,1 960 个表达上调,2 010 个表达下调,单独表达的基因1 495 个;而0. 2 Gy 照射后差异表达基因共3 304 个,1 640 个表达上调,1 664 个表达下调,单独表达的基因1 144 个;此外,0. 2 Gy 与2 Gy 相比共有3 690 个差异表达基因,1 862 个表达上调,1 828 个表达下调,单独表达的基因1 285 个。
图2 和图3 分别给出了不同比较组差异表达基因的GO 生物过程通路和KEGG 功能通路的生物信息学分析结果。首先是0. 2 Gy 与0 Gy 比较组,GO 生物过程通路按统计学显著性排前三位的分别是线粒体的电子传递链、中性粒细胞脱颗粒、细胞死亡调控,差异基因主要聚集在细胞分裂、细胞粘附等过程,KEGG 通路分析发现p <0. 01 的信号通路有氧化磷酸化、DNA 复制、衰老等;0. 2 Gy与2 Gy 比较组,GO 生物过程通路按统计学显著性排前三位的分别是细胞粘附、细胞死亡调控、中性粒细胞脱颗粒,差异基因主要聚集在细胞粘附、细胞增殖、细胞分裂过程,KEGG 通路分析发现p<0. 01 的信号通路有内质网中蛋白质加工、紧密连接等,其他还有细胞周期、氧化磷酸化、错误修复、粘附连接等;2 Gy 与0 Gy 比较组,GO 生物过程通路按统计学显著性排前三位的分别是线粒体的电子传递链、细胞分裂、mRNA 剪接,差异基因主要聚集在细胞分裂、细胞增殖、细胞粘附以及有丝分裂细胞周期G1 -S 期转换过程,KEGG 通路分析发现p<0. 01 的信号通路有氧化磷酸化、细胞周期、DNA复制、紧密连接等,其他还有TGF-β 信号通路等。
2. 2 高、低剂量照射SRA01 / 04 细胞后基因表达差异
为了探究低剂量电离辐射诱导眼晶状体浑浊的分子机制,并与高剂量电离辐射诱导白内障的机制比较异同,我们将重点分析0. 2 Gy 与0 Gy 比较组和0. 2 Gy 与2 Gy 比较组的差异表达基因。GO 生物过程和KEGG 结果表明,0. 2 Gy 与未照射(0 Gy)和较高剂量(2 Gy)相比,相同之处可能在于差异表达基因都与细胞增殖、细胞粘附这一生物过程相关。细胞增殖、粘附相关的差异表达上调的基因(FC>2)见表2、表3,基因表达的热图分析见图4。0. 2 Gy 照射后,差异基因变化倍数最高的前三位分别是原癌基因PIM1、促炎因子IL-1β和编码抗凋亡蛋白的BRE,0. 2 Gy 与2 Gy 相比,差异基因变化倍数最高的前三位分别是编码抗凋亡蛋白的BRE、促炎因子IL-18 和DNA 修复酶XRCC6,两比较组相同的基因为:BRE、JUN、IL -1β、GADD45A、S100A7A。
2. 3 Real-time PCR 验证差异表达的基因
选择0. 2 Gy 照射后表达水平显著上调的基因,以及在0. 2 Gy 与0 Gy 和0. 2 Gy 与2 Gy 比较组中表达水平都显著上调的基因( PIM1、BRE、JUN)进行Real-tiem PCR 验证,并且在0. 2 Gy 照射后表达水平下调的基因中也选择了一个差异倍数变化4. 659 倍且与细胞增殖相关的HMGB1 基因。SRA01/ 04 细胞和HLE-B3 细胞分别经不同剂量(0、0. 05、0. 075、0. 1、0. 2、0. 5 和2 Gy)γ 射线照射后24 h、48 h,利用Real-time PCR 检测差异基因表达变化。相比于保持着稳定的上皮细胞极性的SRA01/ 04 细胞,HLE-B3 细胞在连续培养中,不仅具有细胞增殖能力,还保持着稳定分化能力。
结果显示,SRA01/ 04 细胞中(图5),PIM1 的表达量在照射后24 h,大部分低剂量组相比于未照射组表达水平都显著升高( p < 0. 05);照射后48 h,相比于未照射组,0. 05 Gy 和0. 1 Gy 照射组表达水平显著升高(p<0. 05),较高剂量(2 Gy) 组表达水平显著下降(p <0. 05)。HMGB1 表达量在照射后24 h、48 h,相比于未照射组,某些低剂量点有升高趋势,其中0. 075 Gy 照射后24 h,HMGB1表达水平显著升高(p<0. 05),而2 Gy 照射后表达水平显著下降(p <0. 05)。BRE 的表达水平在低剂量照射后24 h 有随剂量升高而升高的趋势,到较高剂量点(2 Gy) 表达回到基线水平;照射后48 h,低剂量组相比于未照射组表达水平有升高趋势,2 Gy 组表达水平显著下降(p <0. 05)。JUN的表达量在0. 05 Gy、0. 075 Gy 照射后24 h 升高,2 Gy 照射后显著升高( p < 0. 05),其余低剂量组JUN 的表达水平都显著下降( p < 0. 05);照射后48 h,表达量变化与基因芯片分析结果相反。HLE-B3 细胞中( 图6), PIM1 的表达变化与SRA01/ 04 细胞中表达趋势基本一致,但表达水平升高不显著(p >0. 05)。HMGB1 的表达量在照射后24 h 表现为随剂量升高而升高,照射后48 h 大部分低剂量组相比于未照射组表达水平也有升高趋势。BRE 的表达量在低剂量照射后24 h 和0. 05 Gy、0. 075 Gy 照射后48 h,与未照射组相比表达水平显著升高(p <0. 05)。JUN 的表达量在0. 075 Gy、0. 2 Gy 和2 Gy 照射后48 h 显著升高(p<0. 05),其余时间点和剂量点未见明显变化。综上, 在遗传背景不同的两种细胞中, PIM1、HMGB1、BRE 在低剂量电离辐射照射后剂量响应比较好,照射后24 h,大部分低剂量组相比于未照射组表达水平明显升高,48 h 仍然有升高趋势,但在某些低劑量组升高不显著,并且这三个基因在SRA01/ 04 细胞2 Gy 照射后48 h,表达水平显著下降,在HLE-B3 细胞中也有表达水平下降趋势。这提示低剂量电离辐射作用于眼晶状体可以诱导某些与刺激细胞增殖、迁移相关的基因表达升高,可能与低剂量电离辐射诱导晶状体浑浊有关。
3 讨论
随着流行病学随访调查的更加系统全面、更准确的照射剂量测量方法以及新诊断技术的发展,研究结果均表明急性和长期受到低剂量辐射的人群发生眼晶状体浑浊的风险增加。因此,ICRP 在2011 年大幅降低了晶状体的剂量阈值至0. 5 Gy(原为2 Gy),对于职业照射,每年的晶状体当量剂量限值也从原来150 mSv 大幅降低至20mSv,且连续5 年内任何一年都不能超过50mSv[16] 。新建议的提出也影响了对低剂量电离辐射如何导致晶状体浑浊的机制研究。
Hamada[17] 提出眼晶状体细胞不一定对细胞死亡具有辐射敏感性,其对辐射的敏感性可能与上皮细胞过度增殖、纤维细胞异常分化、DNA 双链断裂修复不当或缓慢、端粒效应、衰老、晶状体蛋白改变、非靶向效应、炎症以及遗传因素有关。对于易受低剂量辐射的人群,诱导晶状体浑浊的剂量限值可能要比ICRP 推荐的0. 5 Gy 更低,低剂量电离辐射诱发晶状体浑浊的分子机制可能与高剂量不同。体内研究表明,低剂量尤其是0. 1 Gy、0. 25 Gy X 射线照射后24 h,小鼠晶状体外周区细胞密度和增殖明显增加,cyclin D1 的表达水平也升高,而高剂量1 Gy 和2 Gy 照射后cyclin D1 的表达水平显著降低[18] 。Vigneux 等[19] 研究发现,HLE-B3 细胞受0. 25 Gy X 射线照射后7 天,细胞增殖和迁移能力显著增强, 而较高剂量(0. 5 ~2 Gy)时回落到未照射水平,且高于0. 25 Gy 照射后细胞粘附能力持续降低。有研究也发现小于0. 5 Gy 的X 射线照射人晶状体上皮细胞,活性氧和活性氮的生成略有降低、细胞活力增强、细胞增殖能力也增强,而大于0. 5 Gy 照射后,产生了大量的活性氧和活性氮、细胞活力降低,表明了低剂量电离辐射可以激活抗氧化机制,有效清除活性氧,而较高剂量照射下,过量的活性氧可能导致细胞死亡并诱发白内障[20] 。
本文筛选了高、低剂量照射眼晶状体上皮细胞后差异表达基因,并进行GO 生物过程和KEGG分析。GO 生物过程表明,0. 2 Gy 照射后,差异表达基因显著富集在细胞分裂、细胞死亡调控、细胞粘附等方面;0. 2 Gy 与2 Gy 相比,差异表达基因显著富集在细胞粘附、细胞增殖、细胞死亡调控等方面;2 Gy 照射后,差异表达基因显著富集在细胞分裂、细胞增殖、有丝分裂细胞周期G1 -S 期转换、细胞粘附等方面。同样, KEGG 分析也表明,0. 2 Gy 照射后,差异基因显著富集的信号通路是氧化磷酸化、与炎症相关的IL-17 信号通路、与衰老相关的信号通路、DNA 复制以及与细胞增殖、分化相关的MAPK 信号通路等;0. 2 Gy 与2 Gy 相比,差异基因显著富集的信号通路除氧化磷酸化外,还有与细胞粘附相关的粘附连接和紧密连接、细胞周期、错配修复等;2 Gy 照射后,差異基因显著富集的信号通路是氧化磷酸化、细胞周期、DNA复制、紧密连接、TGF-β 信号通路等。综上表明电离辐射诱导晶状体上皮细胞功能改变在氧化磷酸化、细胞增殖、细胞粘附等方面与正常晶状体上皮细胞相比有显著差别。本研究结果中发现,低剂量电离辐射照射晶状体上皮细胞后与较高剂量照射后相比,KEGG 分析显示差异表达基因在错配修复通路中显著富集,这提示低剂量电离辐射引起晶状体上皮细胞DNA 损伤后修复可能不当,从而导致基因组不稳定。已有文献发现,低能X 射线照射小鼠头部,72 h 后仍能在晶状体细胞核中检测到大量的DNA 氧化损伤标志物8-OHG 以及DNA 单链修复蛋白XRCC1,随后几个月观察到上皮细胞异常迁移到内部,表明初始细胞DNA 损伤遗传给子细胞并引起细胞异常行为,导致白内障[21] 。Ahmadi 等人[7] 用低剂量( <0. 5 Gy) γ 射线照射人晶状体上皮细胞,发现照射后24 h DNA损伤仍然存在,作者推测这种持续未修复的损伤可能导致基因组突变,传递给子细胞,导致基因组不稳定并发生白内障。
本研究应用基因芯片技术筛选了可能与低剂量电离辐射诱导眼晶状体浑浊发生发展的关键基因,对其中与细胞增殖相关的候选基因PIM1、HMGB1、BRE 和JUN 进行了初步验证。两种人晶状体上皮细胞受不同低剂量照射后, 细胞内PIM1、HMGB1 和BRE 基因发生不同程度的表达水平升高变化。我们观察到HMGB1 在两种细胞中表达水平的变化趋势不太一致,在SRA01/ 04 细胞中,HMGB1 只在0. 075 Gy 照射后24 h 表达水平显著升高,2 Gy 照射后表达水平显著降低;而HLE-B3 细胞中,HMGB1 在0. 2、0. 5 和2 Gy 照射后24 h 以及0. 05 Gy 照射后48 h 表达水平显著升高。这种差异可能是由于两种细胞遗传背景和细胞特性不同,对辐射敏感性也不同。遗传背景在低剂量电离辐射诱导晶状体浑浊中发挥着重要作用。体外研究发现来自不同供体的HLE-B3 细胞和HLEC 细胞经0. 1、0. 25 和0. 5 Gy ( 剂量率:0. 065 mGy/ min)照射后1 h,HLE-B3 细胞活力在各剂量点显著降低,而HLEC 细胞只在0. 5 Gy 照射后细胞活力降低, 表明HLE-B3 细胞相比于HLEC 细胞对辐射更加敏感[7] 。
在其他细胞类型,PIM1 在视网膜中过表达可激活Stat3、Akt1 和Akt2 通路,并通过线粒体通路抑制神经节细胞凋亡,促进受损视神经再生和功能恢复[22] 。此外,PIM1 在血管平滑肌细胞中表达升高通过调控细胞周期,诱导大多数细胞阻滞于S期和G2 / M 期, 促进细胞增殖和存活[23] 。Mohammad 等人[24] 研究表明,HMGB1 在人视网膜微血管内皮细胞中过表达可诱导MMP-9 表达上升,细胞迁移能力显著增强,促进血管生成。BRE基因主要在大脑和生殖器官中表达,在细胞存活、分化、凋亡和再生中发挥重要作用。Yeung 等人[25] 研究发现,BRE- / - 小鼠卵巢中卵泡数量变少,颗粒细胞增殖受到抑制,卵泡闭锁增加,且γH2AX和与DNA 损伤相关基因的表达升高,表明卵巢中缺少BRE 基因表达会使DNA 损伤修复效率低,导致细胞过度凋亡,进而诱导闭锁卵泡的形成。目前关于PIM1、HMGB1 或BRE 基因在眼晶状体方面的功能研究很少。根据本研究结果,我们推测细胞增殖相关的PIM1、HMGB1、BRE 基因在低剂量电离辐射照射人晶状体上皮细胞后表达水平的变化,可能与低剂量电离辐射诱导晶状体上皮细胞异常增殖和迁移密切相关。但仍需进一步体内、体外研究确定这些基因在晶状体上皮细胞中的作用机制。
综上所述,本研究对较低剂量(0. 2 Gy) 和较高剂量(2 Gy)γ 射线照射人晶状体上皮细胞后差异表达基因的生物学功能和通路进行了预测分析和初步验证,为进一步阐明低剂量电离辐射诱导晶状体浑浊及白内障的发生机制以及辐射防护提供了实验依据和可能的分子靶点。
猜你喜欢
江苏农业科学(2016年8期)2017-02-15 19:14:01
江苏农业科学(2016年8期)2017-02-15 19:13:40
山东农业科学(2016年12期)2017-01-21 14:31:11
中国校外教育(下旬)(2016年11期)2016-12-27 11:42:42
中国现代医生(2016年27期)2016-12-21 16:05:30
中国教育信息化·基础教育(2016年10期)2016-12-20 20:09:27
今传媒(2016年11期)2016-12-19 11:35:50
电脑知识与技术(2016年27期)2016-12-15 19:19:25
中国科技纵横(2016年17期)2016-11-30 21:40:27
中国医药导报(2016年25期)2016-11-30 07:20:06
